
Abstract
Morphine tolerance is one of the most common complications in patients with chronic pain. Many patients with morphine tolerance have poor efficacy in the treatment of primary pain, and are accompanied by the side effects. Previous studies have found that many mechanisms are involved in morphine tolerance, but few researches could fully explain morphine tolerance, and no effective treatment for morphine tolerance has been found. One expression profiling data set was downloaded from the Gene Expression Omnibus (GEO) database. The probes would be transformed into the homologous gene symbol by means of the platform's annotation information. GEO2R was used to search for differentially expressed long noncoding RNAs (lncRNAs) and differentially expressed genes (DEGs) that were differentially expressed between spinal cord samples. Receiver operator characteristic curve analysis was performed to determine the ability of the hub lncRNAs to predict morphine tolerance. Through the principal component analysis, the intragroup data repeatability is fine in the GSE110115. A total of 10 genes were identified as hub genes from the protein–protein interaction network with degrees ≥10. Compared with the normal saline group, the expression levels of LncRNA XR_006440, XR_009493, AF196267, MRAK150340, and MRAK037188 were more downregulated, while the expression levels of MRAK046606, XR_005988, DQ266361, uc.167−, and uc.468+ were more upregulated in the morphine tolerance group. LncRNAs and DEGs were differentially expressed between the morphine tolerance group and nonmorphine tolerance group, which may be involved in the development of morphine tolerance, especially LncRNA DQ266361, uc.167−, and Mmp9, CCL7 genes.
Introduction
Morphine tolerance is a decrease in drug efficacy due to prolonged or heavy use, or a higher dose of morphine is needed to produce the same analgesic effect, while the increase of drug dose will further maintain or even aggravate morphine tolerance, forming a vicious cycle. With the increase of patients with cancer pain and neuropathic pain, opioids are widely used as powerful analgesics. Abuse is accompanied by many side effects, particularly morphine tolerance. At present, many patients with morphine tolerance have poor efficacy in the treatment of primary pain, and are accompanied by the side effects of drug dependence, addiction, anxiety, depression, and serious decline in quality of life. As a result, more and more related studies have been conducted (Kalso et al., 2004; Volkow and McLellan, 2016). Mechanisms of morphine tolerance include the opioid receptor desensitization theory, receptor downregulation theory, and receptor subtype theory, and glial cells activate associated neuroinflammation theory (Raval et al., 2003; Watkins et al., 2009; He et al., 2011), while these results cannot fully explain the mechanism of morphine tolerance.
As important noncoding RNA, long noncoding RNAs (lncRNAs) are involved in many important physiological activities of the body. Salmena et al. (2011) proposed the CeRNA theory to explain the lncRNA action pathway, that is, lncRNA acts on the downstream miRNA, affecting its gene silencing effect and thus affecting gene expression. Jiang et al. (2015) believe that lncRNA acts on surrounding homologous coding genes. Although the specific action mechanism of lncRNA remains to be studied, a large number of studies have found that lncRNA is involved in the occurrence and development of a variety of diseases by affecting the expression of target mRNA. Wang et al. (2018) found that lnc-TSI combines with MH2 domain of Smad3 and acts on TGF-β/Smad pathway to participate in the fibrosis of chronic kidney disease. Bester et al. (2018) found that transcriptional activation of GAS6-as2 lncRNA led to excessive activation of GAS6/TAM pathway, which was involved in drug tolerance of various tumors. Hu et al. (2019) found that lncRNA PKIA-ASI is involved in the neuropathic pain process by affecting CDK6 expression, suggesting that it may be a potential therapeutic target. Recently, it has been found that noncoding RNA is also involved in the occurrence and maintenance of morphine tolerance and plays an important role in it. However, the study on the relationship between lncRNAs and morphine tolerance is still in the preliminary stage and has not been clarified.
Bioinformatics is a hot technology about the application of biological big data mining and analysis. Through this technology, the differentially expressed genes (DEGs) and noncoding RNA of patients with diseases and normal people can be analyzed from the public big data, and further screening can help researchers find the most closely related parts of diseases. Bioinformatics has proven to be a promising technology for the exploration of disease biomarkers and targeted therapies (Sun et al., 2019).
Therefore, this study aims to screen and identify differentially expressed lncRNA (DER) and DEGs related to morphine tolerance by mining the public database with microarray technology. Through further data analysis, we found 10 lncRNA and 10 DEGs that were more likely to be involved in the occurrence and maintenance of morphine tolerance. This study provides a basis for further exploring the mechanism of morphine tolerance and targeted therapy.
Materials and Methods
Access to public data
The Gene Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo) is an open functional genomic database of high-throughput resource, including microarrays, gene expression data, and chips (Edgar et al., 2002). One expression profiling data set GSE110115 (GPL15690; Arraystar Rat LncRNA microarray) was downloaded from the GEO database. The probes would be transformed into the homologous gene symbol by means of the platform's annotation information. The GSE110115 data set contained five normal spinal cord samples treated with normal saline and five spinal cord samples from morphine tolerance rats.
