
Abstract
Anti-CRISPR (Acr) proteins are phage-borne inhibitors of the CRISPR-Cas immune system in archaea and bacteria. AcrIIC2 from prophages of Neisseria meningitidis disables the nuclease activity of type II-C Cas9, such that dimeric AcrIIC2 associates with the bridge helix (BH) region of Cas9 to compete with guide RNA loading. AcrIIC2 in solution readily assembles into oligomers of variable lengths, but the oligomeric states are not clearly understood. In this study, we investigated the dynamic assembly of AcrIIC2 oligomers, and identified key interactions underlying the self-association. We report that AcrIIC2 dimers associate into heterogeneous high-order oligomers with the equilibrium dissociation constant KD ∼8 μM. Oligomerization is driven by electrostatic interactions between charged residues, and rational mutagenesis produces a stable AcrIIC2 dimer with intact Cas9 binding. Remarkably, the BH peptide of Cas9 is unstructured in solution, and undergoes a coil-to-helix transition upon AcrIIC2 binding, revealing a unique folding-upon-binding mechanism for Acr recognition.
Introduction
Clustered regularly interspaced short palindromic repeat (CRISPR) and CRISPR-associated (Cas) proteins constitute the adaptive immune system in archaea and bacteria against invading bacteriophages and plasmids.1,2 As a countermeasure, phages employ anti-CRISPR (Acr) proteins to inhibit the host CRISPR-Cas defense system. 3 Acr proteins were first identified in the bacteriophages resistant to the type I-F CRISPR-Cas system of Pseudomonas aeruginosa.4,5 Acr proteins typically lacked sequence homology between themselves, but were strongly associated with a downstream helix-turn-helix motif, referred to as the anti-CRISPR-associated (aca) gene. 6 Bioinformatics analysis based on aca genes lead to the discovery of Acr proteins against the type II-C CRISPR-Cas system in prophages of Neisseria meningitidis, revealing AcrIIC1, AcrIIC2, and AcrIIC3 proteins—the first-in-class Cas9 inhibitors. 7
AcrIIC2 was discovered in the prophage of N. meningitidis in the neighborhood of the aca3 gene, inhibiting the nuclease activity of type II-C Cas9 in vitro and in cellulo. 7 Crystal structures demonstrated that an AcrIIC2 dimer interacted with the bridge helix (BH) of Cas9, occluding the guide RNA binding site.8,9 Although the AcrIIC2 dimer was sufficient for Cas9 inhibition, AcrIIC2 appeared to aggregate as large oligomers in solution, with undefined subunit stoichiometry. 8 In this study, we investigated the AcrIIC2 oligomerization in solution by light scattering and spectroscopic measurements. Light scattering data revealed that AcrIIC2 dimers self-associated into high-order oligomers in a concentration-dependent manner, with the equilibrium dissociation constant (KD) estimated as ∼8 μM. We then identified key electrostatic interactions for the oligomerization, such that surface charge mutations effectively promoted a stable dimer. We employed the engineered AcrIIC2 dimer to investigate the target recognition in solution, demonstrating that AcrIIC2 induces a coil-to-helix transition in the isolated BH peptide of Cas9 upon complex formation.
Materials and Methods
Cloning, expression, and purification
The synthetic acrIIC2 gene was cloned into a pBT7 vector and transformed into the Escherichia coli strain BL21(DE3) (Invitrogen). Cells were grown in lysogeny broth (LB) medium at 37°C until the optical density at 600 nm reached 0.6–0.8, and induced by 1 mM isopropyl-β-
N-terminal His6-tagged N. meningitidis Cas9 in the pMCSG7 vector was purchased from Addgene. The vector was transformed into the E. coli strain Rosetta(DE3) (Invitrogen). Cells were grown in LB medium at 37°C to an optical density of 0.6–0.8 at 600 nm, and induced by 1.0 mM IPTG for 16 h at 16°C. Cells were harvested by centrifugation and resuspended in 20 mM Tris-HCl, pH 7.4, 500 mM NaCl, 5% glycerol, and 5 mM BME, 1 mM PMSF supplemented with 1 mg/mL lysozyme and protease inhibitor cocktail. After lysis by using EmulsiFlex-C3 (AVESTIN) and centrifugation, the supernatant was loaded onto a HisTrap column (GE Healthcare) and eluted with a 0–500 mM imidazole gradient. The proteins were further purified using a HiTrap Heparin column (GE Healthcare) with 0–1 M NaCl gradient, and further with the Superdex 200 26/600 column (GE Healthcare) in 20 mM Tris-HCl, pH 7.4, 200 mM NaCl, and 5 mM BME.
