
Abstract
In recent years, there has been increasing demand for red tilapia, which are commercial strains of hybrids of different tilapiine species or red variants of highly inbred Nile tilapia. However, red tilapia phenotypes are genetically unstable and affected by environmental factors, resulting in nonuniform coloration with black or dark-red color blotches that reduce their market value. Solute carrier family 45 member 2 (SLC45A2) is a membrane transporter that mediates melanin biosynthesis and is evolutionarily conserved from fish to humans. In the present study, we describe the generation of a stable and heritable red tilapia phenotype by inducing loss-of-function mutations in the slc45a2 gene. For this purpose, we identified the slc45a2 gene in Nile tilapia and designed highly specific guide RNAs (gRNA) for its genomic sequence. Multiplex microinjection of slc45a2-specific ribonucleoproteins to Nile tilapia zygotes induced up to 97–99% albinism, including loss of melanin in the eye. Next-generation sequencing of the injected zygotes demonstrated that all injected fish carried mutant alleles with variable mutagenesis efficiencies. Sanger sequencing of the genomic target region in the slc45a2 gene from fin clips, sperm, and F1 offspring of a highly mutant male identified various genomic indels and germline transmission of the sperm-identified indels. Overall, this work demonstrates the generation of somatic and germline slc45a2 mutant alleles, which leads to complete albinism in Nile tilapia.
Introduction
Tilapias are the second most aquacultured fish group in the world, with a global production of roughly 5.5 million tons per year, mostly of Nile tilapia (Oreochromis niloticus). 1 In recent years, red tilapias, which are usually different hybrid strains of Nile tilapia crossed with Mozambique, blue, and Zanzibar tilapias (O. mossambicus, O. aureus, O. hornorum), have been gaining worldwide popularity, and their prices have increased accordingly.2,3 The market price of red tilapia in the Philippines is double that of gray-colored wild-type (WT) tilapia, and for similar reasons, 85% of the tilapias grown in Malaysia are red tilapia strains.4,5 It has been suggested that the red coloration is due to faulty melanophore development. However, the resulting phenotype is unstable, as black or dark-red spots reduce the fish market value.2,6 An alternative approach of phenotypic selection for red coloration has resulted in a significant loss of genetic variability due to a founder effect. 7 Moreover, current red tilapia strains are more sensitive to cold stress induced by overwintering 2 and display reduced growth compared to other commercially used Nile tilapia strains.4,8
Solute carrier family 45 member 2 (SLC45A2), also known as membrane-associated transporter protein (MATP), absent in melanoma-1 (Aim-1), oculocutaneous albinism type 4 (OCA4), B gene, and as albino (alb), is an evolutionarily conserved key mediator of melanin biosynthesis.9–13 SLC45A2 shares high similarity with sucrose transporter proteins in animals and plants. However, it is likely to affect tyrosinase activity through regulation of melanosomal pH.14,15 It was previously suggested that the red phenotype results from dermis blood irrigation in tilapia with perturbed melanophore development. 6 Nonetheless, red to albino-like tilapia strains display black eye pigmentation that is usually accompanied by a variable rate and pattern of black blotching.2,3,6,16 In contrast, naturally occurring loss-of-function mutations in the slc45a2 gene resulted in complete and heritable albinism in medaka 9 and zebrafish, 15 which was transiently phenocopied in Atlantic salmon (Salmo salar) 17 and marine medaka (Oryzias melastigma). 18 Thus, the currently available strains of red tilapia are less likely to have resulted from a naturally occurring null mutation in the slc45a2 gene.
Several methods allowing reverse genetics in fish have been developed in recent years. These include the zinc-finger nucleases, TALEN, and CRISPR-Cas9 methods. 19 In addition to the high scientific value of these technologies, they also possess great potential for agriculture-related applications, such as perturbation of genes with a negative agricultural-commercial value. 20 Among these methods, CRISPR-Cas9 is considered the most efficient due to its relative technical simplicity and because of its high mutagenesis rates in vivo.19,21 Furthermore, it was recently demonstrated that the high effectiveness of the CRISPR-Cas9 system can allow the characterization of phenotypes resulting from loss-of-function mutations already in the injected fish (F0). 21 Indeed, microinjection of slc45a2- or tyr-specific gRNA with Cas9 mRNA resulted in somatic deletions of these genes and reduced melanin formation in the skin of salmon and lamprey larvae.17,22 While zebrafish embryos are transparent and have thin chorions, making them highly amenable to genomic manipulation, and salmon zygotes develop to the two-cell stage at 24 h post fertilization, allowing time for technical microinjection,23,24 Nile tilapia zygotes are opaque, have a thick chorion, and reach the two-cell stage at around 1.5 h post fertilization, making them less amenable to microinjection.25,26 Nonetheless, application of TALEN and CRISPR-Cas9 methodologies in Nile tilapia was shown to be feasible.27,28
Current data suggest that although red tilapia strains are available, their color phenotypes are not uniform, and the regulating genes remain largely unknown.2,3,16 This emphasizes the need for a tilapia strain with an inheritable uniform red color that is stable throughout life and which is established on a clear genetic background that would allow tractability of its inheritance under various breeding and genetic selection protocols. In the present work, we describe the identification of slc45a2 in Nile tilapia and utilization of CRISPR ribonucleoproteins (RNPs) for induction of somatic and germline indels in the tilapia slc45a2 exon 1. We then demonstrate the use of high-resolution melt (HRM) curve analysis for high-throughput screening of F1 heterozygotes, which identified heterozygotes for the generation of an albino-red tilapia strains carrying a germline slc45a2 null alleles.
