
Abstract
Abstract
This study aimed to identify stromal molecular signatures associated with breast and prostate cancer. The microarray data GSE26910 was downloaded from Gene Expression Omnibus database, including six invasive breast tumor stroma, six matched normal controls, six invasive prostate tumor stroma, and six matched controls. The differentially expressed genes (DEGs) in invasive breast and prostate tumors stroma were, respectively, identified. Then common stromal genes (B_P.DEGs) were further screened. Protein–protein interaction (PPI) network was constructed and Gene Ontology analysis was performed. Besides, gene–chemical interactions were mapped in Comparative Toxicogenomics Database to screen the chemicals related to feature genes. The results showed that, in total, 16 B_P.DEGs were identified. Thereinto, only seven B_P.DEGs were mapped into PPI, and only four functional modules (adenylate cyclase activating polypeptide 1 (pituitary) receptor type I (ADCYAP1R1) module, aspartoacylase (ASPA) module, glutathione S-transferase mu 5 (GSTM5) module, and periplakin (PPL) module) were involved in important biological processes associated with cancer progression. In addition, the chemicals, such as dihydrotestosterone, apocarotenal, testosterone, and progesterone, were screened for the roles of feature genes in the progression of breast and prostate cancer. In conclusion, ADCYAP1R1, GSTM5, and PPL were stromal molecular signatures and might play a key role in the progression of breast and prostate cancer.
1. Introduction
In recent years, tumor stroma has increasingly becoming a focus in the field of cancer research (Vannucci, 2015; Wendler et al., 2017). Stromal changes during malignancy play an important role in promoting growth, invasion, and metastasis of cancers (Bremnes et al., 2011). However, many treatments mainly target the tumor cells and disregard the fact that cancers are heterogeneous and their growth is due to interactions between tumor cells, stromal cells, and microenvironment in which they live (Allegra et al., 2010; Khamis et al., 2012). Therefore, better understanding the signatures of the stromal environment may be equally important for future cancer therapies as well as understanding cancer genetics per se.
Extensive evidence has supported the significant contribution of stroma to the development of various tumors (Räsänen and Vaheri 2010; Eiró and Vizoso, 2012; Bebelman et al., 2018). The development of cancer are regulated by interactions with components of stroma, including fibroblasts/myofibroblasts, epithelial cells, and immune cells along with the extracellular matrix (ECM) and extracellular molecules (Tlsty and Coussens, 2006; Basanta et al., 2011; Nowarski et al., 2017). Specifically, it has been observed in prostate cancer that interactions with the local microenvironment are critical to the cancer progression (Basanta et al., 2011). Stromal carcinoma-associated fibroblasts extracted from invasive human breast tumors are more competent to promote the growth of breast cancers and to enhance tumor angiogenesis (Kalluri and Zeisberg, 2006). Besides, previous studies have screened several stromal molecular signatures associated with cancer progression and metastases (Binkley et al., 2004; Farmer et al., 2009). Stromal cell-derived factor-1 (SDF-1) plays roles in promoting tumor growth and enhancing angiogenesis through chemokine (C-X-C motif) receptor 4 (CXCR4) and endothelial progenitor cells in breast carcinomas (Orimo and Weinberg, 2006). Therefore, identification of the stromal molecular signatures in the reactive stroma and elucidation of tumor–stroma cross talk at the molecular level may help to improve our understanding of the progression of cancer.
In this study, we utilized the bioinformatics methods to screen the differentially expressed genes (DEGs) in the stroma surrounding invasive breast and prostate primary tumors, respectively, and performed functional analysis for DEGs. In addition, gene–chemical interactions were mapped in the Comparative Toxicogenomics Database (CTD) to screen the chemicals for the roles of feature genes in the progression of cancer. We aimed to explore potential mechanism underlying the progression of breast and prostate cancer and discover stromal molecular signatures for future cancer therapies.
