
Abstract
Brassica napus is the most important oil crop plant for edible oil and renewable energy source worldwide. Yield loss caused by pod shattering is a main problem during B. napus harvest. In this study, six BnSHP1 and two BnSHP2 homoeologs were targeted by the CRISPR (clustered regularly interspaced short palindromic repeats)-Cas9 (CRISPR-associated protein 9) genome editing system and multiple SHP1 and SHP2 mutated lines were identified for evaluating the contribution for pod-shattering resistance. Our data suggest that BnSHP1A09 is probably a promising homoeolog for controlling lignin contents at dehiscence zone. Simultaneous mutation of BnSHP1A09/C04-B/A04 and BnSHP2A05/C04-A exhibited reduced lignified layer and separation layer adjacent to valves and replum. The pod-shattering resistance index (SRI) subsequently increased to 0.31 in five homoeolog mutation lines compared with the wild type (SRI = 0.036), which provide the theoretical basis for breeding of commercial pod-shattering resistance variety.
Introduction
Brassica napus is an oilseed crop and widely grown in Canada, China, India, and the European Union. 1 China alone contributes 20% of the total world production. Pod shattering is a major problem in B. napus worldwide. When the fruit of B. napus ripens, shattering occurs early in the abscission zone due to the deposition of lignin content. Previous reports have shown that fruit dehiscence accounts for about 20% of yield loss of the total production during harvest and can reach up to 50% under adverse climatic conditions. 2
Oilseed rape and model plant Arabidopsis thaliana (Arabidopsis) belong to Brassicaceae and share similar morphological features in a pod structure, which is a benefit for investigating the genetic mechanism of seed dehiscence.3,4 Mature silique of B. napus and Arabidopsis comprise two valves, a replum and valve margins. The two valves are fused by a valve margin and replum, which comprised a separation layer (SL) and a lignified layer (LL). 5 The regulation of pod dehiscence at the mature stage is mediated by a complex genetic control pathway. The inflorescence morphogenesis is well established by a five-class ABCDE model. Among them, the D-Class is related to the SHATTERPROOF1/2 (SHP1/2) genes, which belong to the MADS-box gene family and are considered the primary regulator of dry fruit dehiscence. 6
In the single Atshp1 or Atshp2 mutant, valve margin identity and cell differentiation usually persist as in the wild type, whereas in the double Atshp1/2 mutant, valve margin did not develop, and the silique became indehiscent. 7 SHP proteins regulate the expression of the basic helix-loop-helix gene family, such as ALCATRAZ (ALC), INDEHISCENT (IND), and SPATULA (SPT). 8 SHP1/2 and IND are involved in the differentiation of both LL and SL, whereas ALC and SPT are only involved in the formation of SL.9,10 Expression of both SHP1 and SHP2 genes causes pod dehiscence by promoting cell differentiation and lignification in the strips of the valve margins. 11
One approach for generating pod-shattering-resistant material in oilseed rape is to downregulate the expression of SHP1/2 genes and ultimately activate the expression of FUL genes, which positively regulate pod-shattering resistance.12,13 Clustered regularly interspaced short palindromic repeats (CRISPR)-CRISPR-associated protein 9 (Cas9) genome editing system can precisely introduce double-stranded breaks within the targeted sequences and has been improved to be very efficient for eliminating undesirable agronomic traits in B. napus.14,15
Bnalc mutants generated by CRISPR-Cas9 lacked SL and exhibited partial resistance to pod shattering. Single-copy mutant of BnIND.A03 (aaCC) showed partial resistance, but the double mutant of both BnIND.A03 and BnIND.C03 (aacc) homoeologs exhibited complete pod-shattering resistance. 16 Knockout mutagenesis in all five homoeologs of JAGGED gene generated a callus-like cylindrical pod structure with severe defect in seed development, whereas mutation of BnJAG.A08 homoeolog leads to increased pod-shattering resistance. 17
The majority of oilseed rape genes have more than two homoeologs, which possess nearly identical genomic sequences between different copies located in subgenomes of polyploid species. 18 In contrast, homologues usually refer to gene or chromosome descent from a common ancestor. Different homoeologous gene copies of allopolyploid B. napus always exhibited redundant or diverse regulatory roles, which is a critical problem for genetic improvement in polyploid crops.
In this study, CRISPR-Cas9 system was successfully used to characterize the key functional homoeologs of BnSHP1/BnSHP2 genes in improving pod-shattering resistance. Cross-sectional and lignin staining results suggested that BnSHP1A09 homoeolog is crucial for regulating lignin composition at dehiscence zone. By combining mutagenesis at five homoeologs (BnSHP1A09, BnSHP1A04, BnSHP1C04-A, BnSHP2C04-B, and BnSHP2A05), a shattering resistant line was generated with reduced LL and SL at the DZ. These results would benefit genetic improvement of pod-shattering resistance trait in B. napus.
Materials and Methods
Selection of target sequences and vector construction
The oilseed rape SHATTERPROOF (SHP1) and (SHP2) genome sequences were downloaded from the B. napus genome resource website by BLAST searching analysis. Eventually, eight SHP homoeologs (BnaA04g01810D, BnaC04g23360D, BnaC04g52620D, BnaA05g02990D, BnaC08g29530D, BnaA09g55330D, BnaC09g10650D, and BnaC06g27260D) were selected for designing sgRNAs. Two different sgRNA expression cassettes were constructed, and each cassette carried two sgRNAs with 20 bp target sequences. The targeted exon region was selected upstream of the MADS-box domain neighboring 5′-NGG protospacer adjacent motif. CRISPR-P 2.0 online software was used for the design of highly efficient sgRNA with minimal off-target effects. 19
The oligos used in constructing sgRNA vectors are listed in Supplementary Table S1. The first cassette was designed for targeting four SHP homoeologs (BnaC06g27260D, BnaC08g29530D, BnaC09g10650D, and BnaA09g55330D), whereas the second cassette was designed for targeting the other four SHP homoeologs (BnaA04g01810D, BnaC04g23360D, BnaC04g52620D, and BnaA05g02990D).