The BH (a.a. 55–91 of N. meningitidis Cas9) gene was cloned into a pET15b vector engineered with an N-terminal His6–MBP tag followed by the TEV protease cleavage site, and transformed into the E. coli Rosetta(DE3) cells. Cells were grown in LB medium at 37°C until the optical density at 600 nm reached 0.6–0.8, and induced by 0.5 mM IPTG for 20 h at 16°C. Cells were harvested by centrifugation and resuspended in 20 mM Tris-HCl, pH 7.4, 100 mM NaCl, 2 mM BME, and 1 mM PMSF. After lysis by EmulsiFlex-C3 (AVESTIN) and centrifugation, the supernatant was loaded onto a 5-mL HiTrap Heparin column (GE Healthcare) pre-equilibrated with 20 mM Tris-HCl, pH 8.8, 100 mM NaCl, and 2 mM BME, and the bound proteins were eluted by a linear gradient up to 2 M NaCl. The proteins were then loaded onto a 5-mL HisTrap HP column equilibrated with 20 mM Tris-HCl, pH 8.8, 300 mM NaCl, and 2 mM BME, and eluted by a linear gradient of 20–500 mM imidazole. The His6–MBP tag was cleaved by the TEV protease and removed using a 5-mL HiTrap SP column (GE Healthcare) to obtain the intact BH peptide.
Multi-angle light scattering
Multi-angle light scattering (MALS) was measured at 25°C on a TSK-gel-G3000SWXL column (0008541; Tosoh Bioscience, LLC). Size exclusion chromatography (SEC)-MALS experiments were performed using an Agilent 1200 high performance liquid chromatography System (Agilent Technologies) combined with a Wyatt DAWN Heleos II and a Wyatt Optilab T-rEX differential refractometer (Wyatt Technology). The equipment was precalibrated with 1 mg/mL bovine serum albumin (66 kDa). The samples were eluted in 20 mM Tris-HCl, pH 7.4, 200 mM NaCl, and 2 mM BME, and the results were normalized and analyzed using the Astra 6 software (Wyatt Technology).
Estimation of the dissociation constant for the AcrIIC2 self-association
We estimated the dissociation constant for the AcrIIC2 self-association, based on the assumption that a single equilibrium constant governs the progressive association of AcrIIC2 dimers into larger oligomers. The total concentration of AcrIIC2 was determined experimentally by the absorbance at 280 nm. Initially, two AcrIIC2 dimers form a tetramer (Reaction 1), and then a dimer and a tetramer make a hexamer (Reaction 2).
Dimer + Dimer ⇄ Tetramer (Reaction 1)
Dimer + Tetramer ⇄ Hexamer (Reaction 2)
We employed two quadratic equations: one for Reaction 1, and the other for Reaction 2. When initial concentrations of dimer and tetramer are given as d and t, respectively, and the final tetramer concentration is given as t + x, Reaction 1 is described as follows:
Dimer + Dimer ⇄ Tetramer
Initial concentration: d d t
Final concentration: d − 2x d − 2x t + x

Next, initial concentrations of dimer, tetramer, and hexamer are given as d, t, and h, respectively, and the final hexamer concentration is given as h + x. Then, Reaction 2 is described as follows:
Dimer + Tetramer ⇄ Hexamer
Initial concentration: d t h
Final concentration: d − x t − x h + x

The two equations allow for the calculation of AcrIIC2 dimer, tetramer, and hexamer, if the total AcrIIC2 concentration and a KD value are provided. In brief, the dimer and tetramer concentrations are obtained from Reaction 1, and the hexamer concentration is obtained from Reaction 2. To obtain the concentration of each species in full equilibrium, we reiterate the calculation of Reactions 1 and 2 in a recursive manner, such that each output feeds as the input of the other equation until the concentrations no longer vary in the next round of calculations. When the system reaches the equilibrium, we calculate the population-weighted average molecular mass and compare this with the experimental value from the MALS measurement. A grid search is used to find the KD value that best matches the calculated mass with the experimental one. We used the Microsoft Excel program to calculate the equilibrium concentration and the average molecular mass. The oligomerization reaction can be extended in principle to take account of higher-order oligomers such as octamer and decamer.
We analyzed MALS data of the 140 μM injection of AcrIIC2, and employed the dimer, tetramer, and hexamer as major components in equilibrium. Considering that a 10-fold dilution took place during the MALS measurement (∼1 mL of elution compared with 100 μL of injection volume), there was ∼14 μM of total AcrIIC2 (or ∼7 μM of the AcrIIC2 dimer) in the elution of the 140 μM injection. Concentrations of the three oligomeric components were determined by iteratively calculating quadratic equations with a fixed total dimer concentration (7 μM) and a grid search of the equilibrium constant.
