
Abstract
During mammalian embryo development, reprogramming of DNA methylation plays important roles in the erasure of parental epigenetic memory and the establishment of naive pluripotent cells. Multiple enzymes that regulate the processes of methylation and demethylation work together to shape the pattern of genome-scale DNA methylation and guide the process of cell differentiation. Recent availability of methylome information from single-cell whole genome bisulfite sequencing (scBS-seq) provides an opportunity to study DNA methylation dynamics in the whole genome in individual cells, which reveal the heterogeneous methylation distributions of enhancers in embryo stem cells. In this study, we developed a computational model of enhancer methylation inheritance to study the dynamics of genome-scale DNA methylation reprogramming during exit from pluripotency. The model enables us to track genome-scale DNA methylation reprogramming at single-cell level during the embryo development process and reproduce the DNA methylation heterogeneity reported by scBS-seq. Model simulations show that DNA methylation heterogeneity is an intrinsic property driven by cell division along the development process, and the collaboration between neighboring enhancers is required for heterogeneous methylation. Our study suggests that the mechanism of genome-scale oscillation might not be necessary for the DNA methylation heterogeneity during exit from pluripotency.
1. Introduction
In mammalian development, reprogramming of DNA methylation (5-methylcytosine) patterns plays a crucial role in defining cell fate. Upon fertilization, DNA methylation marks represent an epigenetic barrier that restricts mammalian development and, hence, need to be restored and subsequently rebuilt with the commitment to particular cell fates (Seisenberger et al., 2013). The segregation of cell lineages gives rise to different somatic tissues associated with tissue-specific DNA methylation patterns (Styblo et al., 2000; Hon et al., 2013; Greenberg and Bourchis, 2019). The genome-wide DNA methylation reprogramming events coincide with the changes in concentrations of DNA methyltransferases (DNMTs) and the enzymes that initiate the removal of DNA methylation (ten-eleven-translocation family proteins, TETs) (Seisenberger et al., 2013; Greenberg and Bourchis, 2019). The recent maturation of single-cell sequencing technologies has enabled us to observe a variety of sequencing information at individual cell level, such as the genome, transcriptome, and epigenome (Stuart and Satija, 2019). It becomes a challenge issue in computational biology to develop single-cell based computational model that can help us to better understand the process of DNA methylation pattern formation, as well as cell fate decision during early embryo development.
The availability of methylome information from single-cell whole genome bisulfite sequencing (scBS-seq) provides an opportunity to study DNA methylation patterns in the whole genome in individual cells (Smallwood et al., 2014; Farlik et al., 2015; Rulands et al., 2018). A recent study applied scBS-seq to embryo stem cells (ESCs) cultured under naive [two chemical inhibitors (“2i”) of MEK1/2 and GSK3
In Rulands et al. (2018), it was proposed that the DNA methylation heterogeneity is associated with coherent, genome-scale oscillations in DNA methylation, and the amplitude is dependent on the CpG density. Moreover, a mathematical model of delay differential equation with autocatalytic de novo methylation was proposed to show that global oscillations may emerge from the biochemistry of methylation turnover due to Hopf bifurcation with increasing values of the time delay, and a Kuramoto model was applied to describe the global heterogeneous coupling of CpGs through DNMT3a/b binding. Nevertheless, genome-scale oscillations in DNA methylation is a very strong assumption, which may imply global oscillations in transcriptions of most genes, and is not supported by the experimental data of DNA methylation dynamics during transition from naive to primed pluripotency in vitro (Fig. 1F). Hence, we asked how the transitions of DNA methylation heterogeneity should be explained through a simple mechanism?