Methods
Fish handling
Experiments were approved by the Agricultural Research Organization Committee for Ethics in Using Experimental Animals (approval number 472/17 IL). The Nile tilapia stock originated from Lake Manzala, Egypt, and was brought to Israel in the 1990s. Sexually mature Nile tilapia were maintained in 150 L aquaria in harems consisting of one male and four to six females. The temperature was maintained at 24–26°C, and the photoperiod was set at 14L:10D. Harems were constantly monitored for spawning behavior. Fertilized zygotes were collected for microinjection immediately after natural spawning or by performing in vitro fertilization (IVF), as previously described. 29 Larvae for F1 and F2 analysis were collected 2–3 days after natural fertilization.
Cloning of target sites for gRNAs
Genomic DNA (gDNA) was extracted from fin-clip samples using the HotSHOT method. 30 Genomic target sequence was downloaded from the UCSC genome browser. Amplification of slc45a2-exon1 genomic region (accession no. XM_003451484) was performed using specific primer pairs (Table 1), as previously described. 31 Briefly, polymerase chain reaction (PCR) products were amplified using DreamTaq Green PCR Master Mix (Thermo Fisher Scientific, Vilnius, Lithuania) and analyzed on 1% agarose (LifeGene, Modi'in, Israel) containing Redsafe™ stain (Intron Biotechnology, Seongnam, Korea) in 1 × Tris-acetate acid-EDTA buffer (Biological Industries, Kibbutz Beit-Haemek, Israel). PCR products of the predicted amplicon size were extracted from the gel using a GEL/PCR Extraction Kit (Hy Laboratories Ltd., Rehovot, Israel), cloned into a pGEM-T easy vector (Promega, Madison, WI) and sequenced using T7 primer at Hy Laboratories Ltd.
Design of CRISPR target sites and synthesis
Single guide RNA (sgRNA) design and synthesis was performed, as previously described 32 with slight modifications. Briefly, gRNA was designed using CHOPCHOP, 33 transcribed in vitro using MEGAshortscript T7 kit (Life Technologies, Carlsbad, CA), and purified using a miRNeasy kit (Qiagen, Germantown, MD). slc45a2-specific crRNAs and tracrRNA (TRACRRNA05N) were purchased from Sigma–Aldrich Israel Ltd. (Rehovot, Israel) and diluted with 10 mM TRIZMA buffer (Sigma–Aldrich Israel Ltd.). Recombinant Cas9 (rCas9) protein was produced by the Weizmann Institute of Science Protein Purification Unit (Rehovot, Israel) using the pET-28b-Cas9-His 34 (Alex Schier Lab Plasmids, Addgene, Cambridge, MA) as a template. Sequences of gRNA targets are listed in Table 2.
PAM, protospacer adjacent motif.
Microinjection procedure
Microinjection to tilapia zygotes and gRNA/Cas9 heterocomplexing were performed as previously described.26,32 Tilapia zygotes were collected immediately after natural spawning or IVF and chilled on ice to 21°C to prolong microinjection time at the single-cell stage. In each microinjection session, zygotes were injected with a mix containing a single or two to three mixed slc45a2 gRNAs (100 pmol for each crRNA and tracrRNA; 39.72 pmol for sgRNA) mixed with rCas9 (4.4 μg) to a final volume of 9 μL. gRNA/tracrRNA/rCas9 mix was incubated for 5 min at room temperature to allow the generation of RNP heterocomplexes. Microinjection was performed using glass capillaries (1B150F-4 100 mm; WPI, Sarasota, FL) pulled on a Pul-1000 four-step micropipette puller (WPI). Microinjections were carried out using PV 830 Pneumatic Picopump (WPI) calibrated to deliver a volume of ∼3–5 nL. Following microinjection, zygotes were allowed to develop at 27°C under constant agitation. Three sets of injection were performed for gRNA1, one set for sgRNA1 + gRNA2 + gRNA3, and four sets for gRNA2 + gRNA3. Each injection set contained ∼30–50 zygotes with a wide mortality range and mutation rate. Control non-injected zygotes were grown separately under the same conditions.