Circular dichroism spectroscopy
Circular dichroism (CD) spectroscopy was conducted at 25°C using a J-815 CD spectrometer (Jasco Products). Wave scans were acquired by sampling data at 0.5 nm intervals between 190 and 260 nm for far ultraviolet (UV) CD measurement. Far UV CD spectroscopy was carried out with 10 μM of protein in 20 mM sodium phosphate, pH 7.4, 200 mM NaCl, and 2 mM BME using a 1-mm quartz cuvette. The far UV CD spectrum was obtained from an average of three scans and the results were presented as the molar ellipticity (deg·cm2/dmol) at each wavelength.
Single guide RNA preparation
DNA encoding a minimal T7 promoter upstream of a single guide RNA (sgRNA) of N. meningitidis Cas9 (with a random sequence without targeting sites in E. coli: 5′-TAATACGACTCACTATAGGGTGCGCGGCGCATTACCTTTACGTTGTAGCTCCCTTTCTTATTTCGGAAACGAAATGAGAACCGTTGCTACAATAAGGCCGTCTGAAAAGATGTGCCGCAACGCTCTGCCCCTTAAAGCTT-3′) was synthesized by Bioneer. The DNA template for RNA transcription was prepared by using the Gigaprep kit (Epigenetics). The sgRNA was prepared in vitro by mixing rNTPs (rATP, rGTP, rCTP, and rUTP), MgCl2, T7 RNA polymerase (P266L mutant), inorganic pyrophosphatase, and the DNA template in the transcription buffer. After 6 h of transcription at 37°C, synthesized RNA was precipitated by ethanol treatment overnight, purified using 12% denaturing polyacrylamide gel electrophoresis (19:1 cross-linking ratio), and electro-eluted using the Elutrap electroelution system (Whatman). Purified RNA was desalted and exchanged into water using Amicon Ultra centrifugal filters (Merck Millipore).
In vitro cleavage assays
NmeCas9 (2.5 μM) was incubated with AcrIIC2 of 2.5, 5, 12.5, and 25 μM concentrations in 20 mM Tris-HCl, pH 7.5, 10 mM KCl, 5 mM MgCl2, 1 mM DTT, and 5% glycerol for 10 min at 37°C. sgRNA (2.5 μM) was then added to the NmeCas9–AcrIIC2 mixture for 10 min at 37°C. Next, the DNA substrate linearized by ScaI digestion was added to a final concentration of 5 nM for 30 min at 37°C. Finally, the reaction products were incubated with proteinase K for 10 min at 50°C, and then visualized on a 1% agarose gel.
Crystallization, X-ray data collection, and structure refinement
The protein crystals of AcrIIC2 were grown using the sitting-drop vapor diffusion method at 20°C by mixing 0.4 μL of 16 mg/mL AcrIIC2 in 20 mM sodium phosphate, pH 7.4, and 2 mM BME, with the same volume of 10 mM Tris-HCl, pH 7.0, 30% (v/v) 2-propanol, and 0.3 M ammonium acetate. 20% (v/v) glycerol was used as a cryoprotectant for data collection. The crystal was vitrified in the nitrogen gas stream and the data were collected using a Pilatus3 6M CCD area detector (Dectris) at the BL-11C experimental station of Pohang Light Source, Korea. Crystals of AcrIIC2 belong to the hexagonal space group P6522, with unit cell parameters of a = b = 71.4 Å, c = 316.6 Å, α = β = 90°, and γ = 120°. All raw data were processed and scaled using the HKL2000 program package. 10 The data collection and refinement statistics are summarized in Supplementary Table S1.
A set of single-wavelength anomalous dispersion data from a crystal of the SeMet-substituted AcrIIC2 was used to solve the phase problem at a resolution of 2.62 Å. Phase calculation, density modification, and initial model building were carried out using PHENIX AutoSol and AutoBuild of the PHENIX program suite. 11 The models were further built manually using the program COOT, 12 and refined with the program REFMAC, 13 including the bulk solvent correction. Water molecules and ligands were added using the program COOT and were manually inspected. 5% of the data were randomly set aside as the test data for the calculation of Rfree. 14 The stereochemistry of the final structures was evaluated using MolProbity. 15
Nuclear magnetic resonance experiments
The nuclear magnetic resonance (NMR) samples were prepared in 20 mM sodium phosphate, pH 7.4, 200 mM NaCl, 2 mM BME, and 0.01% NaN3. 1 H– 15 N heteronuclear single quantum correlation (HSQC) spectra of 0.3 mM 15 N-AcrIIC2 and 15 N-AcrIIC2R64D,R65D were recorded at 25°C on a Bruker 850 MHz spectrometer equipped with a z-shielded gradient triple resonance cryoprobe. For the BH of Cas9, 0.1 mM 15 N-BH was titrated with unlabeled AcrIIC2R64D,R65D in a stoichiometric manner. The NMR spectra were processed and analyzed using NMRPipe 16 and NMRView 17 programs.