2. Methods
2.1. Affymetrix microarray data
The array data of GSE26910, which was contributed by Planche et al. (2011), were obtained from National Center of Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo/), based on the platform of Affymetrix Human Genome U133 plus 2.0 Array. The data sets available in this analysis contained 24 specimens, including 6 specimens of stroma surrounding invasive breast primary tumors, 6 matched specimens of normal stromal breast tissues, 6 specimens of stroma surrounding invasive prostate primary tumors, and 6 matched specimens of normal stromal prostate tissues.
2.2. Data preprocessing and DEGs screening
The raw expression data that we downloaded had performed quartile data normalization by the robust multiarray average (Irizarry et al., 2003) algorithm of Affy2 in R/Bioconductor (Davis, 2007). The gene expression matrix of specimens was acquired based on the mean value of this gene if multiple probes corresponded to the same gene.
t-Test (Smyth, 2005) in Limma package was used to identify DEGs in the stroma surrounding invasive breast primary tumors (B.DEGs) and stroma surrounding invasive prostate primary tumors (P.DEGs), respectively. p-Value <0.01 and | log fold change (FC) | ≥ 0.5 were considered as the cutoff for DEGs screening.
The DEGs in B.DEGs and P.DEGs with the same expression pattern (high or low expression) were considered as B_P.DEGs, and these genes were feature genes involved in the interaction of tumor cell and stroma.
2.3. Protein–protein interaction network construction and functional modules analysis of network
Search Tool for the Retrieval of Interacting Genes (STRING) (Franceschini et al., 2013) database collects resource of experimental and predicted protein interactions information. The interaction pairs in STRING database were displayed with a combined score. Protein–protein interaction (PPI) network is becoming available for identifying cellular functions of proteins in various organisms (Patil and Nakamura, 2005). Cytoscape (Shannon et al., 2003) is a bioinformatics software platform for integrated models of biomolecular interaction networks. In our study, the B_P.DEGs were mapped into STRING database to construct PPI network with combined score of protein pairs not <0.8 as the cutoff. Cytoscape was used to visualize PPI network and identify highly connected nodes (Hubs). In addition, Gene Ontology (GO) (Ashburner et al., 2000) enrichment analysis is an approach for functional module studies of large-scale genomics or transcriptomics. ClusterProfiler (Yu et al., 2012) in R/Bioconductor package was used to annotate and visualize the GO function associated with biological process of modules containing B_P.DEG. Then the p-value was adjusted by the Benjamini and Hochberg method (Benjamini and Hochberg, 1995), and the adjusted p-value <0.05 was selected as the significant threshold.
2.4. Construction of gene–chemical interaction network
CTD (Davis et al., 2011, 2013) is an integrating database that describes interactions of chemical with genes/proteins, associations between chemicals and diseases, as well as diseases and genes/proteins, which promote our understanding about the effects of environmental chemicals on human health. Based on the information of CTD, the gene–chemical interactions related to “Homo sapiens” were screened to construct the gene–chemical interaction network. Notably, only the chemicals of which action manner inverse to the expression levels of feature genes (high expression vs. decrease; low expression vs. increase) in the process of their interaction were selected.
3. Results
3.1. Screening of B.DEGs, P.DEGs, and B_P.DEGs
After comparing the stroma surrounding invasive breast and prostate primary tumors with the normal samples using t-test, respectively, a total of 792 B.DEGs (352 upregulated and 445 downregulated) were identified, while 379 P.DEGs (155 upregulated and 224 downregulated) were screened. Our results showed that B.DEGs were significantly more than P.DEGs.
As shown in Figure 1, 16 overlaps (B_P.DEGs) between B.DEGs and P.DEGs were identified. Among them, 3 B_P.DEGs were upregulated (B_P.UP.DEGs) (Table 1-A) and 13 B_P.DEGs were downregulated (B_P.DOWN.DEGs) (Table 1-B). These B_P.DEGs were considered as feature genes for further analysis.