Intragroup data repeatability test
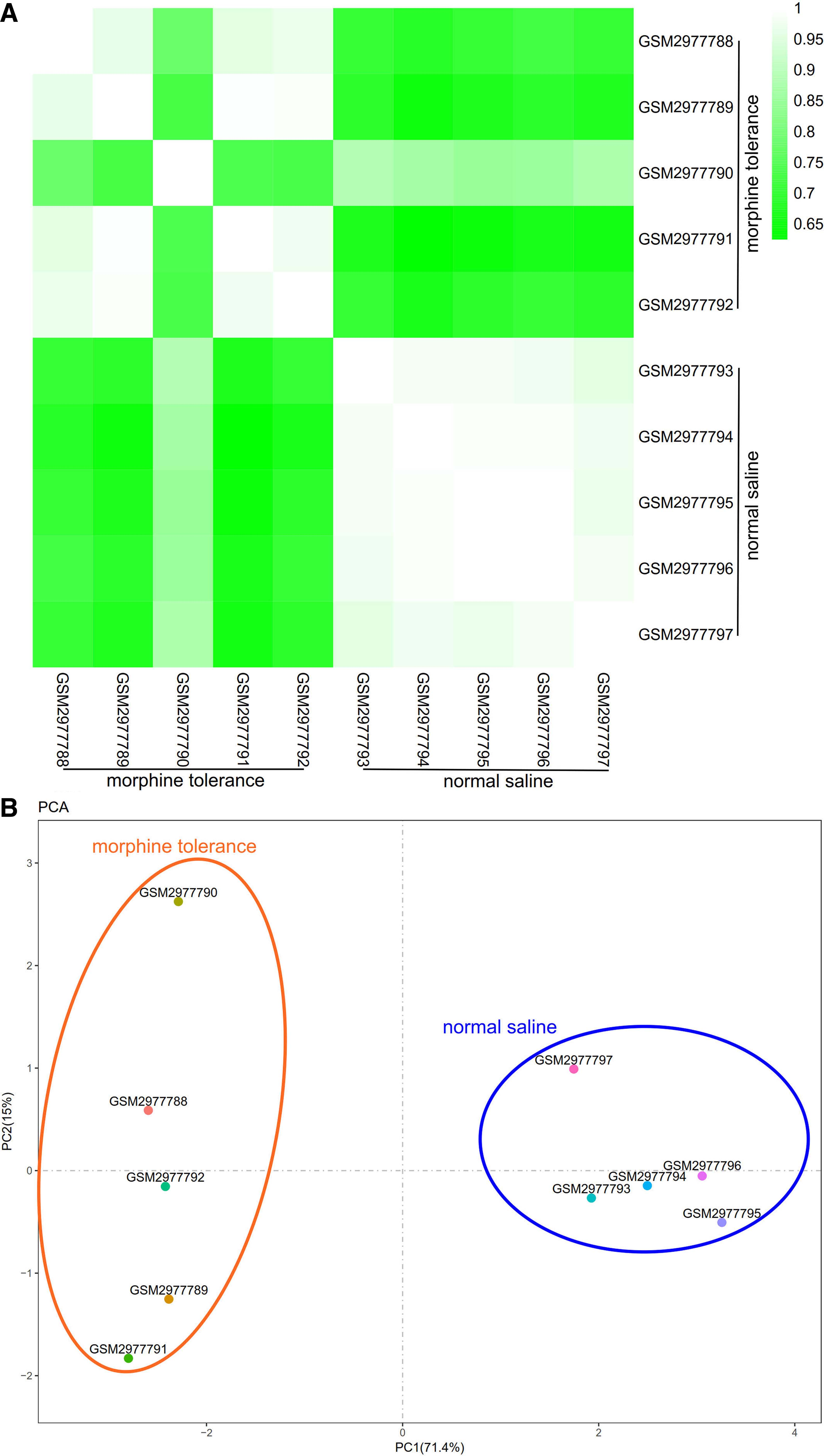
The Pearson's correlation test was performed to verify the intragroup data repeatability in each group. The R programming language is the software and operating environment for statistical analysis and drawing. The correlation between every sample from the same data set was visualized by the heat maps that were drawn with the R programming language. Principal component analysis (PCA) is a common method for sample clustering, which is often used for gene expression, diversity analysis, resequencing, and other sample clustering based on various variable information. Furthermore, the intragroup data repeatability of each data set was tested by sample clustering analysis.
Identification of differentially expressed lncRNAs and differentially expressed genes
GEO2R (www.ncbi.nlm.nih.gov/geo/geo2r) is an interactive online tool to identify DERs and DEGs by comparing samples from the GEO series (Barrett et al., 2013). GEO2R was used to search for DERs and DEGs that were differentially expressed between normal spinal cord samples and morphine tolerance spinal cord samples. The cutoff criteria were a p-value of <0.05. The Venn plots were used to intersect the two data sets to obtain the common DERs. The volcano maps were drawn using the volcano plotting tool (https://shengxin.ren). Then, the DEGs were screened and cited from former research (Shao et al., 2018).