Analysis of injected embryos and offspring
gDNA was extracted from whole embryos 4 days post fertilization (dpf) or from fin clips and sperm from F0 and F1 fish and was used for amplification of the target region in the genomic slc45a2-exon1. Amplification products were cloned and sequenced as described above. Retrieved sequences were aligned by MUSCLE.
Next-generation amplicon sequencing
The target region of gRNA2 and gRNA3 was amplified from gDNA of 40 injected fish using sp1 and sp2 primers (listed in Table 1). Each amplicon was purified using GEL/PCR Extraction Kit (Hy Laboratories Ltd.). Samples were subsequently used for the generation of sequencing libraries and sequenced according to the Illumina NovaSeq system protocols at Syntezza Bioscience Ltd. (Jerusalem, Israel). Data of 39 successfully sequenced samples were analyzed using NGS Cas-analyzer. 35 The results were subsequently analyzed manually according to length, gRNA location, indels identified, and the reads of each allele. Unmapped sequences were considered insignificant footprint. The frequency of each allele was calculated from total reads.
HRM analysis
gDNA was extracted from fin-clip samples of 330 F1 fish using HotSHOT method. Each reaction consisted of 5 μL Accumelt HRM Supermix (Quanta Biosciences, Gaithersburg, MD), 1 μL DNA (diluted 1:400), 3.4 μL ultra-pure water, and 0.3 μL of 10 μM for forward and reverse primers each (Table 1). The reaction was performed on 96-well plates. Genomic DNA of WT and known mutant alleles were used as control samples and were run in duplicates using a StepOnePlus™ v2.3 Real-Time PCR System (Applied Biosystems, Foster City, CA). Amplification was performed under the following conditions: 95.0°C for 10 min, 40 cycles at 95.0°C for 15 s, 60.0°C for 30 s, and 72°C for 15 s. Melting curves were generated by a single stage of 95.0°C for 15 s, 60.0°C for 1 min, followed by a temperature increase to 95.0°C at a 0.3% ramp rate. The database was further analyzed using the Applied Biosystems High-Resolution Melt Software v3.1 (Thermo Fisher Scientific, Waltham, MA) with an analysis temperature range of 84.1–89.9°C.
Melanin quantification
Between 5 and 10 scales were randomly sampled from the body centre of 20 randomly selected F1 fish (5 WT and 15 heterozygous) and were genotyped by HRM. Additionally, scales from F2 homozygous mutants were sampled to demonstrate a complete melanin loss. Scales were fixed in paraformaldehyde and then washed three times in phosphate-buffered saline. Scales were imaged using a Nikon SMZ25 stereoscope. Lateral-line canal positive scales were omitted due to their structural divergence. For each scale, epithelial area was measured, and a melanophore cell count was performed using ImageJ. Average melanophore density from two to eight scales/fish was calculated as melanophore/mm2.
Results
Identification of the slc45a2 gene in the Nile tilapia genome
To identify the Nile tilapia slc45a2 gene, a BLASTn search was performed against the nucleotide collection database of Nile tilapia (taxid: 8128), using the previously identified zebrafish slc45a2 mRNA sequence (NM_001110377)11,15 as the query input. As expected, this search yielded a single predicted slc45a2 mRNA (XM_003451484), which was localized to LG7 and shared >77% identity at the 5′ of the nucleotide sequence. Phylogenetic analysis of slc45a2 mRNA sequences from various vertebrate species, including mammals, birds, reptiles, amphibia, and fish and using the agnathan sea lamprey as the phylogenetic root, showed that slc45a2 is evolutionarily conserved from fish to human (Fig. 1A). Furthermore, the slc45a2 gene displayed high syntenic conservation not only with zebrafish but also with human and chicken (Fig. 1B). These findings support the hypothesis of a common ancestral slc45a2 gene in vertebrates and suggest functional conservation for SLC45A2 throughout evolution.
