
Abstract
The root architecture of wild tomato, Solanum pimpinellifolium, can be viewed as a network connecting the main root to various lateral roots. Several constraints have been proposed on the structure of such biological networks, including minimizing the total amount of wire necessary for constructing the root architecture (wiring cost), and minimizing the distances (and by extension, resource transport time) between the base of the main root and the lateral roots (conduction delay). For a given set of lateral root tip locations, these two objectives compete with each other—optimizing one results in poorer performance on the other—raising the question how well S. pimpinellifolium root architectures balance this network design trade-off in a distributed manner. In this study, we describe how well S. pimpinellifolium roots resolve this trade-off using the theory of Pareto optimality. We describe a mathematical model for characterizing the network structure and design trade-offs governing the structure of S. pimpinellifolium root architecture. We demonstrate that S. pimpinellifolium arbors construct architectures that are more optimal than would be expected by chance. Finally, we use this framework to quantify structural differences between arbors grown in the presence of salt stress, classify arbors into four distinct architectural ideotypes, and test for heritability of variation in root architecture structure.
1. Introduction
Root architecture in plants is driven by the development of lateral roots that branch off of the main root (Nibau et al., 2008). Our goal is to derive a model that explains the principles governing branching patterns in Solanum pimpinellifolium root architecture. Root architecture design that affords efficient resource transport is crucial to the overall productivity of the plant (Lynch, 1995); however, it can be advantageous to minimize the amount of material required to construct arbors due to environmental constraints such as resource limitations (Niu et al., 2013), soil compaction (Chen et al., 2014), growth environment orientation (Chiatante et al., 2003), and salt stress (Julkowska et al., 2017). Prior study models plant arbors as a weighted graph, and uses the theory of Pareto optimality to resolve the trade-off between minimizing resource transport delay and minimizing material cost (Conn et al., 2017a,b, 2019). We extend prior study by applying a model developed for Arabidopsis to quantify design trade-offs in S. pimpinellifolium.
Root architecture is highly plastic, showing adaptability to the environments and stress conditions such as salt stress (Julkowska et al., 2014). These salt-induced changes in root architecture were previously used to identify genetic components contributing to overall stress-resilience (Julkowska et al., 2017). Through screening S. pimpinellifolium, which is stress-resilient relative to cultivated tomato, we aim to identify novel strategies that contribute to S. pimpinellifolium resilience. Through modeling and analyzing network topology, we attempt to gain further insight into network efficiency and adaptability to stress conditions.
In addition to observing high stress-resilience, we find that S. pimpinellifolium arbors can be visually classified into four distinct architectural ideotypes: “Christmas Tree,” “Telephone Pole,” “Droopy Telephone Pole,” and “Broomstick” (Fig. 1A). However, a preliminary analysis revealed that this classification scheme is not associated with root structure variation according to traditional physical features (Fig. 1B–D). Our goal was to study alternate methods for quantifying network topology, to test whether our qualitative classification scheme captures meaningful variation in root architecture.

Solanum pimpinellifolium root ideotypes across the diversity panel.
Our article is organized as follows. First, we define a graph-theoretic model S. pimpinellifolium arbors that formalizes the objectives of wiring cost and conduction delay. We demonstrate how to resolve the trade-off between these two objectives using the theory of Pareto optimality. We use this framework to investigate how well S. pimpinellifolium arbors optimize wiring cost and conduction delay, and infer whether these objectives are significant drivers of root architecture topology. We then investigate variation in how arbors prioritize objectives. We test whether arbors prioritize objectives in the presence of a salt stressor; we propose a scheme for classifying arbors into four architectural ideotypes, and use network design trade-offs to validate this classification scheme. Finally, we explore whether genetic variation is associated with arbors prioritizing objectives differently.
1.1. Related study
In modeling and analyzing the network topology of S. pimpinellifolium, we relate our study to the Euclidean Steiner tree optimization problem (Agrawal et al., 1995; Arora, 1996; Karpinski and Zelikovsky, 1997). We study the Steiner tree problem with certain constraints on the solution space; in addition, our Steiner trees must balance two cost functions instead of one.
Our study is also closely related to the problem of finding a Light Approximate Shortest-Path Tree (Khuller et al., 1995; Nguyen and Xu, 2007). Given a rooted graph, the goal is to find a spanning tree whose total weight is not too high while maintaining short paths from the root to each vertex. Our study attempts to solve a similar problem but allows for branch points that are not part of the original network.
2. Methods
Hereunder we describe how we use graph theory, as well as the principle of Pareto optimality, to model S. pimpinellifolium root architecture and quantify design trade-offs.
2.1. A graph-theoretic framework for studying S. pimpinellifolium architecture
We model the S. pimpinellifolium root architecture as a weighted connected acyclic graph
Each of the vertices
The main root vertices from a path graph—that is, they can be ordered
Each lateral root vertex has degree 1, and is directly connected to one of the main root vertices. Lateral root vertices do not connect to another lateral root segment.
For each edge
Let the wiring cost be the sum of the edge lengths of all edges in the graph:

Let the conduction delay be the sum of graph distances from the main root base to each lateral root:

2.2. Pareto-optimal root architecture problem
In general, it is impossible to design a root architecture that simultaneously minimizes wiring cost and conduction delay. Furthermore, the trade-off between these two objectives can become arbitrarily large (Fig. 2). To resolve this trade-off, we turn to the principle of Pareto optimality (Da Cunha and Polak, 1967) to define the problem of constructing an optimal arbor.
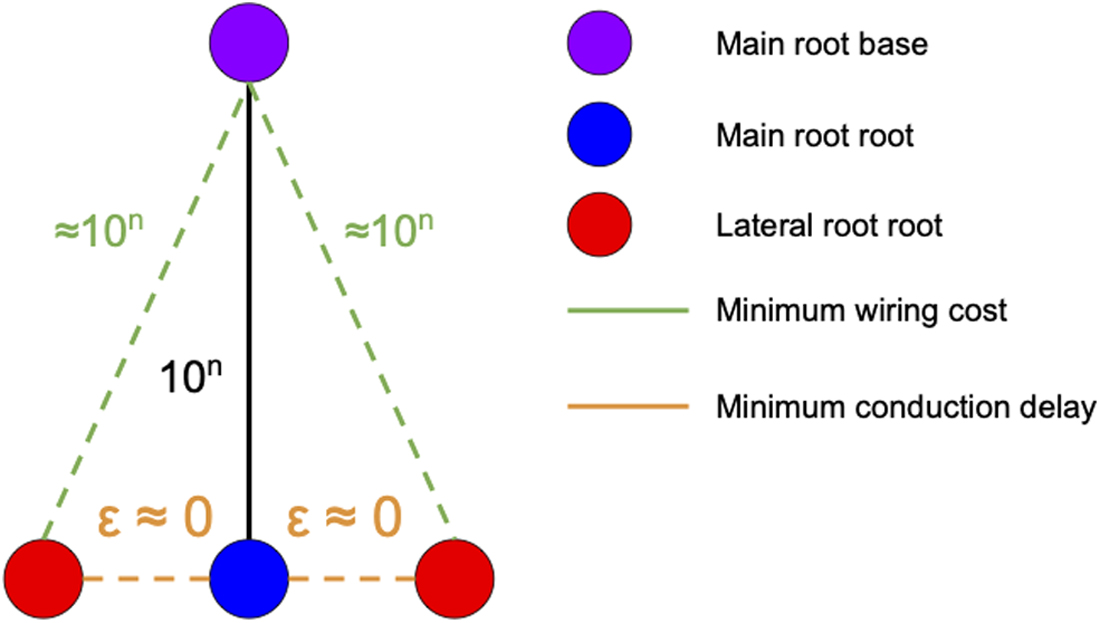
Trade-off between wiring cost and conduction delay. In this example, there are two lateral roots that seek to connect to the main root. The main root base is
As input, we are given the following (Fig. 3A):

Pareto front of optimal solutions.
A set of main root points
A set of lateral root tips
A value
We then construct an architecture graph A by connecting each lateral root to the main root (Fig. 3A). Each lateral root may connect to one of the initial main root points, or it may connect to a midpoint on one of the edges
The parameter
2.3. An algorithm for generating optimal root architectures
Because lateral roots cannot connect to each other, we may simply optimize each lateral root independently. Because the main root is fixed, and because lateral roots have degree 1, optimizing a lateral root reduces to finding the best straight line between that lateral root and a point along one of the main root line segments. This can be done analytically using calculus and linear algebra.
For this section, we will put variables in
A line segment is defined by its two endpoints
Consider a lateral root located at the point
If we connect
The increase in conduction delay is
The total increase in cost is
We then seek to find the value of
We then rearrange the equation to isolate t:

To make the equation easier to read, we define the following variables:

This gives us
Once again, we will define new variable substitutions to make the equation easier to read:
Then we have
To solve for t, we simply use the quadratic formula:
Note that this formula is only valid for
Up to this point, we have assumed that our line segment
Finally, consider a lateral root at point  . We compute the optimal midpoint for each line segment
. We compute the optimal midpoint for each line segment
We repeat this process independently for each lateral root. In this way, we may analytically compute the optimal arbor for a given value of
2.4. Quantifying network design trade-offs in observed architectures
Given an observed architecture A, we may use the aforementioned algorithm to compute the set of optimal root architectures: Pareto front distance: The L-2 distance between costs of arbor A and the costs of the closest optimal tree:
This measures how close the original arbor came to managing an optimal trade-off between wiring cost and conduction delay. Pareto front location: The value of
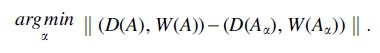
The closer this value to 1, the more strongly the arbor prioritized minimizing wiring cost at the expense of conduction delay.
Under this framework, we tested four hypotheses: (1) S. pimpinellifolium roots are closer to being Pareto optimal than would be expected by chance; (2) S. pimpinellifolium roots respond to salt stress by modifying network design trade-offs; (3) variation in ideotype classification is associated with variation in Pareto front location; (4) Pareto front location is a heritable trait that may be driven (in part) by genetic factors.
2.5. Comparing observed arbors against a null model
Using the aforementioned mathematical framework, we sought to confirm whether S. pimpinellifolium root architecture structure is dictated in part by the principles of wiring cost and conduction delay (as opposed to some other optimization criteria). We did so by comparing S. pimpinellifolium root architecture with the arbors that were Pareto optimal for these objectives. If wiring cost and conduction delay are drivers of S. pimpinellifolium root architecture, then observed arbors should be more similar to the optimal arbors than would be expected by chance.
To evaluate how well S. pimpinellifolium would be expected to optimize wiring cost and conduction delay by chance, we also compared observed arbors with arbors that were randomly generated on the same set of points. This allowed us to establish a baseline for how similar S. pimpinellifolium arbors would be to optimal arbors even if they were explicitly not optimizing wiring cost or conduction delay. To generate a random arbor, we start with the same main root segments, and same lateral root tip locations, as the original observed arbor. Each lateral root tip is then connected to a point chosen at uniform random from the set of line segments comprising the main root.
3. Empirical Data
The natural diversity panel 260 accessions of S. pimpinellifolium was screened for salt-stress-induced changes in root system architecture. The seeds were surface sterilized using 50% bleach for 10 minutes and washed 4–6 times using sterile MiliQ water. The seeds were stratified at 4°C for 24 hours to improve germination. The seeds germinated on square petri dishes (12 × 12 cm) on 1/4 Murashi–Skoog media (1.1 g MS, 5 g sucrose, 1 g M.E.S. buffer, pH 5.8 KOH, and 10 g Dashin agar). Four days after germination, the seedlings were transferred to square petri dishes containing 1/4 Murashi–Skoog media for Control treatment or 1/4 MS media supplemented with 100 mM NaCl for Salt treatment. The plates were scanned every day for 7 consecutive days.
The root system architecture was quantified from the images recorded 0, 1, 2, 3, and 4 days after transfer to Control and Salt plates using SmartRoot software (Lobet et al., 2011). The points (x–y coordinated) representing the observed root system architecture are obtained using SmartRoot. The nodes are added by the software automatically every time a slight change in the root trajectory is observed (Fig. 4). Addition of the nodes in between the observed nodes on the main root trajectory would not affect either the overall shape or size of the root architecture or the number of lateral roots for our model. The global root data were compiled and analyzed in R for lengths of main and lateral roots as well as growth rates of main root and lateral roots. The root node data were used for calculating the Pareto optimality.

Graphic representation of the collected root system architecture coordinates. The position of the nodes corresponding to the locations of the main root (yellow) and lateral roots (green) were collected by using SmartRoot software. The position of each node is added whenever the slight change in the root trajectory is observed, to ensure the most precise tracking.
Plants were manually and qualitatively classified into one of four representative ideotype accessions (Fig. 1A). The root systems where lateral roots were decreasing in length throughout the main root axis were classified as “Christmas Tree.” The root systems where the lateral root was constant throughout the main root axis and where lateral roots were growing at ∼90° angle from the main root axis were classified as “Telephone Pole.” The root systems similar to “Telephone Pole,” but with lateral roots growing under ∼45° angle from the main root axis were classified as “Droopy Telephone Pole.” The root systems where majority of the lateral roots originated at the root base were classified into “Broomstick” ideotype.
4. Results
4.1. S. pimpinellifolium roots are Pareto optimal
For each arbor, we computed the Pareto front of optimal architectures that the arbor could have constructed, and measured each arbor's distance to its respective Pareto front. To test whether arbors are closer to being Pareto optimal than would be expected by chance, we compared each arbor against 20 randomly generated arbors (Fig. 5A). We find that S. pimpinellifolium arbors are more optimal than random trees 98.61% of the time, which is significantly >50% (binomial test,