Construction and analysis of protein–protein interaction network and identification of significant module
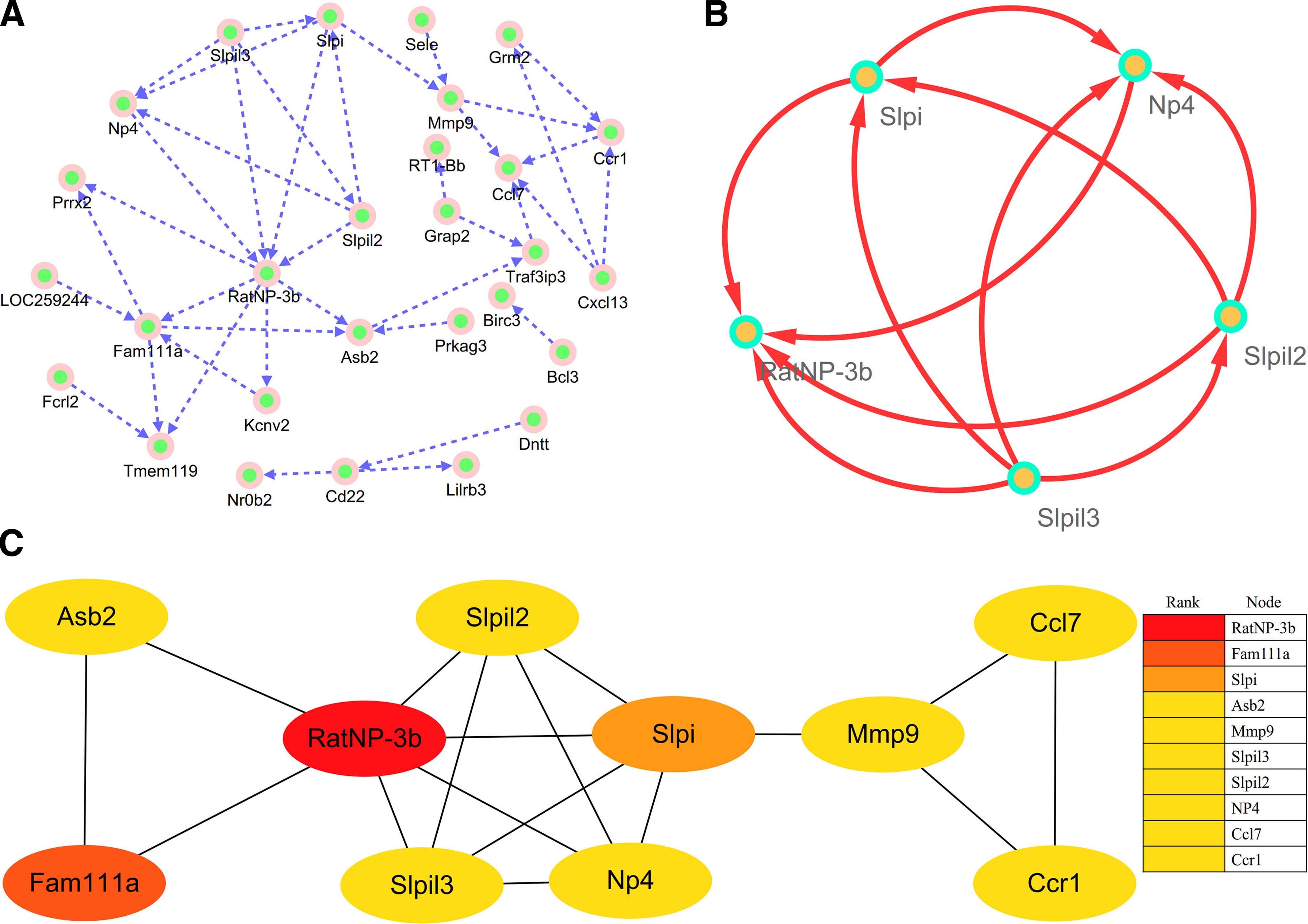
The STRING online database (http://string-db.org) could predict and trace out the protein–protein interaction (PPI) network after importing the common DEGs to Search Tool for the Retrieval of Interacting Genes. The STRING database was used for construction of PPI network of DEGs. Cytoscape (version 2.8) (Smoot et al., 2011), a free visualization software, was applied to visualize PPI networks. Next, the Molecular Complex Detection (MCODE) (version 1.5.1, a plug-in of Cytoscape) identified the most important module of the network map. The criteria for MCODE analysis are degree cutoff = 2, MCODE scores >5, Max depth = 100, node score cutoff = 0.2, and k-score = 2 (Bader and Hogue, 2003).
Identification and analysis of hub genes
When the degrees were set (degrees ≥10), the hub genes were excavated. DAVID (https://david.ncifcrf.gov/home.jsp) (version 6.8) is one online analysis tool suite with the function of Integrated Discovery and Annotation (Huang et al., 2007). Gene Ontology (GO) is an ontology widely used in bioinformatics, which covers three aspects of biology, including biological process (BP), cellular component (CC), and molecular function (MF) (Ashburner et al., 2000). Kyoto Encyclopedia of Genes and Genomes (KEGG) (https://www.kegg.jp) is one of the most commonly used biological information databases in the world. To analyze GO and biological pathway information of DEGs, DAVID online tool was implemented. The rule of statistical significance is p < 0.05. Furthermore, The Biological Networks Gene Oncology tool (BiNGO) (version 3.0.3) was used to analyze and visualize the hub genes' CC, BP, and MF (Maere et al., 2005).
Analysis of hub long noncoding RNAs
The hub lncRNAs were excavated from the former study via obtaining the top 5 upregulated lncRNAs and the top 5 downregulated lncRNAs (Shao et al., 2018). Subsequently, one hierarchical clustering heat map of hub lncRNA expression was visualized with R programming language. Correlation analysis between the hub lncRNAs was performed. For correlation analysis between morphine tolerance and relevant lncRNA expression, the Pearson's correlation coefficient was used. Any test results reaching a liberal statistical threshold of p < 0.2 for each comparison were then entered into a univariable linear regression model, to identify independently predictive lncRNA of morphine tolerance. Finally, we performed receiver operator characteristic (ROC) curve analysis to determine the ability of the hub lncRNAs to predict morphine tolerance. All statistical analyses were conducted using SPSS software (version 21.0; IBM). A p-value of less than 0.05 was considered statistically significant.
Results
Validation of the data sets
To further validate the intragroup data repeatability, we used the Pearson's correlation test and PCA. Based on the Pearson's correlation test, we found that in the GSE110115, there is a strong correlation among samples in the normal saline group, and there also is a strong correlation among samples in the morphine tolerance group (Fig. 1A). Through the PCA, the intragroup data repeatability is fine in the GSE110115. The distances between each sample in the normal saline group were close, and distances between each sample in the morphine tolerance group were also close in the dimension of PC1 (Fig. 1B).