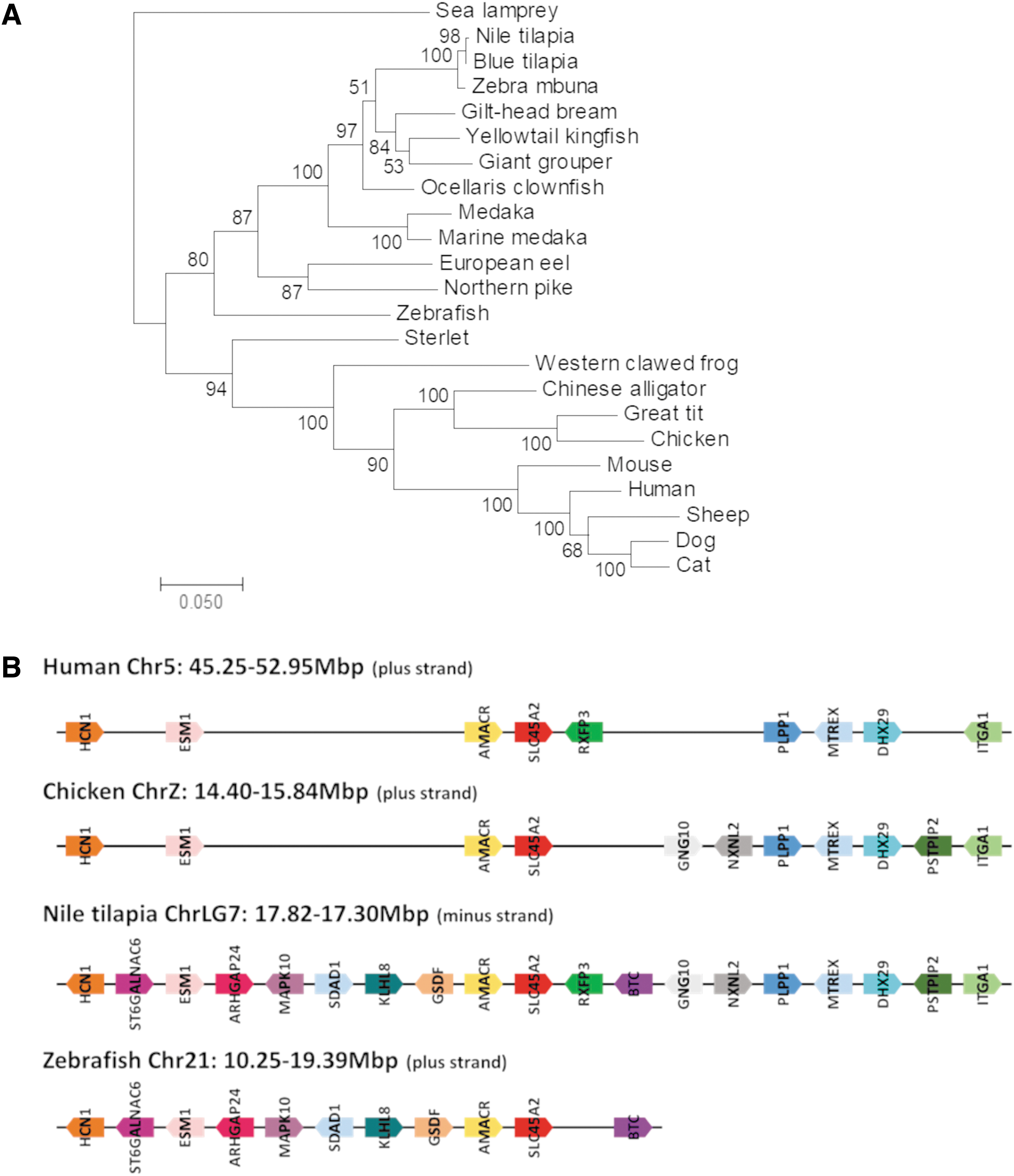
Evolutionary conservation of the Nile tilapia slc45a2 gene. Phylogenetic tree
Design and application of CRISPR-Cas9 to tilapia zygotes
The wide geographical distribution of Nile tilapia results in relatively high genetic diversity.36,37 Aiming to design highly specific gRNAs against slc45a2, we sequenced its exon1 in our in-house brood stock and identified two alleles, each differing in one nucleotide from the public sequence of the Nile tilapia genome (Supplementary Fig. S1). Four specific gRNAs were designed and synthesized, one of which (sgRNA1) was synthesized as sgRNA. Before microinjection, gRNAs were mixed with tracrRNA (trRNA) and recombinant Cas9 (rCas9) protein to allow the generation of RNP heterocomplex.34,38 Nile tilapia zygotes at the single-cell stage were microinjected with slc45a2-RNPs in multiplex and the development of melanin was examined at 4 dpf. At this developmental stage, non-injected tilapia embryos showed a significant amount of melanophores on their body and yolk surface as well as clear melanin formation in the eye (Fig. 2A–C). 25 Strikingly, larvae injected with slc45a2-RNPs exhibited severe to complete loss of melanin in their body and eyes (Fig. 2D–F). Analysis of the target regions using gDNA extracted from slc45a2-RNP-injected larvae demonstrated multiple genomic indels, which varied between embryos (Supplementary Fig. S2). Furthermore, our data showed that both sgRNA and two-component combinations (crisprRNA with tracrRNA) could be used to induce genomic indels by microinjection to zygotes.