Plant material and genetic transformation
The semiwinter-type accession of B. napus, Zhongshuang 6 (ZS6), was used for transformation, and seeds were obtained from the germplasm resource of Oil Crops Research Institute, Chinese Academy of Agricultural Sciences (OCRI-CAAS), Wuhan, China. Mutants generated from wild-type callus plants were grown in the greenhouse under normal conditions (16/8 h of light/dark at 22°C). Agrobacterium tumefaciens strain (GV3101) was used for plant transformation as described by Li et al. 20 Self-pollinated T0 seeds were harvested from the greenhouse and sown in the field of the experimental station of OCRI-CAAS in Hanchuan, China. T1 seeds from homozygous mutants were harvested and grew in the greenhouse to obtain T2 seeds for the next-generation characterization.
DNA extraction and mutant identification
Genomic DNA was isolated from leaf samples by a standard cetyltrimethylammonium bromide method. The positive transgenic plants were identified by polymerase chain reaction (PCR) amplification of the NPTII gene (Supplementary Table S2). The genomic region of the respective gene was amplified using specific primers to ensure gene size (Supplementary Table S3), and then, the PCR product was used for the second round of PCR using another set of primers to obtain product size of 150–350 bp. The annealed PCR product (150–350 bp) was analyzed by 10% nondenaturing polyacrylamide gel electrophoresis (PAGE), as described in a previous report by Li et al 20 (oligo's detail in Supplementary Table S4).
RNA isolation and expression analysis
Total RNA was extracted with the RNAprep Pure Plant Kit (Tiangen, Beijing). The plant materials used for RNA extraction were leaf, stem, developing flower bud, sepals, petals, stamen, ovules, pericarp, and different developmental stages of flower gynoecium, including 13, 14, 15, 16, 17, and 18 days after flowering.
RNA concentration was quantified by a NanoDrop 2000C spectrophotometer (Thermo Fisher Scientific). For cDNA synthesis, 1 μg RNA for each sample was treated with DNase-I to eradicate the genomic DNA and then used as a template for reverse transcription (QuantiTect Reverse Transcription Kit; Qiagen). The expression level of the actin gene was used to standardize the RNA sample for each reverse transcription-polymerase chain reaction. Quantitative real-time PCR was performed with SYBER Green Master Mix (Novogene) using a light cycler 480 real-time PCR system (Applied Biosystem). For each sample, three biological replicates were analyzed with three technical replicates and actin as an internal control.
Pod transverse section preparation
Pod lignin content was detected at stage 17-B right before the pod turning yellow. Fully elongated B. napus pods were harvested, halved, and immersed in 50% formalin aceto-alcohol solution immediately. Sample dehydration was done by passing through a series of dehydrating solutions (tertiary butyl alcohol [TBA], 95% ethanol, absolute ethanol, and distilled water). Dehydrated samples were transferred in tubes, half-filled with TBA and molten Paraplast (paraffin wax) for better wax penetration in tissues. After proper orientation of the tissue, samples were quickly cooled in a refrigerator at 4°C. Trimming of wax blocks was performed with a cutter in such a way that all the edges of the block were parallel with plane surfaces.
These wax blocks containing embedded tissue are then mounted on square wooden blocks for microtomy. Transverse sections of 8–10 μm thickness were straddled on a slide using a water bath before dewaxing in Histo-Clear. Cross sections were stained by 0.1% toluidine blue and 0.5% safranin-O for 30 min. For lignin-specific staining, pod cross sections were placed in 1% phloroglucinol for 2 min and subsequently acidified with 37% HCL drops. Images of pod cross section were visualized by an Olympus IX71 inverted microscope.
Random impact test
Random impact test (RIT) was followed with some modifications 21 for a laboratory test of pod-shattering resistance. Sixty pods were taken from the mutants with the same genotype on SHP homoeologs, and then bulked to make a composite sample for measuring shattering resistance index (SRI) by dividing them into three replications. Pods were dried at room temperature and placed in an incubator at 45°C for 7 h right before measurement. Twenty dry pods were placed in a container (14 cm diameter and 7 cm length) with eight steel balls of 14 mm diameter to shake for 2, 4, 6, 8, and 10 min at 240 RPM. Shattered pods were recorded after each shaking, and SRI was calculated using SRI = 1 − Σ5 i=1 x i (6 − i)/100.
Statistical analysis
The statistical significance of the differences among the means was analyzed using one-way analysis of variance (ANOVA) followed by Duncan's multiple range test in IBM, SPSS statistics (SPSS, Inc., IBM, New York). The differences at p ≤ 0.05 level were considered significant. 22
Results
Sequence analysis of BnSHP genes and vector construction
Bioinformatic analysis identified eight rapeseed SHP genes in B. napus genome. Phylogenetic tree analysis suggested that BnaA05g02990D and BnaC04g52620D, named BnSHP2A05 and BnSHP2C04-A, respectively, have the closest relationship with AtSHP2, whereas BnaA09g55330D (BnSHP1A09), BnaC08g29530D (BnSHP1C08), BnaC09g10650D (BnSHP1C09), BnaC06g27260D (BnSHP1C06), BnaC04g23360D (BnSHP1C04-B), and BnaA04g01810D (BnSHP1A04) showed a common lineage with AtSHP1. Among these SHP1 homoeologs, BnSHP1A04 and BnSHP1C04-B shared a common event of speciation (Fig. 1).