Isothermal titration calorimetry
Isothermal titration calorimetry experiments were performed at 25°C using an iTC200 calorimeter (Malvern). Twenty-four micromolars of AcrIIC2R64D,R65D or wild-type AcrIIC2 in the sample cell was titrated with 160 μM BH in 20 mM Tris-HCl, pH 7.4, and 200 mM NaCl. Twenty consecutive 2 μL aliquots of BH were titrated into the cell, with an injection duration of 4 s and an inter-injection interval of 150 s. The heats associated with dilution of the titrants were subtracted from the measured heats of binding. The titration data were analyzed using the Origin version 7.0 program provided with the instrument.
Results
AcrIIC2 self-associates into large oligomers in solution
We noticed that AcrIIC2 appeared as a large molecular-weight oligomer from the SEC during purification, and questioned the composition of the oligomeric state. Purified AcrIIC2 eluted significantly earlier in the analytical SEC measurement of 500 μM injection concentration compared with that of 25 μM injection, indicating the presence of homomeric association (Fig. 1A). From the peak elution volume and the calibration with standard proteins, the molecular weights corresponded to 45.2 and 27.6 kDa for 500 and 25 μM injections, respectively (Fig. 1A, inset). We proceeded to measure the absolute molecular mass of AcrIIC2 oligomers to determine the subunit stoichiometry using MALS experiments. Light scattering and refractive index measurements revealed heterogeneous oligomeric distributions at different protein concentrations (Fig. 1B). AcrIIC2 (calculated molecular mass of 14,164 Da) upon 140 μM injection concentration showed a distribution of molecular mass ranging from 28 to 72 kDa, yielding the average mass as 41.1 ± 11.9 kDa. The trailing shoulder on the peak and a wide range of measured molecular mass indicated a dynamic equilibrium between different oligomers. Since the lowest mass from MALS corresponded to an AcrIIC2 dimer (∼28 kDa), the AcrIIC2 dimer was likely the basic building block of higher-order oligomers. The measured molecular mass distribution (28–72 kDa) could then be explained by a rapid equilibrium between AcrIIC2 dimers, tetramers, and hexamers. When we elevated the protein concentration, higher-order oligomers appeared with a population shift toward larger molecular weight species. The molecular masses of growing higher-order oligomer fractions were determined as 72.0 ± 7.8, 84.4 ± 10.5, and 102.5 ± 19.8 kDa from the measurement of 350, 490, and 700 μM injection concentrations, respectively. Notably, the lower end of the molecular mass distribution remained as a dimer regardless of the concentration, whereas the upper end continuously increased, such that the largest molecular mass of ∼132 kDa from 700 μM injection concentration corresponded to an AcrIIC2 decamer (Fig. 1B).
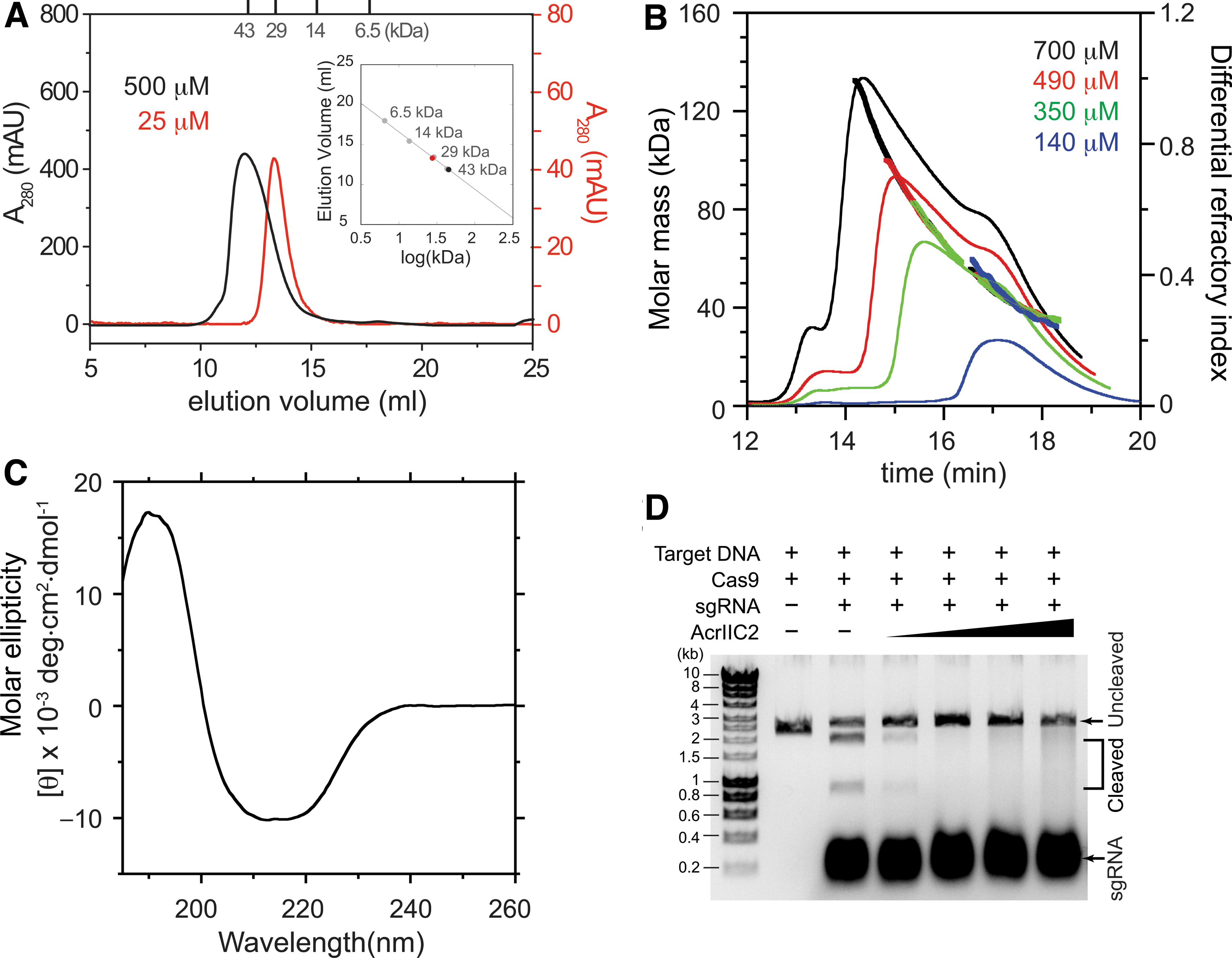