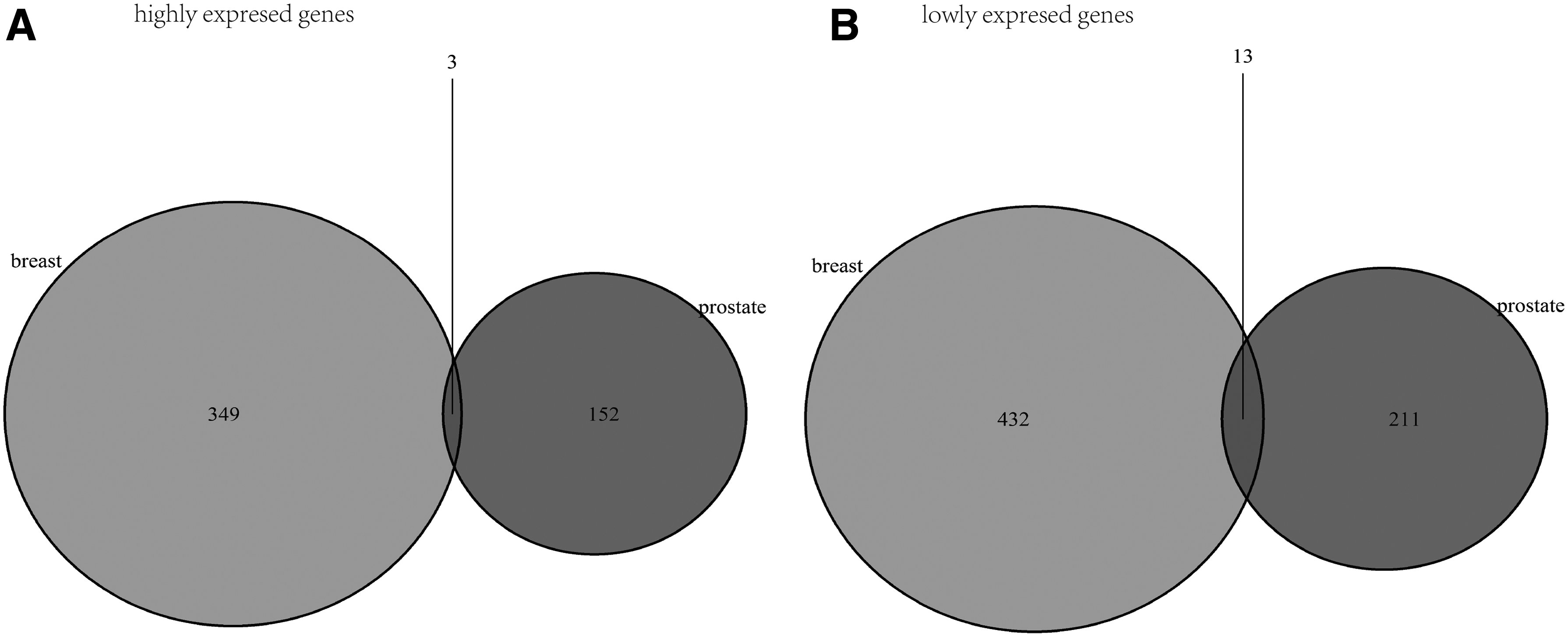
Differentially expressed genes.
Differentially Expressed Common Stroma Genes of Breast and Prostate Invasive Tumor
Symbol represents gene; Entrez gene ID represents the identification number of gene; Gene.Title represents the name of gene; FC represents fold change.
GPCR, G-protein-coupled receptor.
3.2. PPI network construction and functional modules analysis of network
Based on STRING database, only seven B_P.DEGs and their neighbors with combined score ≥0.8 were mapped into PPI network (Fig. 2). Thereinto, four B_P.DEG-coding proteins, including adenylate cyclase activating polypeptide 1 (pituitary) receptor type I (ADCYAP1R1), aspartoacylase (ASPA), glutathione S-transferase mu 5 (GSTM5) and periplakin (PPL), and their neighbors were enriched in GO terms associated with biological process for module functional analysis. Each B_P.DEG-coding protein and its neighbors were considered as a module. The results showed that ADCYAP1R1 module was significantly associated with GPCR signaling pathway (Table 2-A), ASPA module was mainly involved in amino acid metabolic process (Table 2-B), GSTM5 module was mainly involved in response to xenobiotic stimulus and glutathione metabolic process (Table 2-C), and PPL module was involved in cell differentiation and multicellular organismal process (Table 2-D). Notably, the aforementioned four feature genes were downregulated B_P.DEGs.