Phylogenetic tree of rapeseed BnSHP homoeologs with Arabidopsis SHP1 and SHP2 gene sequences. SHP homoeologs and their sequence similarity with Arabidopsis thaliana are expressed in a phylogenetic tree. As per evolutionary history and lineage differences among the BnSHP homoeologs, it seems that BnSHP2C04-A and BnSHP2A05 is close to the Atshp2 sequence. However, other BnSHP homoeologs showed their phylogeny with Atshp1 gene sequence. BnSHP1A04 and BnSHP1C04-B exhibited a relationship with each other and BnSHP1C06, BnSHP1C08, BnSHP1C09, and BnSHP1A09 have shown close phylogenetic lineages. Color images are available online.
Based on the phylogenetic tree, two expression vectors were constructed for targeting the conserved region of four SHP homoeologs (Fig. 2). In the first expression cassette, the fourth exon of BnSHP1C06, BnSHP1C08, BnSHP1C09, and the eighth exon of BnSHP1A09 are identical and were targeted by one sgRNA. The second expression cassette was designed with two sgRNAs, one sgRNA targeted to the second exon of BnSHP2C04-A and the third exon of BnSHP2A05. And the second sgRNA targeted to the eighth exon of BnSHP1C04-B and BnSHP1A04 (Fig. 2).
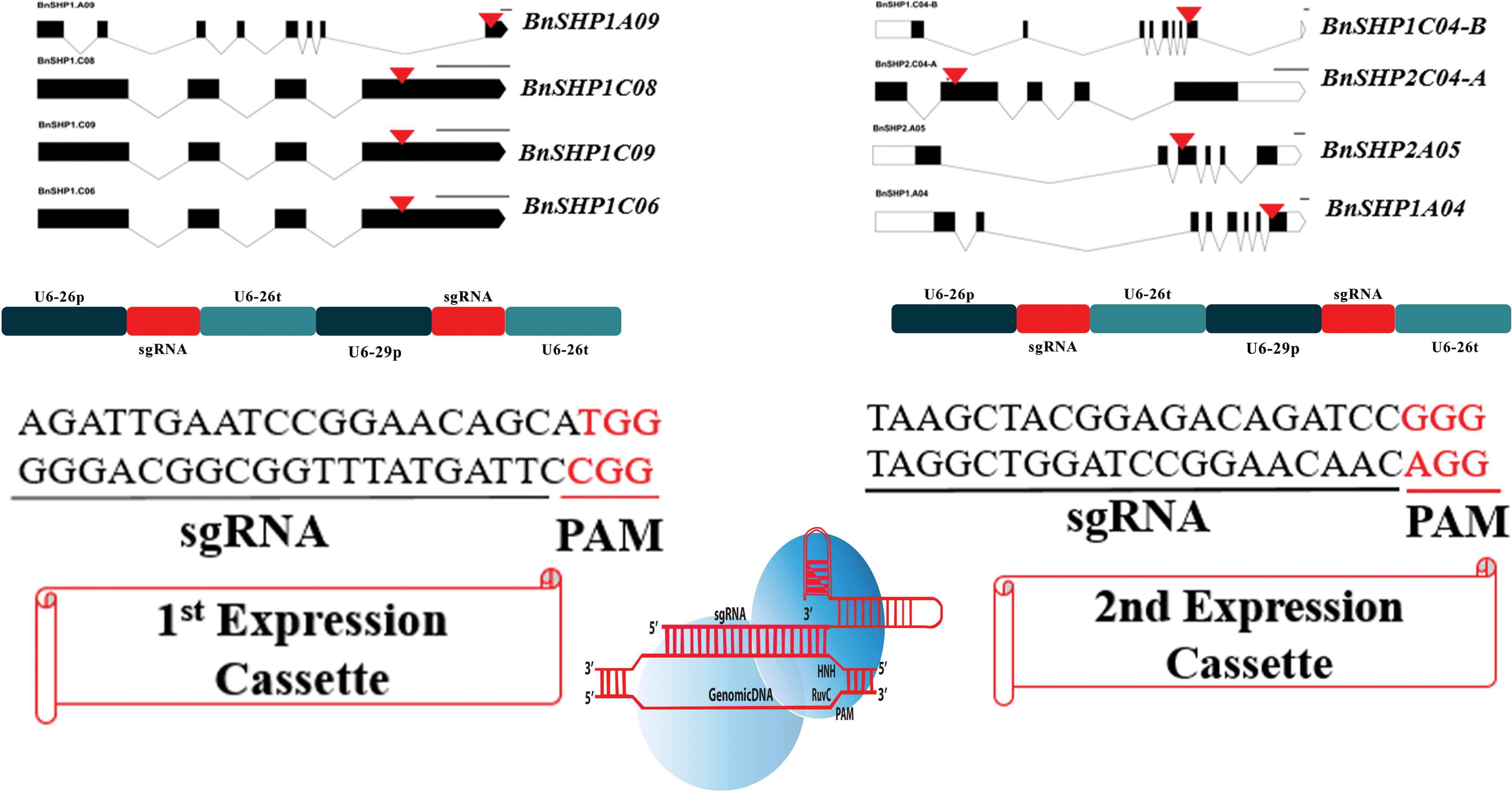
Construction of vectors for targeting oilseed rape SHATTERPROOF genes. Mutation induction was carried out with the CRISPR-Cas9 system in SHP homoeologs by constructing two vectors. The black line represents introns and the black boxes represent the exon regions. sgRNA targets were identified on the coding regions of the genes as mentioned with the red arrow on the exon. Vectors were constructed by using two sgRNAs as U6-26p-1st-sgRNA-U6-26t-U6-29p-2nd-sgRNA-U6-26t. CRISPR-Cas9, clustered regularly interspaced short palindromic repeats–CRISPR-associated protein 9. Color images are available online.