Characterization of oligomeric states, secondary structures, and Cas9 inhibition of AcrIIC2.
The self-association of AcrIIC2 did not originate from aggregation of misfolded AcrIIC2, since the oligomer fractions maintained the secondary structure in the CD spectra (Fig. 1C). In addition, AcrIIC2 diluted from higher oligomeric states (700 μM) effectively inhibited the nuclease activity of N. meningitidis Cas9, supporting that the oligomers were not induced by misfolding (Fig. 1D). In contrast, disruption of the dimer interface previously abrogated the Acr activity of AcrIIC2. 8 Taken together, AcrIIC2 forms a tight dimer for Cas9 inhibition at low concentrations, and the dimer progressively self-associates in a reversible manner upon elevated concentrations.
We then calculated the equilibrium dissociation constant (KD) for the interaction between AcrIIC2 dimers, assuming that a single equilibrium constant governs the overall oligomerization process. The average molecular mass of 41.1 kDa from MALS measurement of 140 μM injection corresponded to ∼1.5 AcrIIC2 dimers per oligomer (average association number). We analyzed the molecular mass distribution of AcrIIC2, employing the dimer, tetramer, and hexamer as major components in equilibrium (see Materials and Methods section for the analysis procedure in detail). Our numerical analysis showed that the KD value of ∼8 μM generated the AcrIIC2 dimer, tetramer, and hexamer populations as 65%, 25%, and 10%, respectively, which best reproduced the experimental association number of 1.5.
Rational mutagenesis prevents AcrIIC2 oligomerization
Recent crystal structures from two independent groups unambiguously reported the AcrIIC2 dimer as the functional state. We examined the crystal contacts between AcrIIC2 dimers to find potential interfaces for the oligomerization. When we generated the symmetry mates from the structural co-ordinate, we could identify electrostatic interactions that assembled three dimers in a triangular shape (Fig. 2A). Specifically, Asp41 and Asp42 of one dimer subunit formed multiple hydrogen bonds and electrostatic interactions with Arg64 and Arg65 of another dimer subunit, such that three dimers associated in a heterologous manner (Fig. 2B, C). Same interactions were observed in two independent crystals, one from 0.1 M sodium citrate, 5% propanol, 18% PEG 4000, and 15% glycerol (space group P41212), and the other from 0.1 M Tris, pH 8.5, and 20% (v/v) ethanol (space group P6322). We also obtained our own AcrIIC2 crystal using 10 mM Tris-HCl, pH 7.0, 0.3 M ammonium acetate, and 30% (v/v) 2-propanol (space group P6522), and again the crystal structure recapitulated aforementioned dimer–dimer contacts. For the record, we determined our crystal structure to a resolution of 2.2 Å, and the data collection and refinement statistics are summarized in Supplementary Table S1. Our crystal structure reproduced