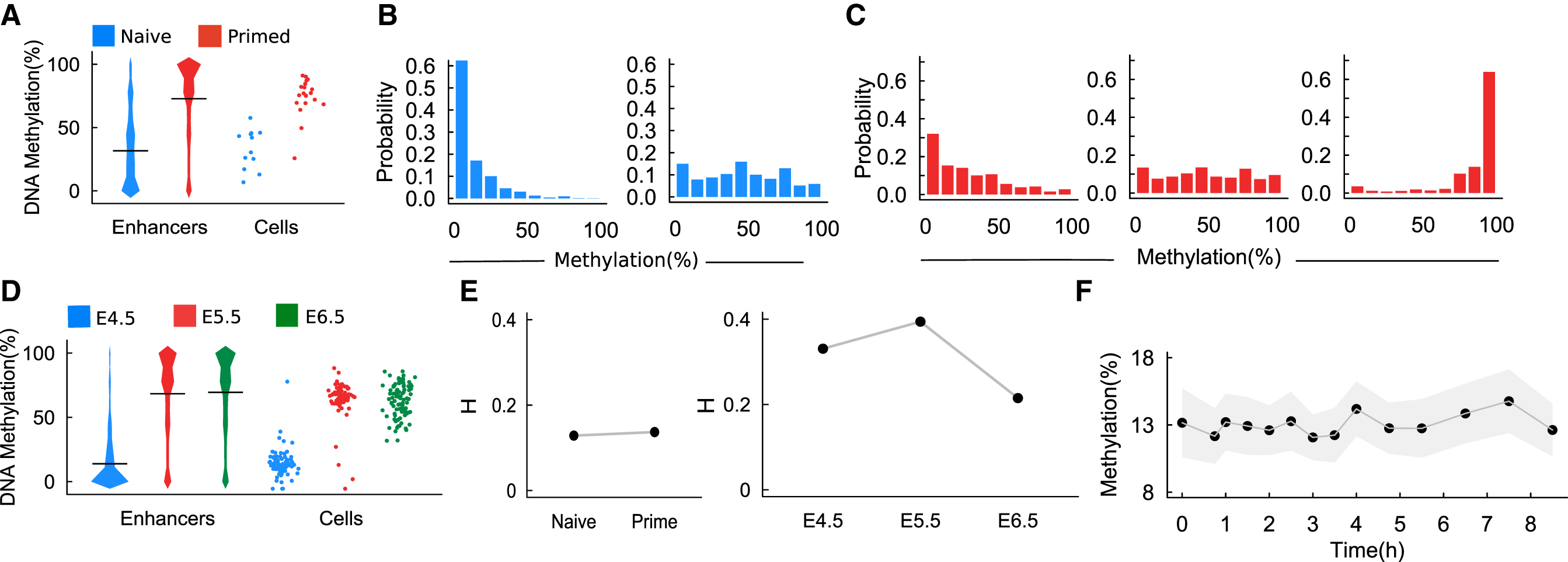
DNA methylation transition in embryo cells.
DNA methylation and chromatin dynamics have been modeled quantitatively in various genomic contexts of biological significance (Sneppen and Dodd, 2011; Haerter et al., 2013; Berry et al., 2017; Huang and Lei, 2017; Song et al., 2017). The collaboration between neighboring CpGs was highlighted in recent studies (Haerter et al., 2013; Song et al., 2017), which play essential roles in the formation of global patterns and the genome-scale transitions of DNA methylation. The collaboration may directly come from the binding of DNMT3a/b to neighboring CpGs (Rulands et al., 2018) or indirectly through the interaction with methylations in the histones H3K9 (Lehnertz et al., 2003) and H3K36 (Weinberg et al., 2019). In addition to the de novo methylation and demethylation, dilution and maintenance of methylated marks during DNA replication may also contribute to the cell-to-cell variance of DNA methylation.
In this study, motivated by the scBS-seq data of heterogeneous methylation at genomic annotations associated with active enhancer elements, we developed a model of DNA methylation, considering the stochastic dynamic of methylation levels of enhancers over cell divisions and the collaboration between neighboring enhancers, to investigate the transition of DNA methylation from naive to primed ESCs. The model focuses on the random inheritance of DNA methylations during cell cycling and can automatically reproduce the DNA methylation heterogeneity on enhancers during embryonic development. Our results suggest that the mechanism of genome-scale oscillation proposed by Rulands et al. (2018) may not be required for the observed heterogeneity during exit from pluripotency.