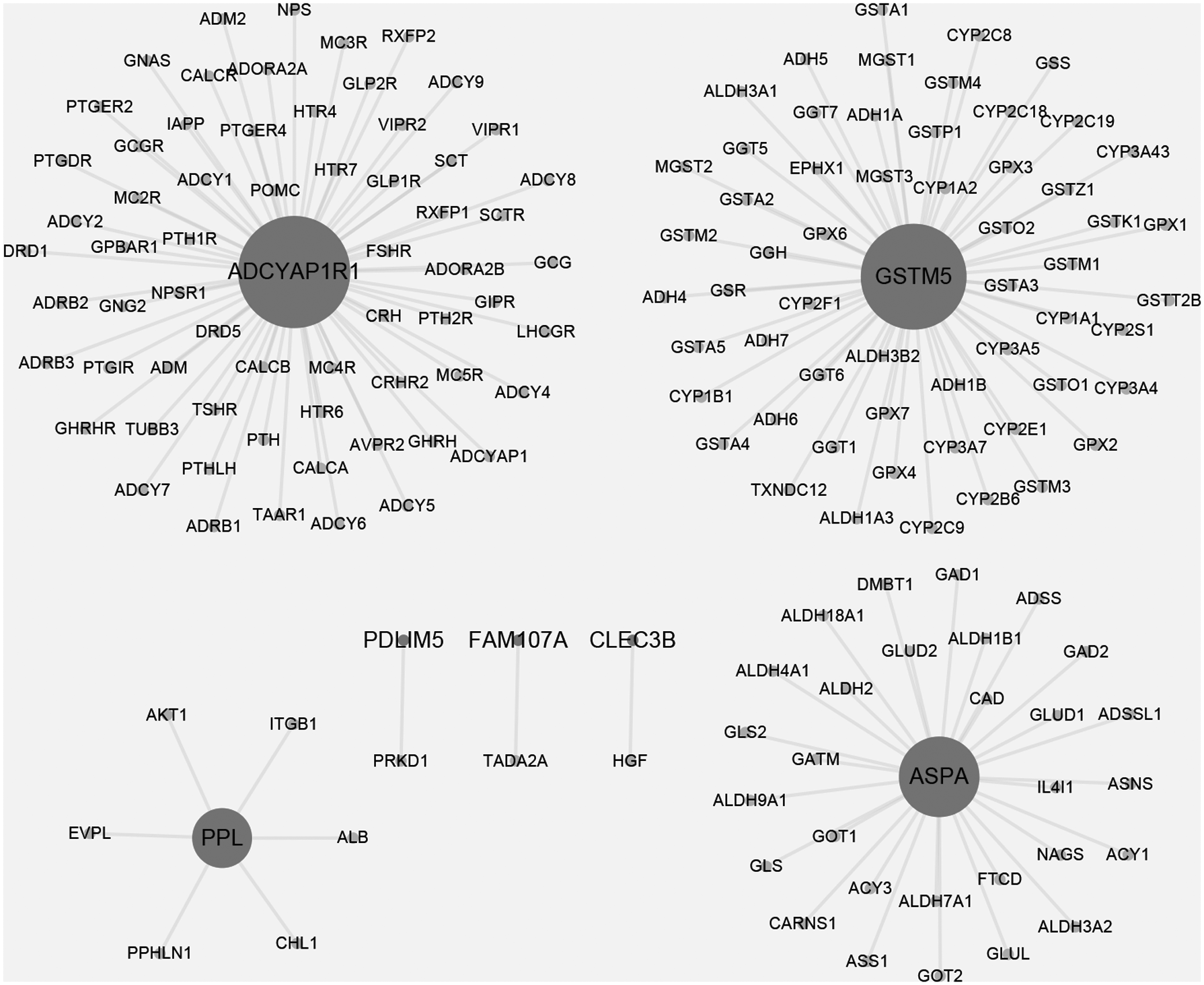
Protein–protein interaction network of common differentially expressed genes in breast and prostate stroma (B_P.DEG). Large nodes are B_P.DEGs, small nodes are neighbors of B_P.DEGs. The gray lines stand for the interaction between genes. Nodes size represents their node degree. ADCYAP1R1, adenylate cyclase activating polypeptide 1 (pituitary) receptor type I; ASPA, aspartoacylase; DEG, differentially expressed gene; GSTM5, glutathione S-transferase mu 5; PPL, periplakin.