The specificity of these sgRNAs was confirmed by blasting rapeseed genome database (www.genoscope.cns.fr/blat-server/cgi-bin/colza/webBlat) (Supplementary Figs. S1 and S2) and CRISPR-P 2.0 website. 19 Both websites showed same on-target sites. In addition, all predicted off-target sequences were carrying less than 0.500 off-score, which means that our selected guide RNAs have no chance to induce off-targets.
The expression pattern of BnSHP homoeologs was carried out and the data suggested that BnSHP2C04-A, BnSHP1C04-B, BnSHP1A04, BnSHP2A05, and BnSHP1A09 homoeologs showed a significantly higher expression in the developing bud of flowers, ovules, pericarp, and a stably exhibited expression in the pod developmental phases from stage 13 to stage 18 (Supplementary Fig. S3). However, three homoeologs (BnSHP1C06, BnSHP1C08, and BnSHP1C09) exhibited very less genetic expression among stages 13 to 18.
Selection of genome editing mutation and evaluation of off-target effect
After genetic transformation, 270 and 247 plants were generated from the first and the second sgRNA cassettes, respectively. Among these regenerated plants, 131/270 and 80/247 kanamycin-positive plants were identified by PCR analysis (Supplementary Figs. S4 and S5). To identify the CRISPR-Cas9-induced mutations, all kanamycin-positive plants were screened by PAGE-based method. Based on PAGE screening and Sanger sequencing, 46 mutants generated from the first expression cassette (1SHP) and 22 mutants generated from the second expression cassette (2SHP) were obtained from those transgenic-positive plants.
Sequencing results suggested that insertion and deletion (Indels) events occurred in all mutants, ranging from 1 to 17 bp, including single-copy mutants (BnSHP1A09, BnSHP2C04-A, BnSHP1A04, BnSHP2A05, BnSHP1C06, BnSHP1C08), double-copy mutants (BnSHP1A09+BnSHP1C06, BnSHP1A04+BnSHP2C04-A, BnSHP1C06+BnSHP1C08), triple-copy mutants (BnSHP1C06+BnSHP1C08+BnSHP1A09), and five plants with quadruple-copy (BnSHP1A09+BnSHP1C06+BnSHP1C08+BnSHP1C09) (BnSHP1A04+BnSHP2C04-A+BnSHP1C04-B+BnSHP2A05) mutants (Supplementary Tables S5.1 and S5.2).
Among all the Indel homozygous T0 mutants, 1SHP 4 -17-11 exhibited mutations at BnSHP1A09, BnSHP1C06, BnSHP1C08, and BnSHP1C09 homoeologs (Fig. 3A). 1SHP 3 -17-3 showed mutagenesis at BnSHP1C06, BnSHP1C08, and BnSHP1C09, whereas the 2SHP 4 -484 line was mutated at four genomic positions on BnSHP2A05, BnSHP1A04, BnSHP2C04-A, and BnSHP1C04-B (Fig. 3B, C). 1SHP 1 -157 showed mutagenesis at BnSHP1A09 homoeolog (Fig. 3D). All these mutants were selected and forwarded to the next generation for further characterization (Supplementary Tables S6.1 and S6.2). All genetic changes in different homoeologs of the mutants were analyzed and protein sequences were predicted from the cDNA sequences. Indels induced in BnSHP homoeologs entirely changed the protein structure (Supplementary Figs. S6–S10).

Identification and characterization of CRISPR-Cas9-induced BnSHP mutant.
To evaluate off-target sites, the potential off-target sites located in BnaC03g72160D and BnaA09g55330D genes were analyzed by Sanger sequencing. We did not find any off-target mutagenesis at both genomic sites in all mutants (Supplementary Fig. S11).
Segregation pattern and inheritance of BnSHP mutations
All 46-T0 mutants generated from the first SHP expression cassette and 22-T0 mutants generated from the second expression cassette were self-pollinated to obtain T1 lines. Twelve T1 plants from each T0 mutant were examined to obtain a homozygous plant with mutations at all targeted genomic positions. After examining 700 T1-mutants, we selected homozygous transgenic plants derived from the first expression cassette and second expression cassette by Sanger sequencing (Supplementary Tables S6.1 and S6.2). The genotype of the mutants generated by crossing 1SHP 1 -157 and 2SHP 4 -484 was also confirmed.
These mutants were selected on genotypic and phenotypic basis by staining pod cross sections. In the F1 population, five mutants were identified in which all five homoeologs were mutated (Supplementary Table S7). After the sowing of F1 seeds, the selfing was carried out to obtain F2 seeds. Among the F2 population of BnSHP 5 -184-14, only 4 mutants (Supplementary Table S8) were identified from 2471 F2 individuals, which comprised Indel mutations at 5 SHP homoeologs.