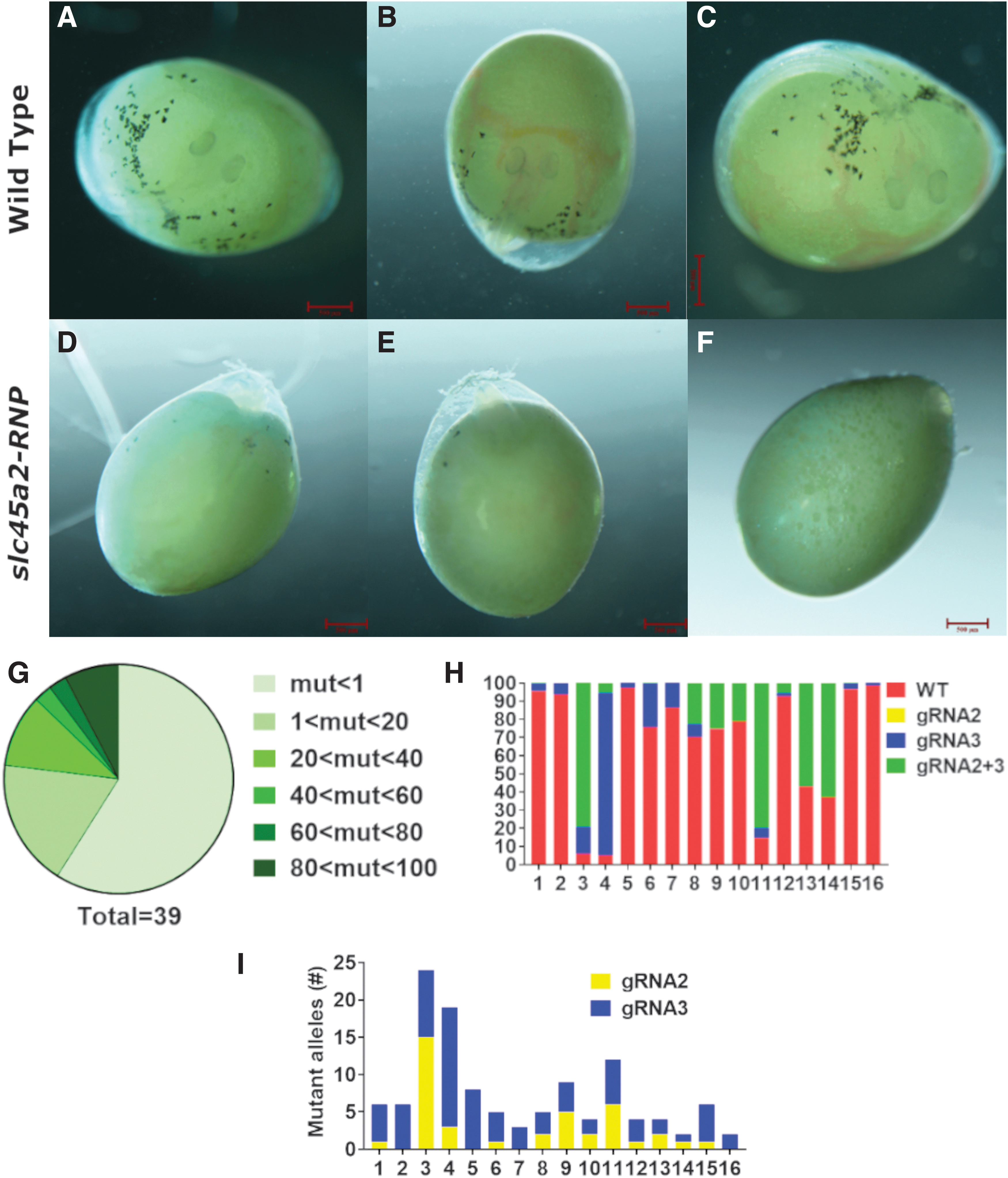
Transient analysis of slc45a2-RNP activity in vivo. Nile tilapia zygotes were microinjected at the single-cell stage with RNPs containing slc45a2-exon1-specific gRNAs. At 4 dpf, naïve embryos exhibited melanin formation in the eye and clear melanophores on the body and yolk surface
Aiming to gain higher resolution of the editing events in injected fish, the genomic target region of gRNA2 and gRNA3 from multiplex-injected fish from four different injection sessions was subjected to amplicon sequencing. This analysis revealed variable levels of mutagenic activity in all injected fish (Fig. 2G). However, all fish displayed mutant alleles (Supplementary Table S1). Analysis of the general mutagenic outcome showed that most mutant alleles resulted from the co-activity of both gRNAs or from sole activity of gRNA3, whereas alleles resulting from exclusive activity of gRNA2 were hardly detected (Fig. 2H). Further analysis of gRNA-specific mutagenic activity showed that the lower activity of gRNA2 was also accompanied by significantly fewer mutagenic outcomes (Fig. 2I). Nonetheless, the combined activity of both gRNAs resulted in the generation of 70 unique alleles (Supplementary Table S2).