Top Five Gene Ontology Terms Associated with Biological Process of Each Module
Modules are named by their corresponding B_P.DEGs; GO ID represents the identification number of enriched GO term associated with biological process; description represents the name of GO term; GeneRatio represents the ratio of genes enriched in GP terms; p.adjust represents significantly adjusted p-value.
DEGs, differentially expressed genes; GO, Gene Ontology; GPCR, G-protein-coupled receptor.
3.3. Interaction between feature genes and chemicals
Based on the information of CTD, the feature genes-chemical interaction network was constructed (Fig. 3). Chemicals, such as dihydrotestosterone, benzo(a)pyrene, apocarotenal, testosterone, and progesterone, interacted with feature genes directly. Thus, feature genes may play roles in cancer progression through interaction with chemicals.
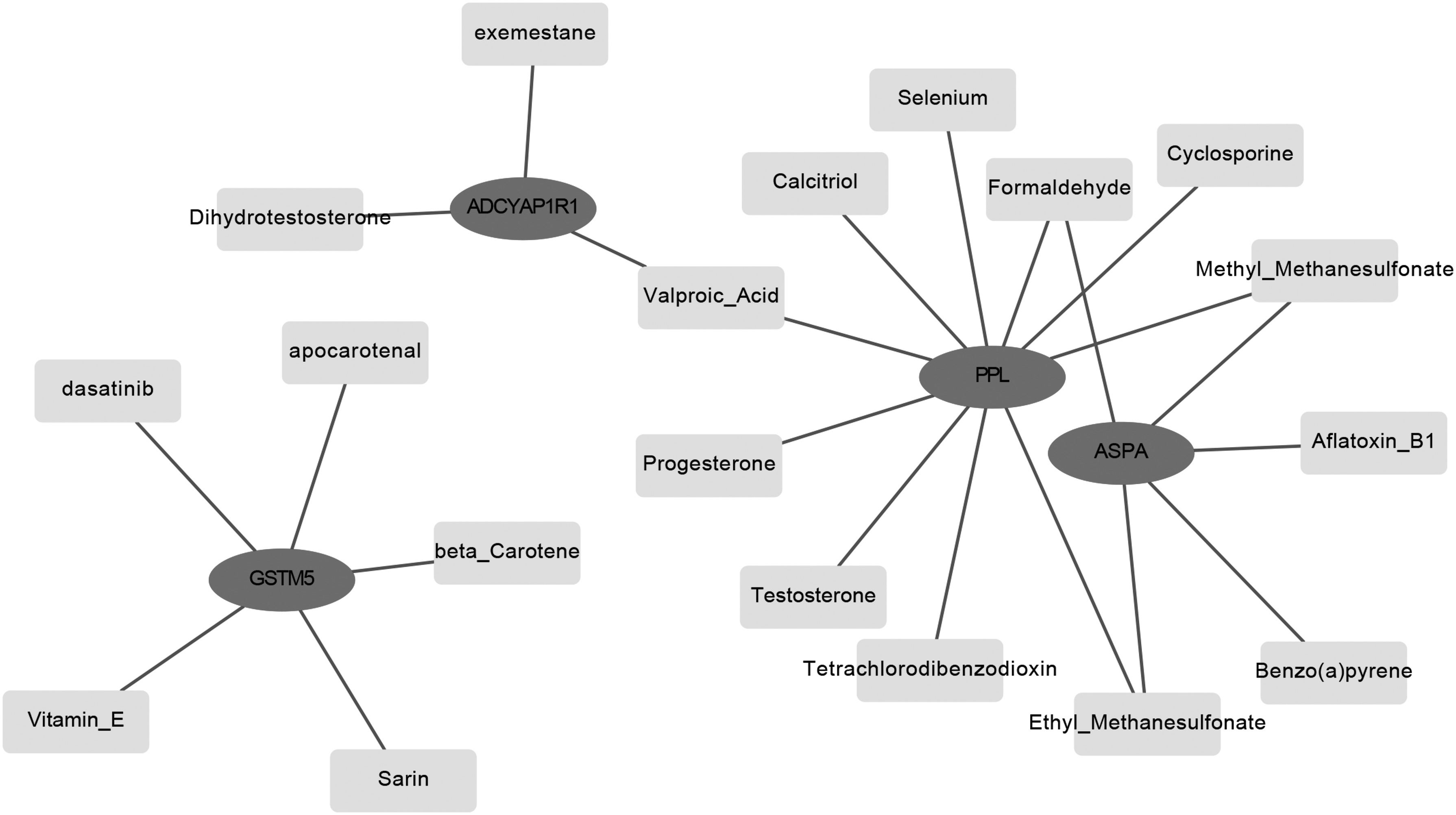
Feature genes–chemicals interactions. Ellipses represent feature genes and rectangles represent chemicals. The lines stand for the interaction of genes and chemicals.