BnSHP homoeologs display functional diversity in the dehiscence zone development
We examined the LL and SL pattern in SHP mutants by staining the transverse section of the pod at the 17-B stage with safranin-O and toluidine blue (Fig. 3 and Supplementary Fig. S12). In wild type, the lignin spread along the DZ, with four to five wide cells continued to the endocarp-b layer (Fig. 3E). In 1SHP 3 -17-3 containing mutagenesis at BnSHP1C06, BnSHP1C08, and BnSHP1C09, there was no change in SL and LL of DZ, but reduced replum was observed (Fig. 3F). However, lines with mutations at all four homoeologs (BnSHP1A09, BnSHP1C06, BnSHP1C08, and BnSHP1C09) or only at BnSHP1A09 homoeolog exhibited the same phenotype at the dehiscence zone, with decreased lignin deposition and discontinuous LL along the valve margins compared with WT (Fig. 3G, I). Cells in the LL spread discontinuously along the length of the valve margins toward the endocarp-b layer. Lignin deposition was not widened to three to four layers of cells as in WT.
These mutants showed a partial indehiscent phenotype due to a clear cut differentiation of SL layer at DZ. However, the 2SHP 4 -484 mutant line showed a large replum area with reduced SL development, but no difference in lignin deposition compared with WT (Fig. 3H). Our previous study suggested that increased replum width in the dehiscence zone might play a role in improving rapeseed pod-shattering resistance. 17 It was therefore hypothesized that BnSHP1C04-B, BnSHP1A04, BnSHP2C04-A, BnSHP2A05, and BnSHP1A09 may regulate the development of replum width. To test whether those five homoeologs have redundant roles, the 1SHP 1 -157 mutant line was crossed with the 2SHP 4 -484 mutant line to combine mutagenesis at five genomic positions in a single mutant named BnSHP 5 -184 for further analysis (Fig. 4).
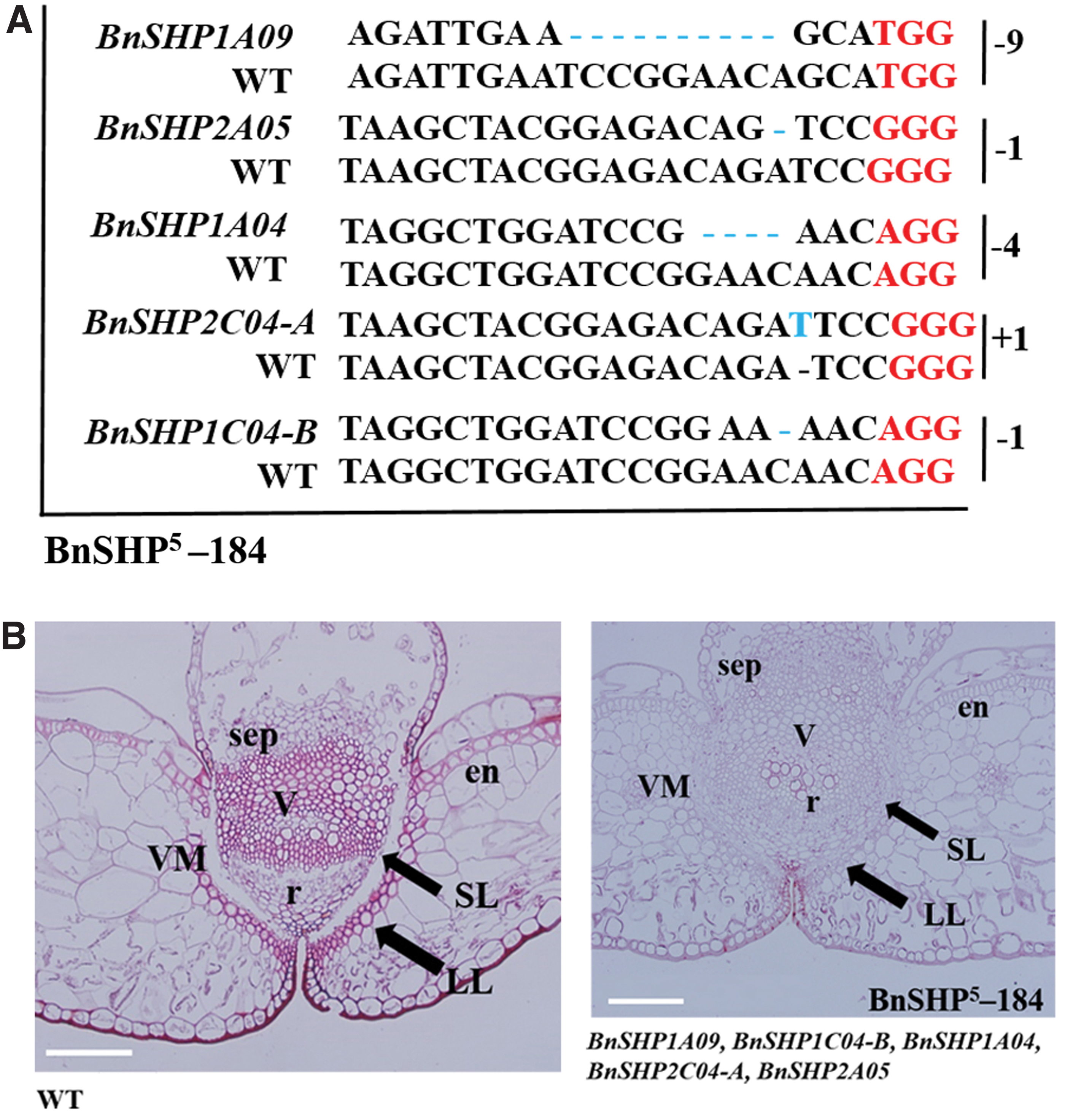
Genotypic and phenotypic comparison of wild-type and BnSHP
5
-184 mutants.