After analysis of the data sets (GSE110115) with GEO2R, the difference between normal saline and morphine tolerance groups could be presented in the volcano plots (Fig. 2).
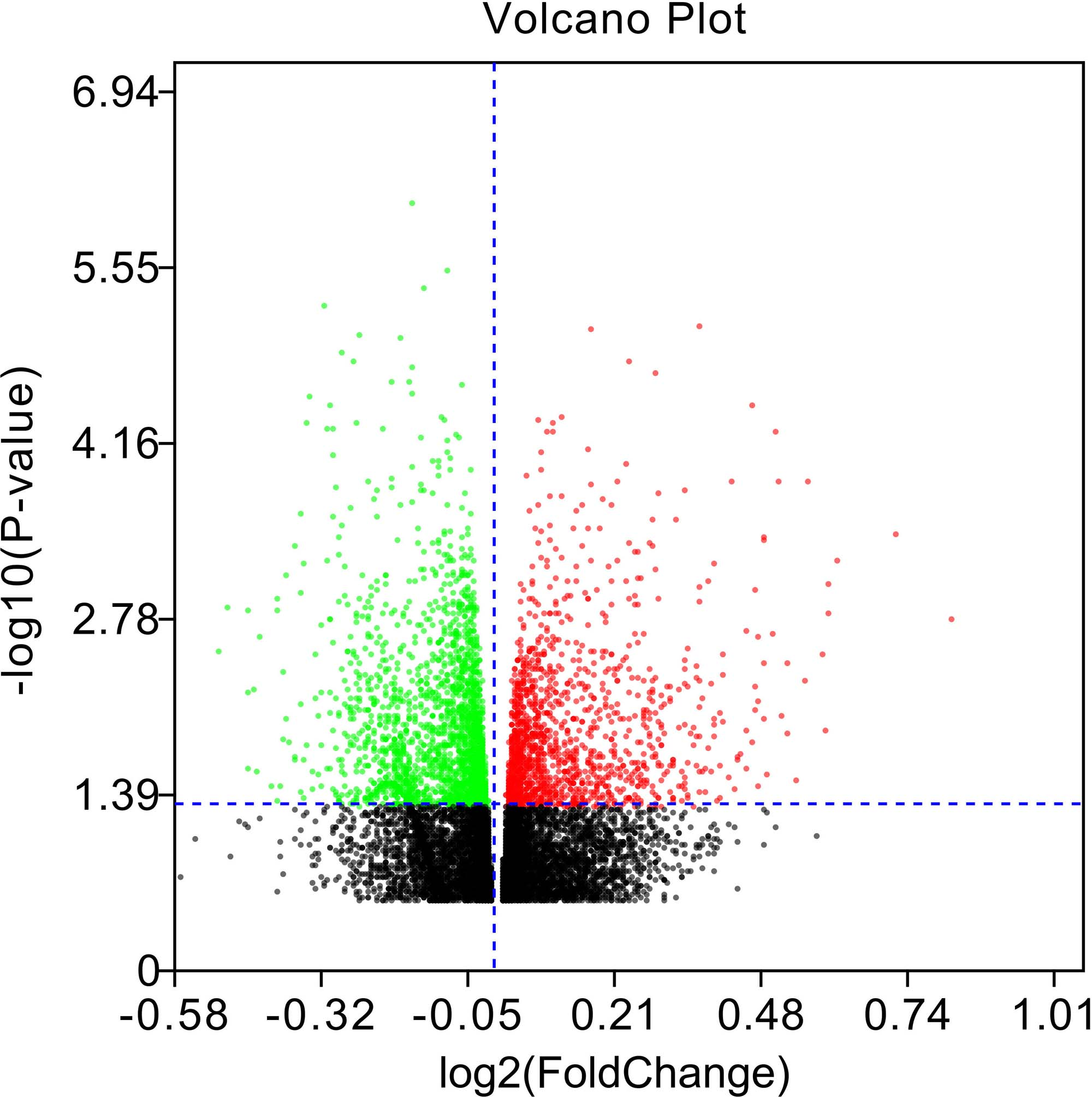
The identification of DERs between morphine tolerance and normal saline samples. The volcano plot presents the difference between morphine tolerance and normal saline samples after analysis of the data set GSE110115 with GEO2R. DERs, differentially expressed long noncoding RNAs.
The PPI network of DEGs was constructed (Fig. 3A), and the most significant module was obtained from PPI network by using Cytoscape (Fig. 3B). A total of 10 genes (RatNP-3b, Fam111a, Slpi, Asb2, Mmp9, Slpil3, Slpil2, NP4, Ccl7, and Ccr1) were identified as hub genes from PPI network with degrees ≥10 (Fig. 3C). The results of GO analysis presented that variations in BPs, CC, and MF of hub genes were mainly enriched in killing of cells of other organisms, negative regulation of endopeptidase activity, defense response to fungus, negative regulation of viral genome replication, antibacterial humoral response, chemokine-mediated signaling pathway, cellular response to interleukin-1, extracellular space, extracellular region, endopeptidase inhibitor activity, and peptidase activity (Table 1). Analysis of KEGG pathway displayed that the all hub genes were primarily enriched in the chemokine signaling pathway and cytokine–cytokine receptor interaction (Table 1). According to GeneCards, summaries for the function of the 10 hub genes were obtained (Table 2). What is more, the BP, CC, and MF analysis for these hub genes is presented based on the BINGO software (Fig. 4A–C).