the same backbone fold of the dimer, featuring a six-stranded antiparallel β-sheet flanked by two α-helices at N- and C-termini (Fig. 2D). Asp41 and Asp42 were located in the β3–β4 loop at the end the of β3 strand, whereas Arg64 and Arg65 were in the β4–β5 loop at the beginning of the β5 strand (Fig. 2F). These charged residues were located opposite the dimerization interface within each AcrIIC2 dimer (Fig. 2D). In addition, they did not contribute to the binding interface of AcrIIC2 for the BH domain to inhibit Cas9 (Fig. 2E), and as such, mutating Asp41 and Asp42 had little effect on the Acr activity of AcrIIC2.8,9
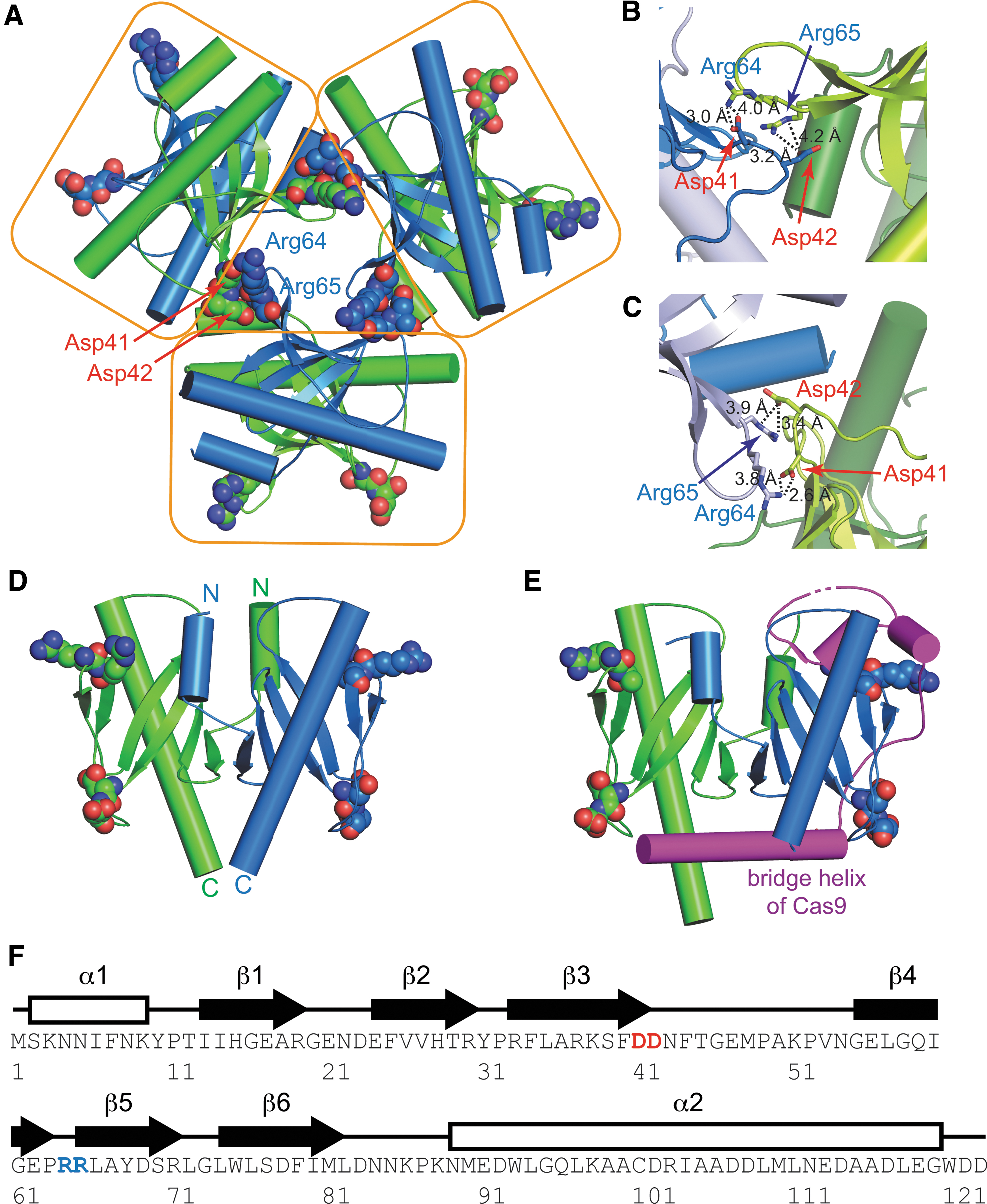
Crystal structures of
We hypothesized that the electrostatic interactions between dimers were responsible for the self-association of AcrIIC2 in solution. When we mutated Arg64 and Arg65 to aspartates, the AcrIIC2R64D,R65D mutant indeed formed a stable dimer in SEC. The light scattering of AcrIIC2R64D,R65D illustrated that the mutant remained as a constant dimer in the range of 70–700 μM injection concentrations, which contrasts the apparent increase of larger oligomers observed for wild-type AcrIIC2 (Fig. 3A). AcrIIC2R64D,R65D was fully capable of inhibiting the nuclease activity of N. meningitidis Cas9 similar to wild-type AcrIIC2 (Fig. 3B). We then examined the 1 H– 15 N HSQC NMR spectra of 15 N-labeled AcrIIC2 and AcrIIC2R64D,R65D. AcrIIC2 showed only a limited number of backbone amide resonances with severe line broadening that was attributed to larger molecular weights and chemical exchanges of oligomers (Fig. 3C). In contrast, AcrIIC2R64D,R65D showed well-dispersed backbone amide resonances of a folded protein, which supports the absence of oligomerization. Observed amide resonances in the HSQC spectra superimposed well between AcrIIC2 and AcrIIC2R64D,R65D, indicating that the backbone fold remained intact in the mutant (Fig. 3C). We also measured the binding thermodynamics of AcrIIC2R64D,R65D and the BH peptide of N. meningitidis Cas9 using isothermal titration calorimetry, and obtained the KD value of 73 ± 22 nM, with a 1:1 stoichiometry between the AcrIIC2R64D,R65D dimer and the BH peptide (Fig. 3D). The binding affinity was similar to the KD value (23 ± 9 nM) obtained between wild-type AcrIIC2 and the BH peptide (Supplementary Fig. S1). Thus, AcrIIC2R64D,R65D represented a functional dimer with Cas9 binding as competent as native AcrIIC2. Our result further confirmed that a compact BH peptide (a.a. 55–91 of N. meningitidis Cas9 in this study) was sufficient to elicit potent AcrIIC2 binding as was observed with a longer version (a.a. 51–123) in the previous measurement. 8

Changes in oligomeric states of AcrIIC2 and its mutant.
AcrIIC2 induces a coil-to-helix transition in the BH peptide