2. Results
2.1. Transition of DNA methylation patterns from naive to primed ESCs
We analyzed the scBS-seq data separately for ESCs cultured under naive (“2i”) and primed (“serum”) conditions (Rulands et al., 2018). Similar to the analysis in Rulands et al. (2018), taking published H3K4me1 chromatin immunoprecipitation sequencing (ChIP-seq) data from primed ESCs (Creyghton et al., 2010) as a definition of enhancer elements, the methylation levels of enhancers in primed ESCs increase comparing with naive ESCs (Fig. 1A). In this study, the methylation level of an enhancer is defined as the average level of all CpG sites contained in the enhancer. For each CpG site, we assigned a value 0 for unmethylated, 0.5 for half-methylated, and 1 for full methylated; hence, the methylation level of an enhancer takes value from the interval [0, 1] (or from 0% to 100% methylated). Moreover, we calculated the distribution of methylation levels of all enhancers in individual cells. Primed ESCs showed higher cell-to-cell variability at the distribution patterns than the naive ESCs (Fig. 1B, C). We also analyzed the parallel scM&T sequencing of in vivo epiblast cells at E4.5, E5.5, and E6.5 (Rulands et al., 2018), which showed an increase in the methylation level in enhancers from E4.5 to E5.5 (Fig. 1D). We note that there are a few cells that show low methylation levels at E5.5; however, all cells have high methylation at E6.5, which suggests a transition dynamics of methylation levels (Fig. 1D).
To quantify the heterogeneity of DNA methylations among different cells, we proposed a definition of heterogeneity index based on the methylation levels of enhancers in each cell. Assuming that there are n cells, and
where
We calculated the heterogeneity index based on the above data from naive and primed ESCs and the cells at E4.5, E5.5, and E6.5 mice embryo. There are no significant changes in the heterogeneity of naive ESCs in comparing with the primed ESCs (Fig. 1E). For the mice embryo cells, the heterogeneity index increases from E4.5 to E5.5 and decreases from E6.6 to E6.5 (Fig. 1E).
To validate the genome-scale DNA methylation oscillations in Rulands et al. (2018), we analyzed the same data from an in vitro “2i release” model in which cells were transferred from naive 2i to primed serum culture conditions. The average methylation levels of enhancers from the first chromosome (ch1) were calculated to obtain the dynamics from 0 to 8 hours after 2i release. The results do not show significant oscillations in the methylation level (Fig. 1F). These findings suggest that the assumption of genome-scale DNA methylation oscillations might not be necessary to explain the transition of methylation and heterogeneity from naive to primed ESCs.
2.2. Stochastic dynamics of enhancer methylation levels
The dynamics of DNA methylation/demethylation have been modeled quantitatively with exquisite details at biochemistry of single CpG sites (Sontag et al., 2006; Haerter et al., 2013; Song et al., 2017; Lei et al., 2018). The methylation state of a single CpG site is often random due to the stochastic biochemical reactions. Nevertheless, the average methylation level of CpG sites associated with a genomic segment is more predictable. During embryo development, the most significant changes in DNA methylation occur during DNA replication when the 5-methylcytosine marks are diluted to two daughter strains and are restored through enzymes DNMT1 and nuclear protein 95 (NP95 or UHRF1). Correlating global DNA methylation with replication timing repli-seq data has shown that late-replicating regions did not have lower DNA methylation than early-replicating regions (Rulands et al., 2018). Thus, while we omit the detailed dynamics between DNA replications, we can represent the methylation level of an enhancer by the average methylation level at late-replication stage of each cell cycle.
2.2.1. Formulation
To consider the dynamics of enhancer methylation levels, assuming that there are N enhancers in a chromatin and letting
over cell cycles t. During cell cycling, the methylation states update as a consequence of the regulations through enzymes DNMT1, DNMT3a/b, and TETs, which lead to the following iteration
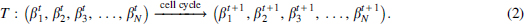
Hence, while we omit the biochemistry details, the stochastic dynamics of enhancer methylation levels can be formulated as an iteration
for each cell cycle. In this study,
We note that the iteration Equation (3) is usually a random map. Given the state of mother cells, the methylation state of the daughter cell is a random valuable whose probability density is dependent on the state of the mother cell. Hence, to formulate the iteration map, we need to write down the conditional probability density function
the probability of
The probability density
where
Now, to define the dependences
and the variance
than
1
Hence, we only need to identify the functions
This simple assumption means the inverse proportion of the variance of enhancer methylation levels with the number of CpGs.