Generation of germline with slc45a2 mutant alleles
Following our analysis of the transient effects of slc45a2 loss of function, we next aimed to induce heritable null mutations in tilapia slc45a2. It was previously demonstrated in zebrafish that multiplexing can increase the mutagenesis throughput and that off-target mutagenesis is of low concern when using the CRISPR-Cas9 transiently in fish zygotes. 21 Our initial implementation of gRNA1 elicited no phenotype in the injected embryos. Additional Sanger sequencing of 10 random injected larvae confirmed that no genomic indels were generated (data not shown). Hence, we targeted slc45a2-exon1 by co-injection of RNPs containing gRNA2 and gRNA3 (Fig. 3A) into Nile tilapia zygotes. As with triple slc45a2-RNP multiplexing, co-injected embryos displayed strong reduction in melanin throughout their body and eyes. While non-injected siblings exhibited normal pigment development at 1 month post fertilization (Fig. 3B), one of the injected F0 fish displayed 97–99% melanin loss (Fig. 3C) suggestive of very strong mutagenic activity during early development. The oculocutaneous albinism (OCA) phenotype, that is, complete loss of melanin, persisted into sexual maturation of this male (Fig. 3D).

Phenotypic analysis of slc45a2-RNPs-induced mutations in adult tilapia. Nile tilapia zygotes were microinjected at the single-cell stage with RNPs containing slc45a2-exon1-specific gRNAs 2 and 3
Next, we aimed to test for possible correlation between slc45a2-RNP-induced somatic and germline indels. The target region of slc45a2-exon1 was amplified using gDNA extracted from fin clips and sperm of the albino male and cloned for individual sequencing. Analysis of 10 colonies from fin-clip gDNA and 18 colonies from sperm gDNA identified six different indels in the fin and eight (seven mutants + WT) different products in the sperm, of which two were identical between fin and sperm (Fig. 4A and B), suggesting a low to moderate correlation between somatic and germ-cell events. Nonetheless, as sperm was extracted by application of mild physical pressure, some of the identified indels in this sample might result from gDNA that originated from somatic cells of the gonad. Thus, the progenitor male was crossed with WT females, and sequencing analysis of fin clips from F1 fish was performed. This analysis demonstrated that all of the mutant alleles identified in sperm, as well as the WT allele, existed in F1 heterozygous offspring (Fig. 4C). Sequencing analysis of gDNA from the F0 albino mutant and its heterozygous F1 offspring clearly showed that our novel slc45a2 mutant alleles are heritable and stable.

Molecular analysis of slc45a2-RNP-induced somatic and germline mutations in Nile tilapia. Genomic slc45a2-exon1 was amplified using gDNA extracted from F0 fin clips
HRM analysis and F2 phenotyping
F1 fish carrying the slc45a2–/+ genotype displayed no altered pigmentation relative to their WT siblings (Supplementary Fig. S3), which raised the need for an efficient method to sort F1 offspring according to their genotype. HRM was previously shown as an effective method to detect genome editing-driven mutations.39–42 Due to its low cost, high throughput, and high sensitivity,41,43 we opted to test its application for the assessment of allele inheritance frequencies in slc45a2–/+ F1 populations. For this, gDNA extracted from the tail clips of 330 F1 offspring spawned by five random females were analyzed using the HRM method. This analysis revealed the existence of all mutant alleles that we identified in sperm. Surprisingly, melt curves of some analyzed embryos did not match any of the reference DNAs of known alleles. Sanger sequencing of the target region in these individuals confirmed the existence of three additional mutant alleles within the F1 population. Thus, we identified 19 different heritable mutant alleles (Fig. 4C). These results emphasize the high throughput and accuracy of this assay for CRISPR-cas9 editing population assessment.
Allele frequencies in the F1 population revealed differential allele heredity (Fig. 5A). Just over a quarter (26.7%) of the F1 population was characterized as WT. While allele O5 (a single-site indel) displayed the highest inheritance frequency (16.7%), allele O8, which probably resulted from multiplex RNP activity, displayed the lowest allele frequency (1.8%; Fig. 5A). Interestingly, other mutant alleles exhibited frequencies of 5.8–11.5% in the F1 population, with no clear correlation between the genomic CRISPR-Cas9 outcome and its level of inheritance (Supplementary Table S3). Despite the high sensitivity of our HRM analysis, we could not differentiate between alleles O7 and O10, which differed by two nucleotides. Therefore, these two alleles are presented together. Analysis of genomic outcomes in each individual site revealed that although both RNPs were microinjected at an identical molar ratio, at the level of germline transmission, site 3 showed higher mutagenic response (73% of the F1 offspring), while site 2 exhibited ∼50% heritable offspring (Fig. 5B and C and Supplementary Table S3). Phenotypic analysis of F2 embryos at 5 dpf showed OCA phenotype with Mendelian inheritance (Fig. 5D–F).