The BH domain of Cas9 adopts an extended helical conformation that connects the nuclease lobe for target cleavage and the recognition lobe for guide RNA binding. Previous complex structures contained co-ordinates of AcrIIC2 and a truncated BH peptide, partly due to proteolytic degradations of Cas9 during sample preparation. Residues 55–78 of the BH domain showed a native helical conformation, whereas the rest of BH (residues 79–91) lacked an electron density map required for structure determination.8,9 This is not unusual, since peptides are often disordered in solution and adopt secondary structures upon binding to cognate target proteins. We questioned whether the AcrIIC2 binding would induce the helical conformation of the BH peptide. The secondary structure of the BH peptide was monitored in the presence and absence of AcrIIC2 using CD spectroscopy. Interestingly, the BH peptide alone was largely unstructured in solution, but readily switched into a helix upon AcrIIC2 binding (Fig. 4A). Subtraction of CD spectra between the BH–AcrIIC2 complex and free AcrIIC2 manifested a large increase in the ellipticity minimum at 208 nm, indicating the helix formation of BH upon complex formation (Fig. 4A). We note that the crystal structure of AcrIIC2 remained unchanged regardless of BH binding,8,9 thus the CD difference indicates the secondary structure formation of the BH peptide. We further examined the HSQC spectra of 15 N-labeled BH before and after AcrIIC2 binding. Free 15 N-BH showed narrow chemical shift dispersion of amide resonances, a common feature of unstructured polypeptides (Fig. 4B). In contrast, titrations with unlabeled AcrIIC2 lead to significant chemical shift perturbations and widening dispersion, supporting a coupled folding and binding mechanism of the BH peptide.

Conformational change of the BH peptide upon binding to AcrIIC2.
Discussion
AcrIIC2 is an acidic protein with the isoelectric point of 4.7. The electrostatic surface potential reveals highly polarized charge distributions, such that positively charged surfaces appear on one side of the AcrIIC2 dimer and negatively charged surfaces on the other side (Supplementary Fig. S2A). We propose a possible model of the AcrIIC2 oligomerization driven by intermolecular electrostatic interactions. In our model, three AcrIIC2 dimers interact with one another in a head-to-tail manner through electrostatic interactions between opposite surface charges, leading to a closed triangular hexamer (Supplementary Fig. S2B). Each dimer–dimer interface buries overall ∼1,374 Å 2 of solvent accessible surface area within the hexamer. Our model features that the dimer subunits in the hexamer are able to engage with new incoming dimers through their charged residues facing outward from the hexamer (Fig. 2A and Supplementary Fig. S2B). Thus, the hexamer would associate progressively with new dimers toward higher-order oligomers, which is limited by the total protein concentration in solution.