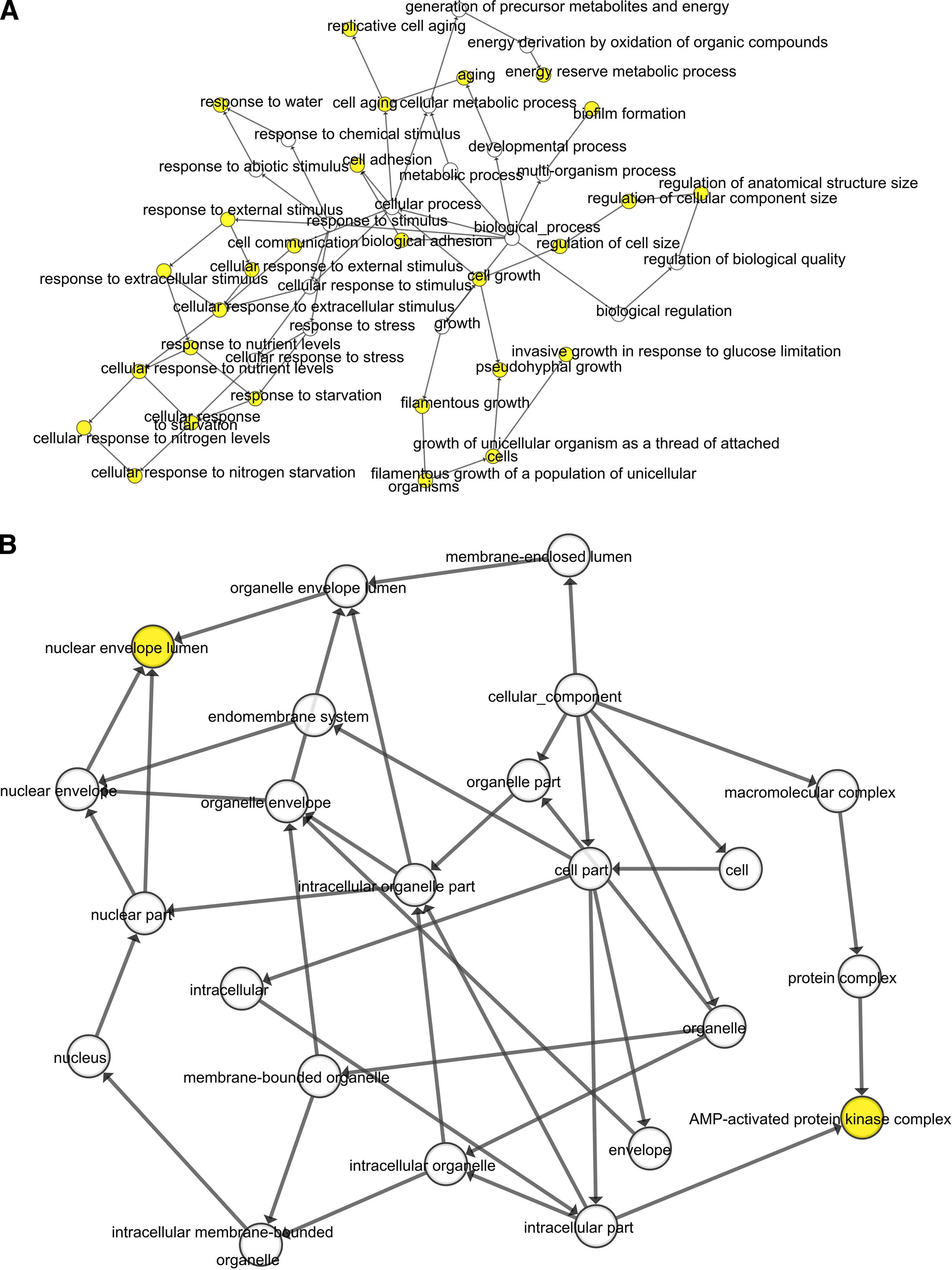
The biological process
Gene Ontology and Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analysis of Hub Genes Between Normal and Morphine Tolerance Groups
GO, Gene Ontology.
Summaries for the Function of 10 Hub Genes
Hierarchical clustering could basically differentiate the morphine tolerance group from normal saline group via the expression levels of hub lncRNAs in the data set GSE110105. Compared with the normal saline group, the expression levels of XR_006440, XR_009493, AF196267, MRAK150340, and MRAK037188 were more downregulated, while the expression levels of MRAK046606, XR_005988, DQ266361, uc.167−, and uc.468+ were more upregulated in the morphine tolerance group (Fig. 5). The heat map showed the correlation between hub lncRNAs in the data set GSE110105. There exit strongly positive correlations between AF196267, XR_009493, MRAK150340, MRAK037188, and XR_006440. And there exist strongly negative correlations between XR_005988, DQ266361, MRAK046606, uc.167−, and uc.468+ (Fig. 6).

Hierarchical clustering showed that the hub genes could basically differentiate the morphine tolerance samples from the normal saline samples in the data sets GSE110115. Upregulation of genes is marked in red; downregulation of genes is marked in green.