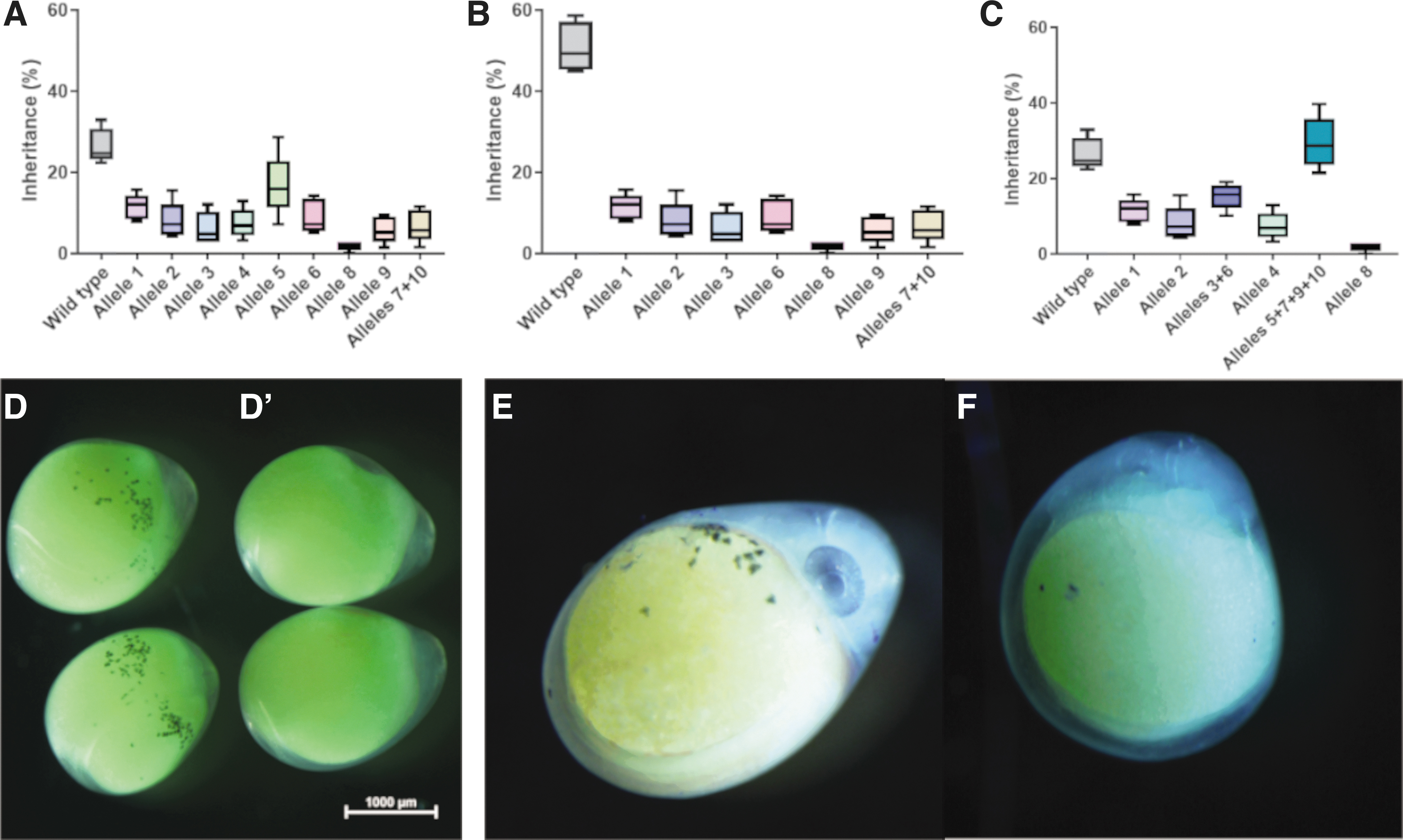
Allele frequency and phenotypic analysis of slc45a2 mutants. High-resolution melt analysis of sequence validated slc45a2 mutant alleles in F1 larvae showed that most alleles shared similar heredity level apart of the highly abundant allele 5 and the relatively rare allele of complete site-to-site deletion
Discussion
Red tilapia is of higher market value and demand than the WT phenotype. However, currently available strains exhibit black and dark-red blotching that reduce their market value, and their unclear genetic background impairs breeding and culture practices.2,3,44 Impairments in SLC45A2 in humans lead to OCA, 12 which is also phenocopied in other vertebrates, including fish.9,15 Additionally, it was previously suggested that red tilapia phenotypes result from dermal blood irrigation within a melanophore-perturbed skin. 6 In the current study, we have identified and partially cloned the slc45a2 gene in Nile tilapia. Then, we used these data to generate highly specific gRNAs for its genomic exon 1 and showed that when these gRNAs are multiplexed as RNPs and delivered into Nile tilapia zygotes, they induce OCA in this species, resulting in a solid-red phenotype. Lastly, we demonstrated the generation of several germline-transmitted null slc45a2 alleles, their screening in a mixed allele population using HRM, and the use of allele-specific F1 fish for the establishment of stable and genetically trackable solid-red tilapia germlines.