Patterning of dehiscence zone in five SHP homoeolog mutants
Simultaneously gathering mutagenesis in a single plant (BnSHP 5 -184) at five homoeologs of SHP gene (BnSHP1A09, BnSHP1C04-B, BnSHP1A04, BnSHP2A05, and BnSHP2C04-A) significantly alters the valve margin fate, especially reduced the lignin deposition and the differentiation of cells in the non-LL (Fig. 4). The number of cells and cell size were increased in the dehiscence zone, especially between the replum and valves. The four layers of cells that form the valve margin exhibited reduced lignification and did not develop valve margin throughout the pod. The lignin deposition was restricted to the base of the replum and extended discontinuously toward the endocarp-b layer.
Variation of pod-shattering resistance in SHP mutants
To further investigate the effect in pod SRI, all those SHP mutant lines were subjected for RIT evaluation. SRI varied significantly from wild type to mutants. RIT results suggested that the five SHP homoeolog mutated line BnSHP 5 -184 exhibited higher SRI (0.31) compared with the other genome editing mutants and WT (SRI = 0.036). The SRI of 2SHP 4 -484 mutants was increased to 0.109, whereas the triple mutants in which three SHP1 homoeologs (BnSHP1C06, BnSHP1C08, BnSHP1C09) were mutated did not show significant difference on SRI compared with WT (Fig. 5). In addition, the SRI of 1SHP 4 -17-11 mutant (BnSHP1A09, BnSHP1C06, BnSHP1C08, BnSHP1C09) and 1SHP 1 -157 (BnSHP1A09) mutant was increased to 0.157 and 0.173, respectively. Thus, these results demonstrated that BnSHP1A09 homoeolog probably played a dominant role in determining pod-shattering resistance. Moreover, the resistance to pod shattering can be further increased by gathering the BnSHP1A09 mutation site with BnSHP2A05, BnSHP1A04, BnSHP2C04-A, and BnSHP1C04-B.
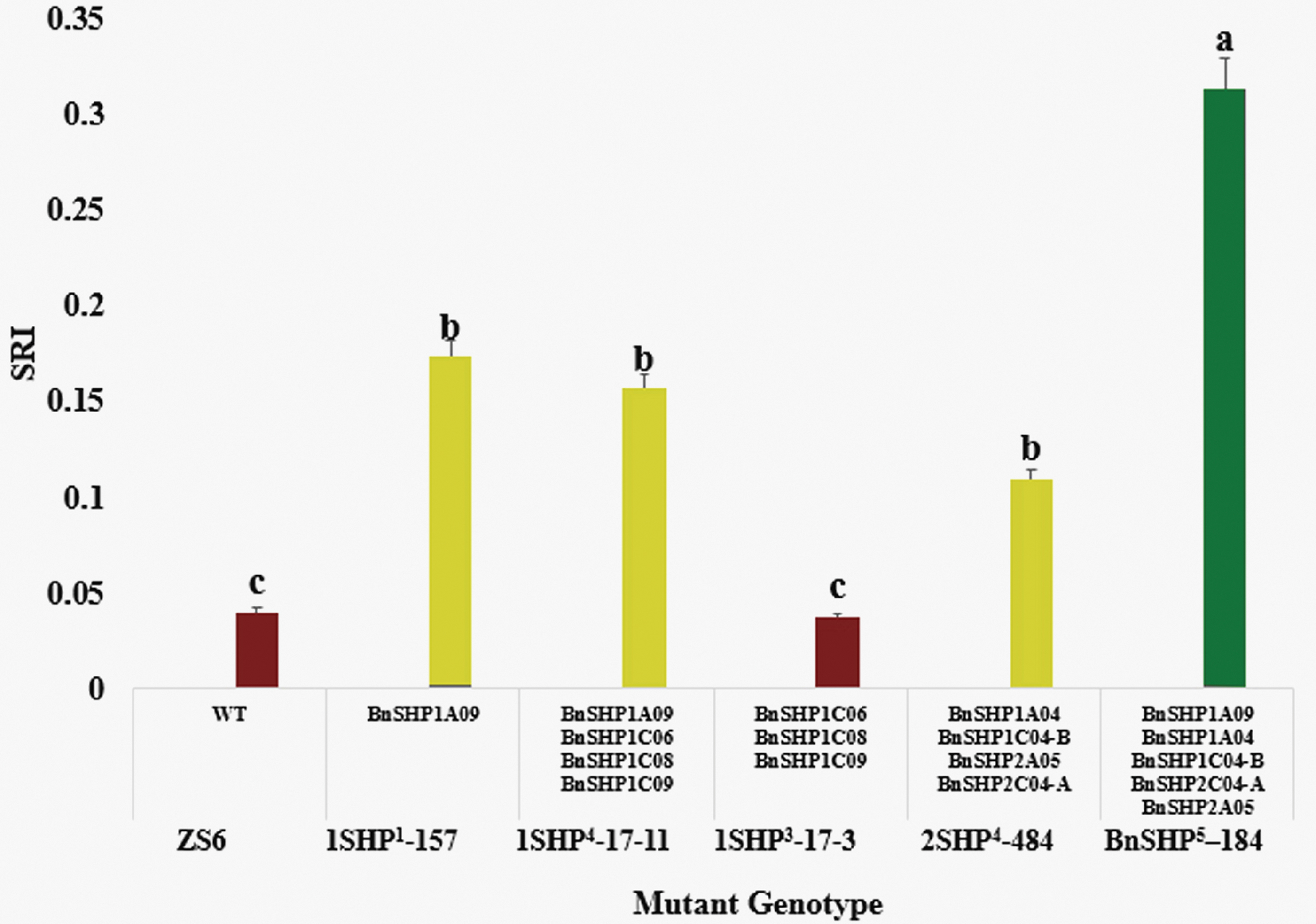
Pod-SRI of oilseed rape SHP mutants. Data were presented as mean of three biological replications ± SE (n = 3). The mean differences were analyzed using one-way ANOVA, followed by the DMRT. The differences were measured for significance at p ≤ 0.05 level. At the top of the bar, different letters show the significant difference at p ≤ 0.05 using DMRT. ANOVA, analysis of variance; DMRT, Duncan's multiple range test; SE, standard error; SRI, shattering resistance index. Color images are available online.