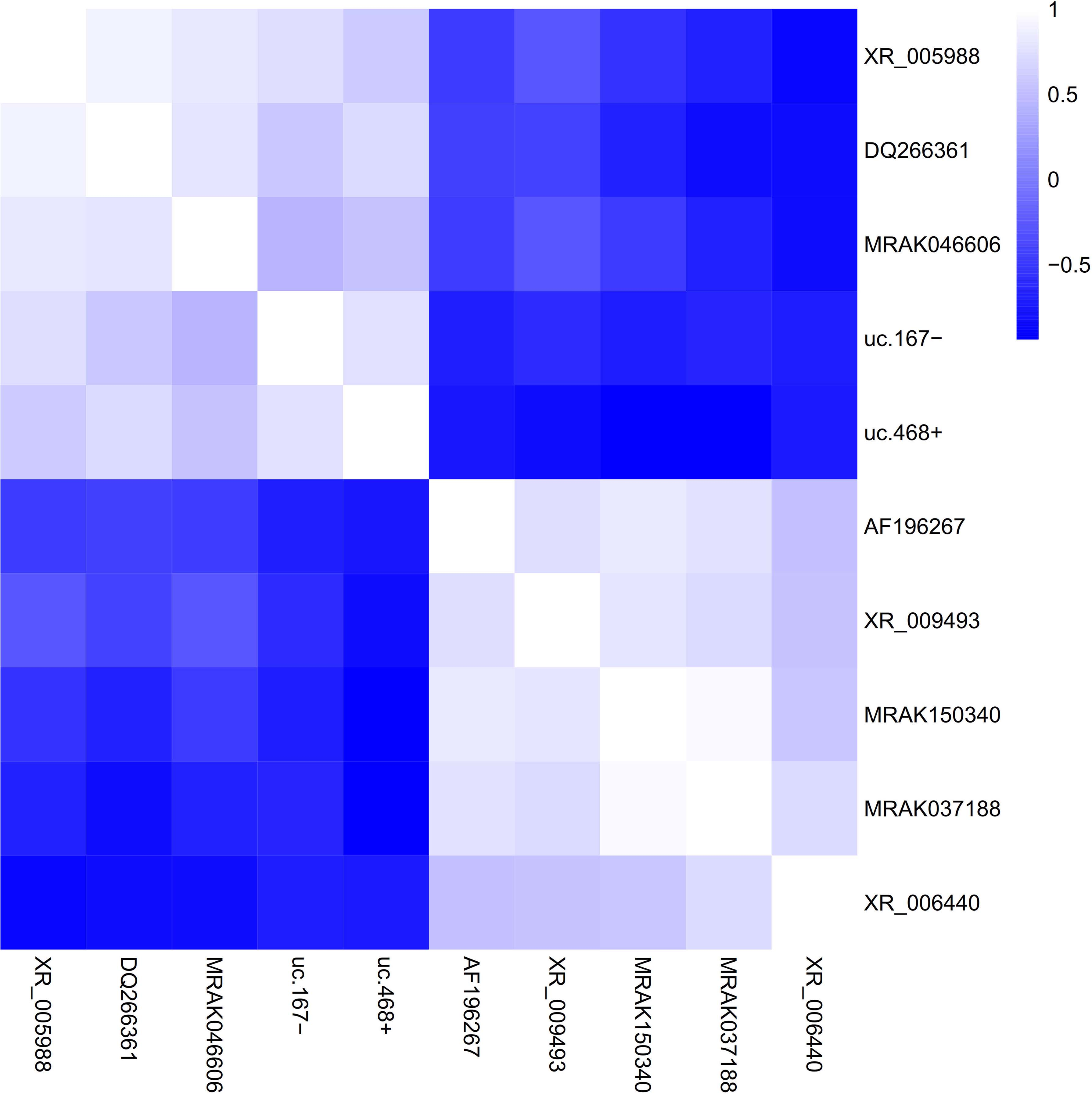
Heat maps presenting the correlations between hub lncRNAs in the GSE110115. The color means the intensity of correlation. When 0 < correlation <1, there exits the positive correlation. When −1 < correlation <0, there exits the negative correlation. And the bigger the absolute value of a number is, the stronger the correlation is. lncRNAs, long noncoding RNAs.
To ensure that the hub lncRNAs had an impact on morphine tolerance, we performed a further analysis of morphine tolerance and hub lncRNA expressions. Pearson's correlation coefficient was used in the correlation analysis, and XR_005988 (ρ = 0.820, p = 0.004), DQ266361 (ρ = 0.876, p = 0.001), MRAK046606 (ρ = 0.714, p = 0.020), uc.167− (ρ = 0.830, p = 0.003), uc.468+ (ρ = 0.911, p < 0.001), AF196267 (ρ = −0.752, p = 0.012), XR_009493 (ρ = −0.719, p = 0.019), MRAK150340 (ρ = −0.906, p < 0.001), MRAK037188 (ρ = −0.936, p < 0.001), and XR_006440 (ρ = −0.825, p = 0.003) were significantly correlated with morphine tolerance (Table 3). In the univariate linear regression model, holding all other variables at any fixed value, the natural logarithmic morphine tolerance remained associated with all the hub lncRNAs (p < 0.05) (Table 3).
The Correlation and Linear Regression Analysis Between Morphine Tolerance and Relevant Long Noncoding RNA Expression
The Correlation and Linear Regression Analysis Between Morphine Tolerance and Relevant Long Noncoding RNA Expression
Pearson's correlation coefficient between MT and relevant characteristics; ρ: Pearson's correlation coefficient.
Univariate linear regression analysis, β: parameter estimate.
Significant variables: p < 0.05.
lncRNA, long noncoding RNA; MT, morphine tolerance.
To identify accurate threshold for hub lncRNAs predicting morphine tolerance, we constructed ROC curves. The expression of all the hub lncRNAs was associated with diagnosis of morphine tolerance (0.8 < AUC <1, p ≤ 0.05) (Table 4 and Fig. 7).