To define the function
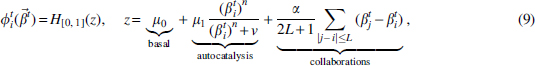
where
Here
The third term in Equation (9) shows the collaboration between neighboring enhancers. Here, we assumed the coherent collective behaviors of DNA methylation/demethylation when the average coupling through enzyme binding is sufficiently strong so that the nearby enhancers tend to the same trends of either methylation or demethylation. The similar mechanism was introduced previously to reproduce the long-distance correlation of DNA methylation between CpGs (Song et al., 2017). Here the collaboration effect is limited by cooperative range L and the coefficient
2.2.2. Numerical scheme and parameters
To model the methylation dynamics following developmental process with the above iteration Equation (2), we first initialized the methylation level of N enhancers
In model simulations, we took
in accordance to the statistics from mouse genome (Fig. 2A). In simulations, we can refer the distribution of methylation levels from a naive cell as the initial condition (Fig. 2B).

Number of CpG sites and the initial methylation level.
2.3. Transition of DNA methylation heterogeneity during the development process
To verify the proposed model, we varied the parameter v (v = 0.1, 0.04, 0.01) to mimic different conditions. For each value v, we initialized a cell with an initial state of a naive cell and ran the model simulation for 15 cell cycles to mimic the transition from naive to primed condition. The simulated distribution of enhancer methylation levels at each cell cycle was calculated and is shown by Figure 3A–C. The enhancer methylation distributions depend on the parameter v: when
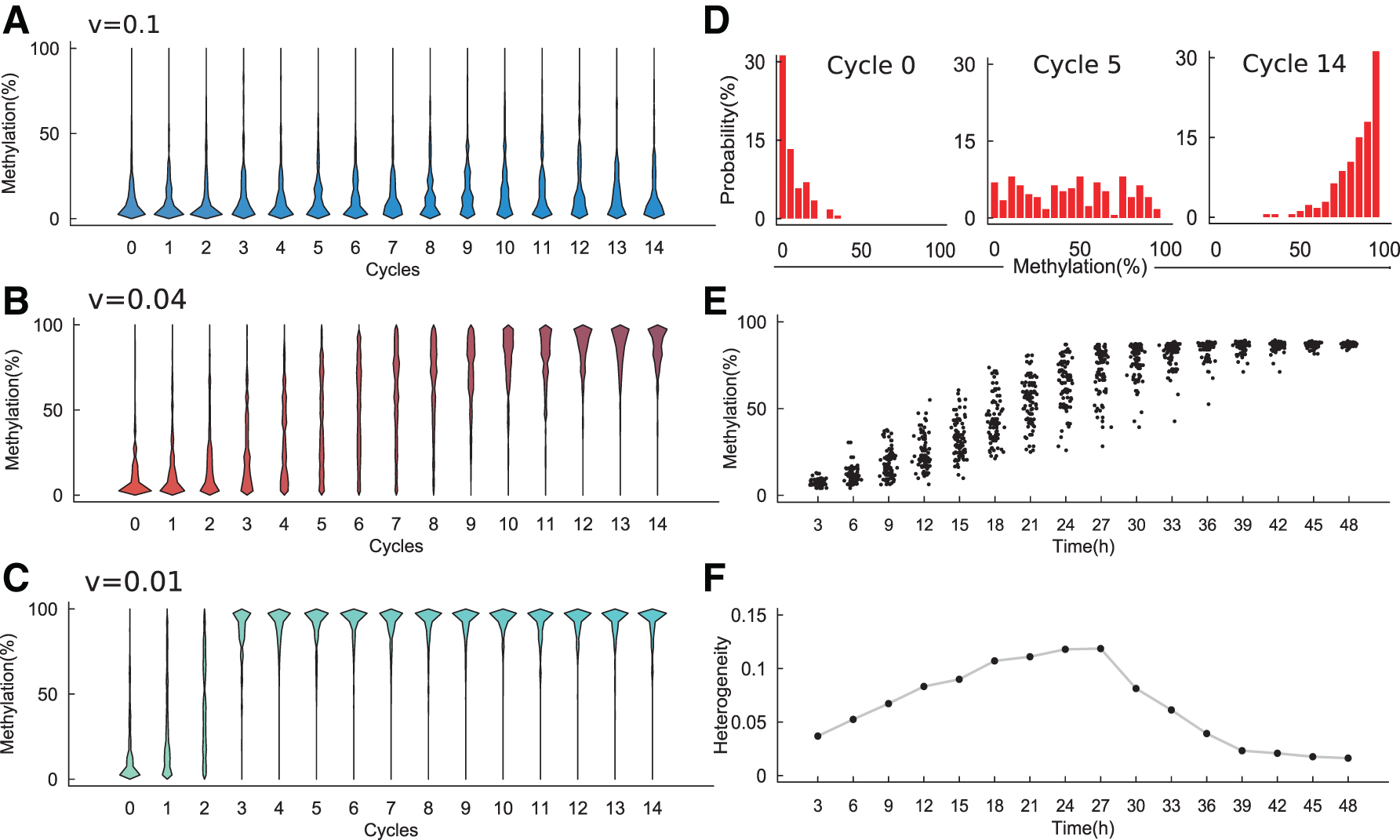
DNA methylation heterogeneity from model simulation.
To further examine the transition dynamics of methylation heterogeneity in ESCs, we set
2.4. Collaboration and methylation heterogeneity
The proposed model includes collaboration between neighboring enhancers so that there are coherent collective behaviors during enzyme binding. To investigate the effects of neighboring collaboration, we varied the parameters

Collaboration and methylation heterogeneity.
To further examine how collaborations may affect the DNA methylation dynamics, we varied the parameters
3. Discussion