Acr proteins targeting the type II CRISPR-Cas system have been actively investigated, and there are currently 23 Acr proteins against the subtype II-A CRISPR-Cas, and 5 Acr proteins against the subtype II-C CRISPR-Cas system.18–20 Structural and mechanistic studies revealed that type II Acrs inhibit their host Cas9 proteins through diverse mechanisms. They occupy the protospacer adjacent motif interaction site of Cas9 to compete with target DNA binding,21,22 block the active site of the Cas9 nuclease domain, 23 induce nonfunctional Cas9 dimerization,24,25 and cause Cas9 degradation in cell culture. 26 AcrIIC2 and AcrIIA15 are uniquely known to interfere with the sgRNA loading onto Cas9.8,9,27 Competing with sgRNA loading poses a challenge, given that Cas9 binds to sgRNA with a high affinity. The sgRNA binding to Cas9 was previously measured as KD of 2–20 nM, which was much stronger compared with KD ∼200 nM of AcrIIC2 binding to Cas9.8,9 It has been reported that a 10-fold molar excess of AcrIIC2 was required to disable the Cas9–sgRNA complex formation in the SEC, where the Cas9–sgRNA binding was reduced by ∼17-fold. 9 In addition, mixing AcrIIC2 and sgRNA simultaneously with Cas9 required ∼20-fold higher concentration of AcrIIC2, compared with AcrIIC2 mixed with Cas9 before sgRNA to achieve similar Cas9 inhibition. 9 Thus, successful Cas9 inhibition would require AcrIIC2 to be expressed in large quantities to outcompete the guide RNA loading onto Cas9. Self-association of AcrIIC2 may bring further advantages, given that oligomers can act as multidentate ligands to enhance the functional binding affinity to Cas9. Protein multimerization is a well-known mechanism in the regulation of signaling proteins that cluster to overcome the low affinity of individual subunits. 28 The low affinity of the signaling proteins supposedly prevents unwanted downstream signaling in response to transient fluctuations in cellular ligand concentrations. It may seem intriguing, however, that phages evolved weak Acr proteins to counter their host immune system. We speculate that the weak Acr activity may benefit phages in terms of population dynamics during host infection. It has recently been reported that weak Acr proteins provide a better opportunity for Acr-carrying phages to compete with Acr-deficient phages for propagation in CRISPR-expressing host cells. 29 Weak Acrs select for Acr-carrying phages during cumulative host infections, whereas strong Acrs benefit Acr-deficient phages as well, serving as a common good of phage community. The previous study demonstrated advantages of weak Acr against the class 1 CRISPR-Cas system, and AcrIIC2 might address a similar advantage against the class 2 CRISPR-Cas system.
We employed engineered monodisperse AcrIIC2 to demonstrate that AcrIIC2 binding induces a helix in the BH peptide. BH is highly basic with conserved arginine residues, and plays an important role in sgRNA binding. 30 One may be curious whether AcrIIC2 binding could also induce a helical conformation in full-length Cas9. To answer this question, one needs to compare the N. meningitidis Cas9 structure in the apo state and in complex with AcrIIC2, both of which are currently unavailable. We instead try to address this question through indirect evidence and inference. First, BH forms a long helix (residues 56–91) in full-length N. meningitidis Cas9 bound to sgRNA. 25 The apo-state structure of type II-C Cas9 is available only from Cas9 of Actinomyces naeslundii, where the overall BH region lacks electron density maps, possibly due to a static or dynamic disorder. 31 This observation leaves a possibility that the BH region forms a stable helix only in complex with sgRNA or AcrIIC2. Second, the BH peptide of Francisella novicida Cas9 forms an elongated helix upon AcrIIC2 binding, such that the helix extends to the nonhelical region (residues 44–52) of Cas9–sgRNA.8,32 This observation suggests that AcrIIC2 binding may promote the helix longer than that in Cas9–sgRNA. The AcrIIC2-induced conformational changes in full-length Cas9 is still speculative, and warrants the structure determination of intact Cas9 and AcrIIC2. We finally wish to briefly remark on the BH peptide as a potential antagonist of AcrIIC2. Given the small size of the BH peptide, it may be an effective and economic way that host bacteria adopt the BH motif as anti-Acr to titrate out AcrIIC2 to quench the inhibition.
In conclusion, we demonstrate that AcrIIC2 self-associates in solution to maintain an ensemble of dynamic oligomers. The oligomerization relies on electrostatic interactions between complementary surface charges that are placed orthogonal to the Cas9 binding interface. In addition, the interaction between AcrIIC2 and the BH peptide of Cas9 reveals a conformational change of BH, which is unique among known Acr–Cas interactions.
Footnotes
Acknowledgments
We thank the Pohang Accelerator Laboratory for the support with data collection, and the high-field NMR facility at the Korea Basic Science Institute, and the National Center for Inter-University Research Facilities of Seoul National University.
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This study was supported by the Creative-Pioneering Researchers Program through Seoul National University (500-20200255) and the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT; 2020R1A4A1018890).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.