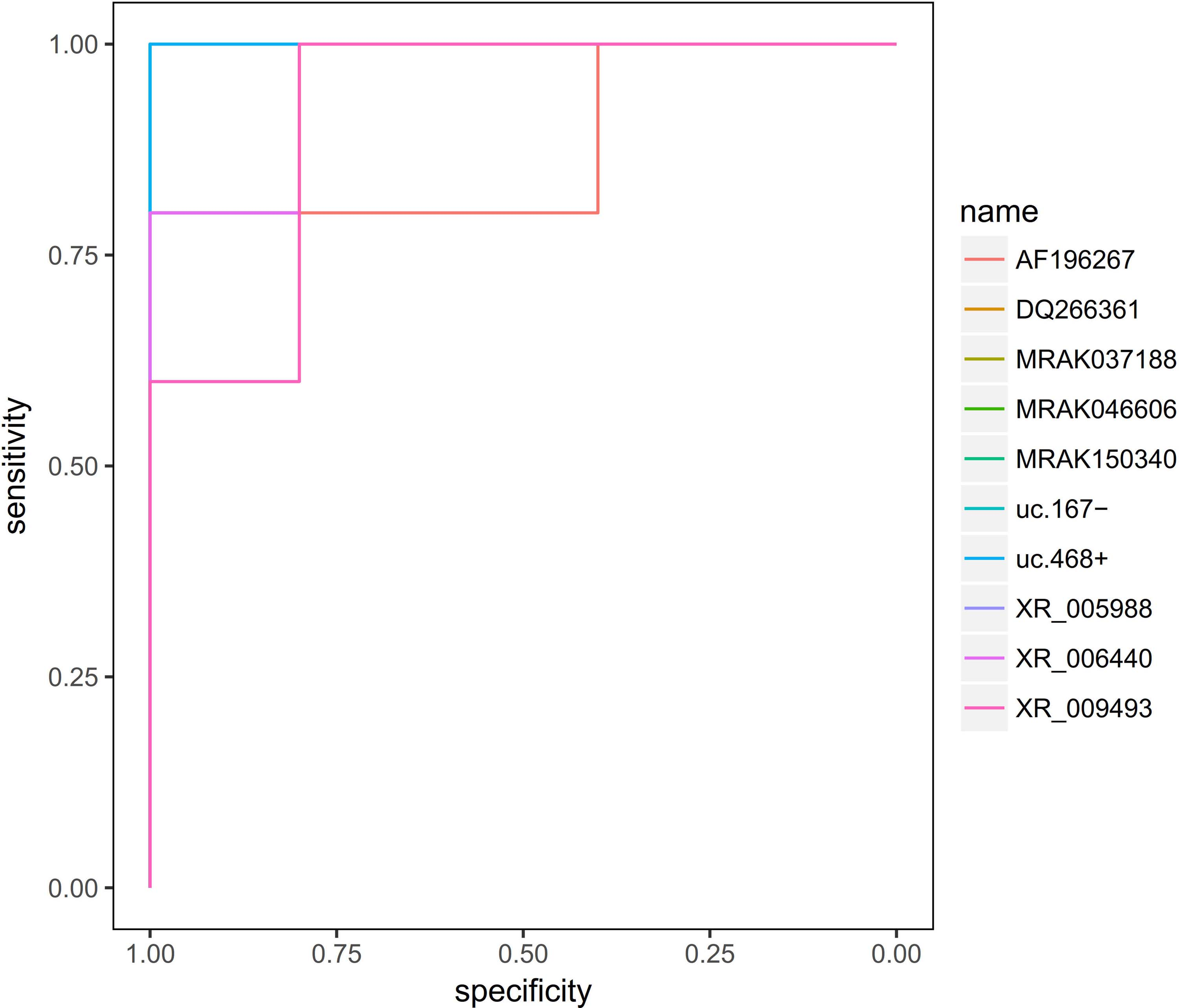
The receiver operator characteristic curves indicating that the all hub lncRNAs could predict morphine tolerance sensitively and specially.
Receiver Operator Characteristic Curve Analysis of Long Noncoding RNA Expression for Morphine Tolerance
Significant variables.
AUC, area under curve; CI, confidence interval; MT, morphine tolerance; ODT, optimal diagnostic threshold.
Morphine tolerance is one of the most common complications in patients with chronic pain. In addition to the tolerance of analgesic effect, related side effects, such as constipation and respiratory depression, are not tolerated, causing a large number of patients with chronic pain to die from side effects caused by a high dose of morphine. Also, deaths from the abuse of drugs for euphoria, such as fentanyl, are on the rise (Roxburgh et al., 2017). Therefore, it is urgent to find out the mechanism of morphine tolerance and effective treatment methods. Previous studies have found that many mechanisms are involved in morphine tolerance, but none can fully explain morphine tolerance, and no effective treatment for morphine tolerance has been found. Corder et al. (2017) found that removing the peripheral μ opioid receptor can eliminate morphine tolerance without affecting analgesia, but the exact molecular mechanism was not elucidated. Huang et al. (2019) found that mir-873a-5p was involved in morphine tolerance, and abnormal expression of lncRNA (Shao et al., 2018) was found in multiple morphine tolerance rats by microsequencing, suggesting the significance of noncoding RNA in morphine tolerance studies.
Bioinformatic technology has been widely used to screen tumor target genes, which is a reliable method to explore the mechanism and find therapeutic targets. We used this technique to explore the DEGs and lncRNAs associated with morphine tolerance. Zhou et al. (2017) found abnormal expression of various lncRNAs and mRNAs in the spinal cord of neuropathic pain rats by RNA sequencing. Furthermore, the function of DERs and target genes was predicted by GO and KEGG biological pathway analysis. Finally, the lncRNA and gene that are most likely to be involved in neuropathic pain were identified, providing a new way to study the mechanism of neuropathic pain (Zhou et al., 2017).
We identified 10 lncRNAs (XR_005988, DQ266361, MRAK046606, uc.167−, uc.468+, AF196267, XR_009493, MRAK150340, MRAK037188, and XR_006440) and 10 genes (RatNP-3b, Fam111a, Slpi, Asb2, Mmp9, Slpil3, Slpil2, NP4, Ccl7, and Ccr1) most associated with morphine tolerance through data mining analysis.