Reprogramming of DNA methylation plays important roles in mammalian early embryo development. ESCs show heterogeneous methylation distributions under primed conditions. To understand the mechanism of methylation heterogeneity, previous studies suggest a mechanism of genome-scale oscillations in DNA methylation. Nevertheless, experiment data did not support the assumption of genome-scale oscillations. In this study, we proposed a computational model for the stochastic transitions of enhancer methylations during cell cycling. The model combines random distribution of methylation marks in DNA replication, autocatalysis of DNA methylation due to the binding of DNMT3a/b, and the collaboration between neighboring enhancers during the reconstruction of methylation marks. The proposed model can nicely explain the transition of methylation level and heterogeneous methylation distributions. Model simulations showed that the proper values of the autocatalysis are important for the heterogeneity between different cells, and increasing the collaboration between neighboring enhancers can promote the heterogeneity. Our model suggests that methylation heterogeneity is a natural consequence of stochastic transition of DNA methylation between cell cycles and the collaboration between CpGs; however, the assumption of genome-scale oscillations might not be necessary for the observed heterogeneous methylation distributions.
The proposed model mainly considers the dynamics of enhancer methylation levels in a cell and omits the biochemical reactions involved in methylation or demethylation, which may be regulated by various enzymes. In the model, we assumed a beta distribution that connects the methylation level in daughter cells with those of the mother cell. The beta distribution can take different forms based on the shape parameters that are defined by the state of mother cells. Thus, the proposed model framework mainly focuses at the general effect of methylation state transition between cell cycles, while omitting the details of biochemical reactions. In contrast, despite the complex biochemical reactions, they mainly affect the methylation levels through the based methylation/demethylation processes and the collaboration between CpGs and, hence, may end up to the function
Footnotes
Author Disclosure Statement
The authors declare they have no conflicting financial interests.
Funding Information
This work is supported by the National Natural Science Foundation of China (11831015 and 11872084).