Identification of transgene-free mutants
To identify transgene-free mutants for potential breeding applications, vector-specific primers were used for PCR (Supplementary Table S2). In the T1 generation, several transgene-free lines were successfully identified, including 1SHP 4 -17-1, 1SHP 4 -17-6, SHP 5 -184, and 2SHP 4 -125. Among the descendants of 1SHP 4 -17 mutant, all 10 T2 generation mutants of 1SHP 4 -17-6 and 1SHP 4 -17-11 showed transgene-free genotype. Among the T2 mutants (1SHP 4 -17-1), 4/10 mutants were identified as transgene-free plants. Also, in the T2 generation of 2SHP 4 -484, one plant exhibited a transgene-free genotype. In the F2 population of BnSHP 5 -184, BnSHP 5 -184-14 was a transgene-free plant (Fig. 6).
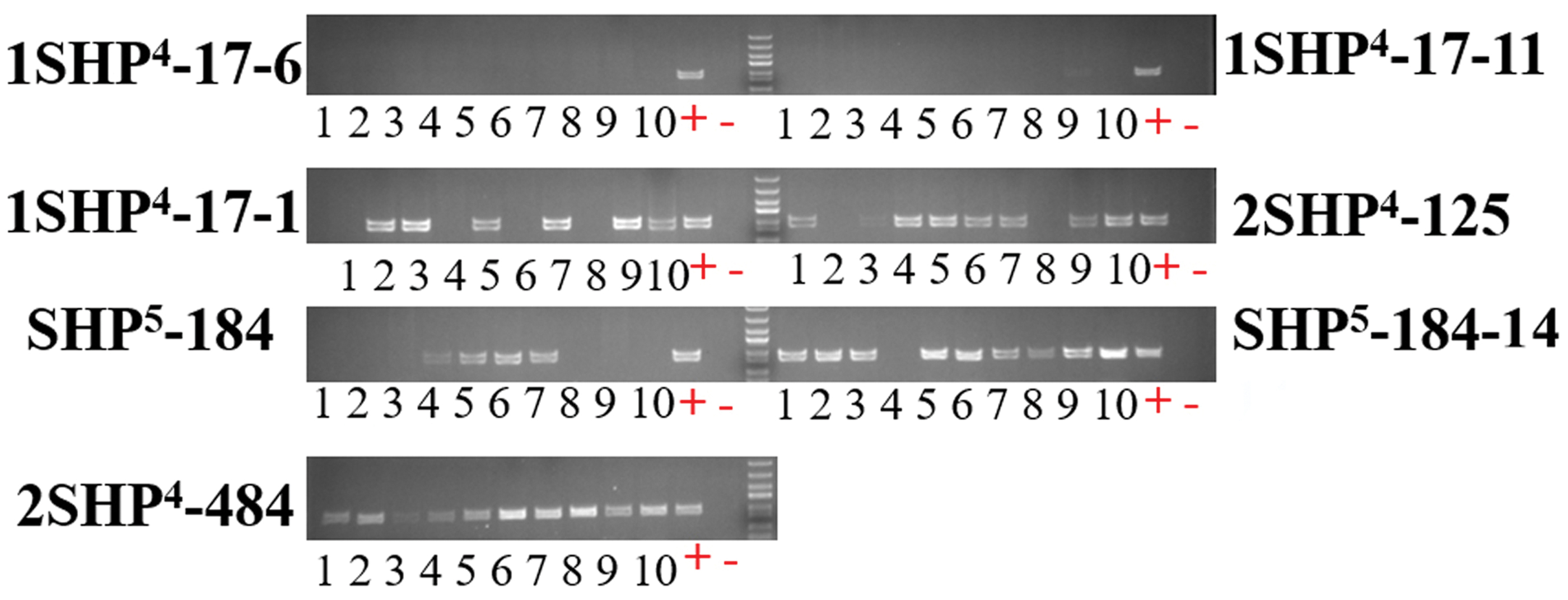
Identification of transgene-free mutants by using NPTII primers. + and − represent positive and negative controls. Ten plants were selected from the population, including one mutant as a positive check (+) and one ZS6-WT as a negative check (−). Those mutants that were not transgene-free showed clear bands of PCR product. However, some mutants were transgene-free, and did not exhibit any band. PCR, polymerase chain reaction; ZS6, Zhongshuang 6. Color images are available online.
Discussion
Preharvest pod shattering is a severe economic and agronomic issue resulting in loss of revenue in B. napus under normal climatic conditions. In harsh climatic conditions, it can reach up to 50% of loss and add volunteer plants in subsequent crops as well. 23 Due to the narrow range of genetic variation, it still lacks valuable germplasm for a commercial B. napus breeding program. 24
In this study, two BnSHP2 homoeologs and six BnSHP1 homoeologs were successfully knocked out by CRISPR-Cas9 to investigate the effects on pod-shattering resistance. By combining mutagenesis at five homoeologs (BnSHP1A09, BnSHP1A04, BnSHP2C04-A, BnSHP1C04-B, BnSHP2A05), a shattering resistant and transgene-free line was produced with reduced LL and SL at the DZ. Thus, it is possible to increase the pod SRI as shown in the mutated lines created by editing BnSHP genes in oilseed rape. The highest SRI reached 0.31, which is nearly 10 times higher than that of WT (0.036). Homozygous transgene-free mutations were obtained in T1 and F2 populations, facilitating the breeding process for commercial variety development using genome editing.