Pigment synthesis is a complex process that involves multiple genes, most of which have multiple copies as a result of fish-specific genomic duplications. However, the genes slc45a2, oculocutaneous albinism type 2 (oca2), and solute carrier family 24 member 5 (slc24a5; also known as OCA6) are exceptional among the melanin-related genes of teleosts, having only a single copy. 11 Therefore, we hypothesized that loss-of-function mutations in slc45a2 would result in albinism. Our bioinformatic analysis demonstrated that Nile tilapia possess a single slc45a2 gene that shares its evolutionary origin and chromosomal synteny with slc45a2 genes of other vertebrates. While SLC45A2 mutations are recessive, 12 most identified loci regulating red tilapia phenotypes are genetically dominant. 16 Furthermore, the recessive “blonde” allele of Nile tilapia was located to chrLG5, and other dominant red and red-blotch alleles were associated with chrLG3, chrLG5, and chrLG15 and to the slc45a1 gene, but not in chrLG7 where tilapia slc45a2 is located. 16 In addition, our slc45a2 mutant tilapia does not express melanin in its eyes or have black blotched skin, which characterize the currently known red tilapia strains.2,3,6,16 These data suggest that current tilapia strains of red phenotypes do not result from perturbation in the slc45a2 gene or protein. Furthermore, previously identified strains of Mozambique “gold” 45 or Nile “blond” and “syrup” 46 tilapias, as well as hybrid strains, all suffer from various levels of WT coloration or blotched progenies, which resulted from impaired melanophore development.6,46 Null slc45a2 alleles were suggested to result in impaired melanin synthesis due to perturbed melanosome activity, leading to OCA.12,14,15 In this view, we chose slc45a2 as our target gene with the aim of impairing general melanin synthesis and thereby generating a solid, non-blotched red phenotype. Although the OCA phenotype level of our F0 fish varied, at least some of the fish exhibited robust melanin loss in both skin and eyes. NGS analysis demonstrated that all injected F0 fish carried mutated alleles in their genome, although as seen in the OCA phenotype, mutation levels varied. Furthermore, our NGS analysis revealed that while almost all mutated alleles resulted from sole activity of gRNA3 or joint activity of gRNA2 with gRNA3, almost no alleles were generated by the sole activity of gRNA2. As this result was consistently found in all injected fish, we suggest that successful activity of gRNA2 is dependent on the efficiency of gRNA3. This could result from chromatin relaxation driven by a gRNA3-dependent double-strand break, which due to the high genomic proximity with the target site of gRNA2 reduces the stringency and allows its activity. Nonetheless, this possibility requires further testing by gRNA multiplexing array.
Analysis of the mutations induced by slc45a2 RNPs showed a mild correlation between somatic and germ-cell alleles in the genomic consequences of the double-strand break induced by Cas9. RNPs microinjection is performed into single-cell zygotes. Yet, only four cells of the embryo are germ-cell committed until the 1,000 cells stage. At this stage, the RNPs are highly diluted and expected to have no mutagenic activity.21,47 Surprisingly, our HRM analysis demonstrated the presence of 10 heritable alleles, which was unexpected, as previous high-throughput work in zebrafish identified a maximum of eight alleles in the fish germline. 21 Whether this difference resulted from variation among species in germline development or increased stability of the RNP complex at lower temperatures remains to be determined. Comparing CRISPR-Cas9-induced somatic mutations in whole fish and fin clips of salmon demonstrated that fin-clip analysis mirrored somatic mosaicism and therefore can serve as an estimate for the knockout phenotype. 17 In this view, the phenotype seen in the F0 tilapia, together with the somatic and germline mutagenesis levels, suggested that the phenotype of slc45a2–/– would be equal to the phenotype seen in the injected fish. Indeed, F2 slc45a2–/– tilapia displayed a clear OCA phenotype and no melanin in their skin or eyes. Previous works in catfish demonstrated that albino fish have similar growth rate to their pigmented siblings, yet they display reduced aggressiveness and shoaling behaviors.48,49 In trout, yellow albinos retain their xanthophores and display normal growth, while the loss of xanthophores results in reduced growth. 50 As slc45a2–/– tilapias displayed the exclusive loss of melanophores, we assume that they will display a normal growth rate. However, this assumption requires further investigation.
Taken together, our findings highlight the applicative potential of the CRISPR-Cas9 system in Nile tilapia culture. Moreover, Nile tilapia is one of the most widely cultured species in global aquaculture and is often hybridized with other Oreochromines.44,51 Thus, the generation of heritable and trackable slc45a2–/– alleles with a red phenotype in Nile tilapia may affect the global tilapia aquaculture. Nonetheless, this would be tested by the attitude of farmers, consumers, and regulators toward consumption of genome-edited fish.
Footnotes
Acknowledgment
We thank Daniela Ben Tov and Aviva Samach from the laboratory of Prof. Avraham Levy, Weizmann Institute of Science for their expertise and advice in NGS data analysis.
Author Disclosure Statement
The tilapia germline has been submitted for provisional patent registration (U.S. application #63/113,892).
Funding Information
This research was funded by grant #20-04-0048 from the Chief Scientist of the Ministry of Agriculture and Rural Development, Israel.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.