4. Discussion
Recently, key roles of tumor stroma in cancer progression and prognosis is being disclosed, and multiple studies have highlighted the advantages of therapeutic targets of the tumor-associated stroma for anticancer therapies (Mittal et al., 2018; Mutgan et al., 2018). However, exploring novel anticancer therapies aimed at targeting stromal cells and microenvironment remains a great challenge. In this study, bioinformatics analysis was used to identify common stromal feature genes involved in the progression of breast and prostate cancer. Stromal feature genes, such as ADCYAP1R1, GSTM5, and PPL, were identified. In addition, functional modules corresponding to the aforementioned stromal genes were constructed and enriched in important biological processes associated with cancer progression. Besides, the chemicals, such as dihydrotestosterone, apocarotenal, testosterone, and progesterone, were screened based on gene–chemical interaction network.
Based on functional modules analysis, results showed that ADCYAP1R module was significantly associated with GPCR signaling pathway. GPCRs, the largest family of cell-surface molecules, play crucial roles in breast and prostate (Bhat et al., 2018; Kübler and Albrecht, 2018). In addition, G protein can couple with and activate adenylate cyclase to increase the concentrations of cyclic AMP through the stimulatory GPCR signaling pathways (Harmar, 2001). Cyclic AMP-response element-binding protein (CREB) can suppress apoptosis, induce cell proliferation, and mediate inflammation and tumor metastasis (Aggarwal et al., 2008). For example, Wnt-5a-induced phosphorylation of cAMP-regulated phosphoprotein of 32 kDa (DARPP-32) inhibits the migration of breast cancer cell in a CREB-dependent manner (Hansen et al., 2009). Overexpression of regulator of G-protein signaling 17 (RGS17) in prostate cancer induces tumor cell proliferation through the cyclic AMP-PKA-CREB pathway (James et al., 2009). Besides, ADCYAP1R1, a membrane-associated protein, is capable of signaling and regulating intracellular concentrations of cyclic AMP through coupling to adenylate cyclase (Harmar, 2001). Therefore, our results suggest that ADCYAP1R1 may be a feature stromal molecule involved in the progression of breast and prostate cancer through regulating concentrations of cyclic AMP mediated by GPCR signaling pathway.
In contrast, our results showed that dihydrotestosterone, a chemical, interacted with ADCYAP1R1 directly in gene–chemical interaction network. Dihydrotestosterone is used for androgen receptor activation in prostate cancer (Titus et al., 2005), and has effects on inhibiting breast cancer cell proliferation (Macedo et al., 2006). In addition, it has been reported that dihydrotestosterone decreases tumor necrosis factor-α (TNF-α) and regulates inflammatory response mediated by nuclear factor-κB (NF-κB) in human endothelial cells (Titus et al., 2005). TNF-α can promote prostate cancer cell migration through upregulation of CCR7 (Maolake et al., 2018). TNF-α promotes breast cancer growth through the positive feedback loop of TNFR1/NF-κB (and/or p38)/p-STAT3/HBXIP/TNFR1 (Cai et al., 2017). Thus, we speculate that dihydrotestosterone is a key chemical and could be used for studying the roles of ADCYAP1R1 in breast and prostate cancer progression according to the interaction of dihydrotestosterone and ADCYAP1R1.