SHP genes belong to the MADS-box gene family. The MADS-box gene family has a diverse regulatory role in the development of root, leaf, and embryo.11,25 MADS-box genes also play roles in floral identity and were subdivided into A, B, C, D, and E classes, called the ABCDE model as well.26–29 These genes played different homeotic functions with various combinations, such as A class genes (APETALA1/2-AP1/2) exert specific function in sepals.30,31 Class-B gene (AP3) expresses in petals, whereas class-C genes (AGAMOUS-AG) express in carpels.32,33 Class-D genes (SHP1/2 and SEEDSTICK-STK) specify their role in ovary formation and ovule development. 34 Class-E (SEPALLATA1/2/3/4-SEP1/2/3/4) genes play a role in the formation of floral organs in four rounds. 27
Moreover, the class-D genes act redundantly at the top of the genetic regulatory cascade that directs the development of DZ.12,35 Bioinformatic analysis showed that there are six and two B. napus homoeologs of SHP1 and SHP2, respectively. Based on phenotypic investigation of the SHP mutations, we found reduced lignin formation at the DZ in a BnSHP1A09 mutated line (Fig. 3G). The same results were obtained in the line with four SHP homoeologs (BnSHP1A09, BnSHP1C06, BnSHP1C08, BnSHP1C09) (Fig. 3F) mutated simultaneously. The mutants carrying BnSHP1A09 mutations showed higher values of SRI probably due to reduced lignin deposition compared with WT (Fig. 5). Mutation of other SHP homoeologs (BnSHP2A05, BnSHP2C04-A, BnSHP1C04-B, BnSHP1A04) also produced pods with different SL and SRI.
However, the combination of five mutated homoeologs (BnSHP1A09, BnSHP2A05, BnSHP1A04, BnSHP2C04-A, BnSHP1C04-B) in a single mutant (BnSHP 5 -184) generated a significantly higher resistance to pod shattering (SRI = 0.31) than both WT (SRI = 0.036) and the two parental mutants. Reduced LL and SL in the five homoeolog mutants were observed along the valve margins in DZ (Fig. 4). These phenotypes suggest that neither mutation at BnSHP1C06, BnSHP1C08, BnSHP1C09, and BnSHP1A09 by first expression cassette nor mutation at BnSHP2C04-A, BnSHP2A05, BnSHP1C04-B, and BnSHP1A04 by second sgRNA cassettes is enough to significantly alter the valve margin cell fate adjacent to the DZ (LL and SL). Similar to the indehiscent phenotypes in Arabidopsis, BnSHP1 and BnSHP2 exhibited redundant regulatory function in DZ development. 8 Pyramiding mutations at more functional homoeologous sites have a significant effect on the enhancement of the targeted trait.
Phenotypic investigation data suggested that mutagenesis in BnSHP1C06/C08/C09 displayed fully developed LL and SL at DZ as WT (Fig. 3C, H). In addition, there is no significant difference between BnSHP1C06/C08/C09 mutant and WT in pod SRI. We speculate that BnSHP1C06, BnSHP1C08, and BnSHP1C09 homoeologs may not directly involve in pod-shattering resistance. These genes may indirectly interact with other genes to produce a typical flower structure. As revealed in the ABCDE model, D-class protein (Atshp1/2) interacts in a larger complex with the E-class proteins. E-class proteins (SEP1/2/3/4) showed redundant functions in the differentiation of sepals, petals, stamens, and carpels.27,36 This complicated interaction of three homoeologs of SHP1 with other proteins of ABCDE-model may have been developed in an evolutionary process of B. napus.
Conclusion
The CRISPR-Cas9 system is a robust tool to obtain functional knockout mutation in a gene with multiple homoeologs. Our results suggested that neither single mutant of SHP1 and SHP2 homoeologs displayed an indehiscent phenotype, but the BnSHP 5 -184 with five mutated homoeologs exhibited an obvious phenotypic change at DZ, and ultimately generated the shattering resistant phenotype. In addition, BnSHP1A09 was identified as a more promising locus for the lignin composition at DZ compared with other SHP1 homoeologs (BnSHP1A04, BnSHP1C04-B, BnSHP1C06, BnSHP1C08, and BnSHP1C09). We have generated stable transgene-free mutants on variable SHP genomic sites and produced polyploid B. napus plants with increased shattering resistance by targeting five homoeologs. Characterization of the BnSHP homoeologs undoubtedly provides new insights, which will benefit the rapid application of pod-shattering-resistant breeding programs in oilseed rape.
Footnotes
Acknowledgments
Project conceptualization: H.Q. and C.L. Literature mining and writing the original article: Q.U.Z. Review and editing: H.Q., C.L., B.J., Z.B., H.M., S.Y., C.W., and M.D. Funding acquisition: C.L., M.D., and H.Q. All authors read and approved the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 31700316 and No. U19A2029), the Science and Technology Innovation Project of Chinese Academy of Agricultural Sciences (Group No. 118), the Earmarked Fund for China Agriculture Research System (CARS-12), the Fundamental Research Funds for Central Non-Profit Scientific Institution (1610172018009), the Natural Science Foundation of Hubei Province (No. 2018CFB543), the Key Program of Technological Innovation in Hubei Province (No. 2016ABA084), and Graduate School of Chinese Academy of Agricultural Sciences.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.