Analysis of Pareto front distance and location.
4.2. Salt stressor affects network design trade-offs
Arbors grown in control conditions tended to prioritize optimizing wiring cost more than arbors grown in conditions with a salt stressor (Fig. 5B). The average value of
4.3. Classification of ideotypes is associated with variation in network design trade-offs
Finally, we find a strong association between an arbor's architectural ideotype and the degree to which an arbor prioritizes trade-offs between wiring cost and conduction delay (Fig. 5C). There are significant differences in Pareto front location between different ideotype groups (Kruskal–Wallis one-way analysis of variance;
4.4. Network design trade-offs are heritable
The variation for Pareto front location was lower within a single genotype than the variation observed across individual genotypes. We calculated broad-sense heritability
5. Discussion
Similar to many natural and engineered systems, S. pimpinellifolium root architecture must balance trade-offs between multiple competing objectives. Generally, there is not a single architecture that is optimal for all possible objectives. Instead, the best the arbor can do is converge to an architecture that is Pareto optimal—meaning it is impossible to improve one objective without sacrificing performance in the other objectives. Our study proposes two objectives—minimizing wiring cost and minimizing conduction delay—that S. pimpinellifolium roots seek to optimize in a Pareto optimal manner. Using graph theory and the principle of Pareto optimality, we define a framework for quantifying the optimality of a given S. pimpinellifolium root architecture.
We apply this model to S. pimpinellifolium imaging data and find that S. pimpinellifolium arbors are significantly closer to being Pareto optimal for these objectives than would be expected by chance. We use Pareto front location to show that S. pimpinellifolium arbors prioritize objectives differently in the presence of a salt stressor. We find that unlike traditional network design metrics, Pareto front location quantifies S. pimpinellifolium architecture in a way that is strongly associated with the architectural diversity we observe qualitatively. Finally, we find that Pareto front location shows a strong association with genetic variation. Overall, our study provides a framework for quantifying S. pimpinellifolium based on how and how well they manage network design trade-offs. More broadly, our study provides a blueprint for quantifying how natural and engineered systems manage trade-offs between several competing objectives.
6. Future Directions
Pareto front location is associated with our classification of S. pimpinellifolium arbors into architectural ideotypes. Future study involves using clustering analysis of Pareto front location to conclusively validate our chosen classification scheme (or identify the correct number of clusters). If we find evidence for the existence of our four identified ideotypes, future study will seek to find the functional basis behind each of these four structures.
In addition to formulating an exact algorithm for generating Pareto-optimal arbors, we seek to study the development process of S. pimpinellifolium. We will take advantage of data tracking the growth of individual arbors over time to measure how arbors prioritization of objectives changes over time, and eventually define a model for the distributed growth process used to generate Pareto optimal architectures.
Future study includes Genome Wide Association Study, to identify underlying genetic components of Pareto front location and distance in S. pimpinellifolium roots. Through reverse genetic approaches we aim to further study genetic components of Pareto front location, their role in root architecture development and further developing better understanding on how S. pimpinellifolium plants prioritize objectives during nonstress and stress conditions.
Finally, we will expand our model to study the trade-off between not only wiring and transport efficiency, but also gravitropism and growth media exploration. Certain arbors may have a higher tendency to grow toward or away from gravitational forces in the growth medium. Furthermore, certain arbors may exhibit higher nutrient uptake at the main root, thus leading lateral roots to explore farther horizontally to avoid competing with the main root. We will expand our mathematical model to include these factors as extra constraints on how lateral roots may connect to the main root (while still optimizing wiring cost and conduction delay.
Footnotes
Authors’ Contributions
Data curation, formal analysis, methodology, software, validation, visualization, and writing (original draft preparation) by A.C. Conceptualization, data curation, formal analysis, investigation, methodology, visualization, and writing (original draft preparation) by M.M.J.
Acknowledgments
We thank the organizing committee for the 2021 Biological Distributed Algorithms (BDA) conference, as well as the reviewers for or BDA submission. We thank Guillaume Lobet for help with understanding the output from the plant imaging software. We thank Graham Zug, who is currently working with Dr. Chandrasekhar on mathematical optimization algorithms for constructing Pareto optimal plant arbors. Finally, we thank bioRxiv for hosting a preprint of our study (DOI: 10.1101/2021.08.12.456185).
Data Availability
Author Disclosure Statement
The authors declare they have no competing financial interests.
Funding Information
Dr. Julkowska's study was financed by the King Abdullah University for Science and Technology and the Boyce Thompson Institute.