GSTM5 module was mainly involved in glutathione metabolic process in this study. The glutathione content of cancer cells is particularly relevant in regulating DNA synthesis, growth, mutagenic mechanisms, and radiation resistance (Estrela et al., 2006). GSTM5 belongs to glutathione S-transferases (GSTs), which is widely involved in glutathione metabolic process and plays regulatory roles in cellular signaling by forming PPIs with critical kinases involved in controlling apoptosis and proliferation (McIlwain et al., 2006; Lo and Ali-Osman, 2007).
Several molecular epidemiological studies have examined the relation between breast cancer risk and GST genetic polymorphisms (Mitrunen et al., 2001). The hypermethylation of the GSTπ regulatory region is the most common somatic alteration identified in human prostate cancer (McIlwain et al., 2006). Although roles of GSTM5 have not been fully discussed, based on the studies, we speculate GSTM5 may play roles in the progression of breast and prostate cancer through glutathione metabolic process. In contrast, previous study has confirmed that GSTM5 is upregulated by apocarotenal treatment (Goralczyk, 2009). Apocarotenals can be produced by asymmetric cleavage of β-carotene and could inhibit PPARγ and PPARα activation (Ziouzenkova et al., 2007). PPARγ is found aberrantly expressed in most breast tumors, and inhibiting endogenous PPARγ signaling may be a promising approach to breast cancer therapy (Zaytseva et al., 2011). Thus, apocarotenals is strongly associated with breast cancer progression. In addition, apocarotenals inhibit the transcriptional activity of NF-κB, which is an important factor involved in prostate cancer earlier (Kong et al., 2007). Besides, apocarotenal could interact with GSTM5 directly based on the analysis of gene–chemicals interaction network. Thereby, our results also suggest that GSTM5 may involve in the progression of breast and prostate cancer through the stimulation of apocarotenals.
PPL module was involved in cell differentiation. PPL as a localization signal plays a role in the protein kinase B (PKB)-mediated signaling (van den Heuvel et al., 2002). PKB is a major player in various essential growth factor-induced cellular processes, such as cell cycle regulation and cell survival (Brazil and Hemmings, 2001). In addition, PKB acts downstream of the lipid kinase phosphoinositide 3-kinase (PI3K) and plays a role in tumor progression (Tsatsanis and Spandidos, 2000; Vivanco and Sawyers, 2002). PI3K pathway acts as hormone resistance in breast cancer (Tokunaga et al., 2006) and is involved in the prostate cancer cell proliferation and metastasis (Xu et al., 2017). In addition, testosterone and progesterone can activate PI13K/Akt signaling, and thus play roles in the progression of breast and prostate cancer (Tokunaga et al., 2006; Kim et al., 2014). This is in line with our results that PPL can interact with testosterone and progesterone directly. Therefore, PPL may be an important stromal molecule playing roles in the apoptosis and proliferation of breast and prostate cancer cells through PKB-involved PI3K signaling pathway.
Taken together, our study indicates that stromal feature genes, such as ADCYAP1R1, GSTM5, and PPL, are stromal molecular signatures and may play a key role in the progression of breast and prostate cancer. ADCYAP1R1 could promote cancer progression through GPCR signaling pathway, GSTM5 may promote cancer progression by glutathione metabolic process, and PPL may be involved in cancer progression through PI3K signaling pathway. Moreover, chemicals, such as dihydrotestosterone, apocarotenal, testosterone, and progesterone, could target the aforementioned stromal feature genes and may make great contributions to the progression of breast and prostate cancer. These findings shed new light on the molecular mechanism of cancer progression and have implications for future research. However, there are no experiments performed in our study, and more genetic experiments of DEGs are still needed to confirm the possible stromal molecules.
Footnotes
Acknowledgment
This study was supported by Shanghai Medical Key Specialty (No. ZK2015A25).
Author Disclosure Statement
The authors declare there are no competing financial interests.