DQ266361 is one of the lncRNAs with the most statistically significant change in p-value, which is highly expressed in the spinal cord of morphine-tolerant rats. However, we have not found relevant research articles through PubMed and EMBASE search. Whereas we boldly speculate that this lncRNA is involved in morphine tolerance, and the relevant mechanism is worth further exploration.
Uc.167− has been extensively studied in myocardial development. By screening and verifying the LncRNA associated with ventricular septal defect, Song et al. (2016) found that uc.167− was involved by regulating the genes encoding downstream cardiac myocytes, suggesting its potential therapeutic target action. Yin et al. (2018) discovered that this lncRNA is involved in heart development by regulating the methylation state of target genes, and further discovered that it's possible BP is Rap1 signaling pathway. We found that uc.167− was highly expressed in morphine-tolerant rats compared with normal rats by the bioinformatic method. We speculate that uc.167− may be a potential therapeutic target by affecting a signaling pathway, regulating downstream target genes, and thereby participating in the occurrence and development of morphine tolerance.
Mmp9 (matrix metallopeptidase 9) is an important extracellular protease, which is mainly involved in the signaling pathway between cells, the phosphorylation of important proteins, and the regulation of synaptic plasticity. Abnormal activation of Mmp9 is involved in the pathophysiological process of various diseases (Lepeta and Kaczmarek, 2015). Nagy et al. (2006) found that abnormal activation of Mmp9 led to continuous depolarization of neurons and changes in synaptic plasticity in hippocampal functional areas, and thus participated in the occurrence of Alzheimer's disease. Zhao et al. (2017) found that Mmp9 release after nerve injury was involved in the activation of microglia cells, and then involved in neuropathic pain. Synaptic plasticity and glial activation may also be mechanisms of morphine tolerance. Some scholars began to explore the relationship between Mmp9 and morphine tolerance. Existing studies have found that Mmp9 is continuously activated in the superficial dorsal horn of spinal cord and dorsal root ganglion after morphine treatment. In particular, Mmp9 activation in the dorsal root ganglia may be involved in the development of morphine tolerance (Nakamoto et al., 2012; Zhang et al., 2017). Consistent with existing studies, we found high expression of MMP9 in the spinal cord of morphine-tolerant rats through bioinformatic analysis, suggesting that it may be involved in the development of morphine tolerance. We speculate that Mmp9 is involved in morphine tolerance by influencing spinal cord synaptic plasticity and glial cell activation-related neuroinflammation. Mmp9 may be a potential therapeutic target for morphine tolerance.
CCL7 is a chemokine involved in inflammation. Chemokines play an important role in maintaining the stability of the internal environment in the body's immune response, and mediate the migration of leukocytes to play an important role in the immune response and inflammatory response (Kimura et al., 2014). CCL7 regulates toll-like receptor (TLR) signaling pathways associated with a variety of inflammation, including infection, cardiovascular disease, and tumor microenvironment (Curtale et al., 2018). Zouggari et al. (2013) found that CCL7 may be a target for the treatment of acute myocardial infarction. Li et al. (2017) found that reducing CCL7 in the spinal cord can inhibit the activation of microglia and alleviate neuropathic pain. Through bioinformatic data analysis, we found that CCl7 was highly expressed in the spinal cord of morphine-tolerant rats, suggesting that it may be involved in morphine tolerance and serve as a therapeutic target. Corbisier et al. (2015) found that chemokine ligands are involved in recruitment of arrestin, and different chemokines bind to ligands to participate in the selective activation of the G-protein-coupled receptor signaling pathway, while G-protein is preferred to agonist, which is believed to play a better analgesic effect and reduce the occurrence of side effects (AAH et al., 2019). In summary, we speculated that CCL7 can be involved in morphine tolerance by influencing morphine use-related inflammatory corpuscles and G-protein-coupled signaling pathways, and can serve as a potential therapeutic target.
Despite the rigorous bioinformatic analysis in this study, there are still some shortcomings. First, due to the small sample size in the data set, it is still necessary to further expand the sample size to obtain more accurate results. Second, this article only analyzed the bioinformatic data, but did not carry out experimental verification. A large number of clinical samples and animal experiments should be used for comprehensive verification, so as to better understand the molecular mechanism of morphine tolerance.
In conclusion, bioinformatic technology could be a useful tool to explore the mechanism of the occurrence and development of morphine tolerance. In addition, LncRNAs and DEGs were differentially expressed between the morphine tolerance group and nonmorphine tolerance group, which may be involved in the development of morphine tolerance, especially LncRNA DQ266361, uc.167−, and Mmp9, CCL7 genes. The key lncRNA and hub genes in these differential expressions can provide new research targets for the diagnosis and treatment of morphine tolerance.
Footnotes
Acknowledgment
We are thankful to Meng-lei Hao for her statistical assistance and suggestions during the submitting process.
Availability of Data and Materials
The data sets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
Authors' Contributions
Y.Q. and L.-b.M. performed the experiment, and were major contributors in writing the article and submitting the article. Z.H. made substantial contributions to research conception. He also designed the draft of the research process. C.-y.D. and Y.-h.H. had been involved in revising the article critically for important intellectual content. B.-c.Y. and T.-j.Z. analyzed the data. All authors read and approved the final article.
Ethics Approval and Consent to Participate
The data for this research were downloaded from the GEO database, one public website. Also, all institutional and national guidelines for the care and use of participates were followed.
Author Disclosure Statement
The authors declare there are no competing financial interests.