
Abstract

Four reports capture structures of TniQ in association with a crRNA-guided surveillance complex.
Like so many “CRISPR stories,” this one starts with an observation by Eugene Koonin and collaborators. In 2017, Peters et al. identified variants of the type I CRISPR system (i.e., I-B and I-F) that lack adaptation machinery (cas1–2) and the nuclease (cas3) necessary for target degradation (Fig. 1). 1 It is not uncommon to find degenerate CRISPR systems. So, an observation such as this might not interest most of us, but Peters et al. doubled down. Using comparative genomics and phylogenetics, they showed that these minimal CRISPR-Cas variants associate with distinct families of Tn7-like transposable elements.
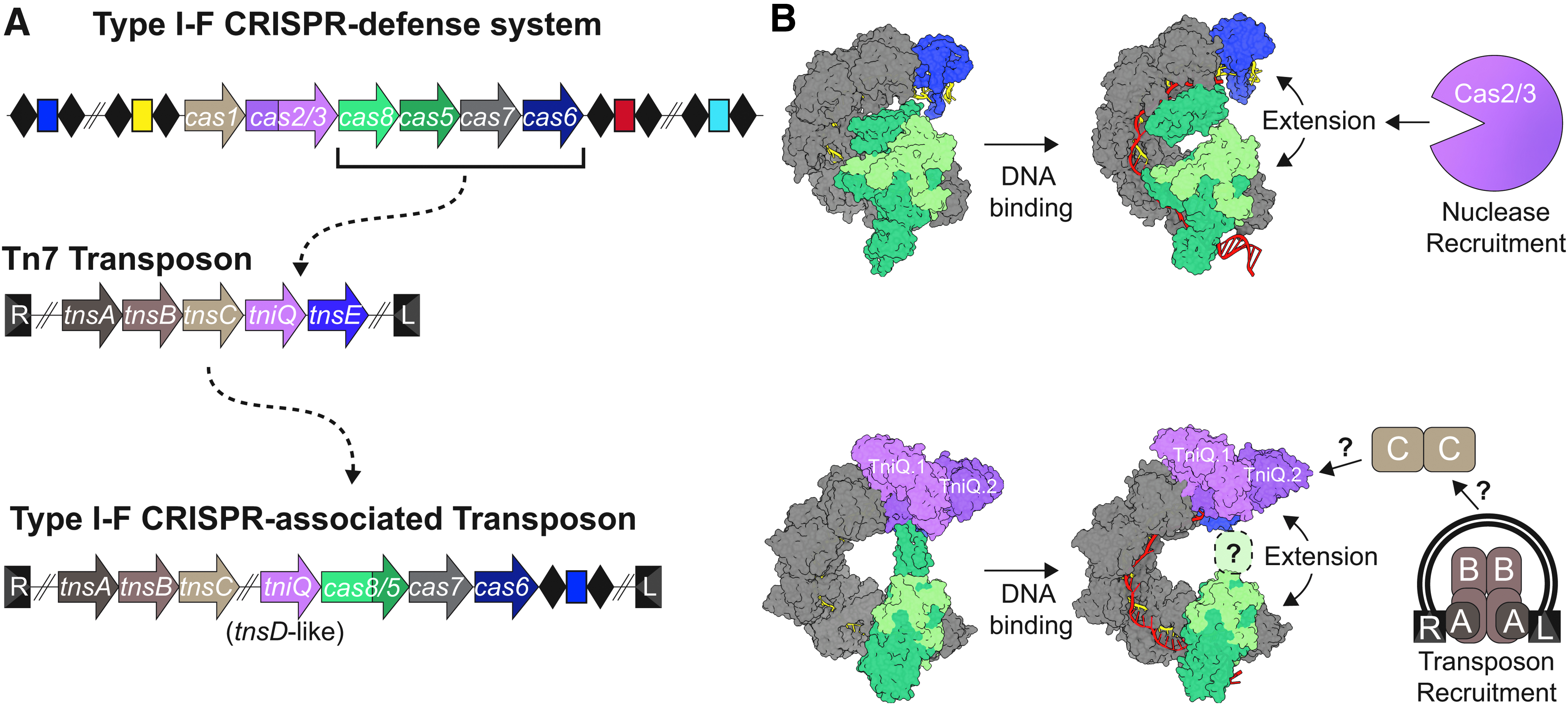
Comparison of the I-F CRISPR defense system to the I-F Tn7-like system.
Unlike most Tn7 transposons, which include tnsABC, tniQ, and tnsE, these particular Tn7-like elements lack the tnsE gene, which is responsible for transposition into plasmid DNA. In place of tnsE, these Tn7-like transposons encode cas genes that are characteristic of a type I CRISPR RNA (crRNA)-guided surveillance complex (Fig. 1). 2 Intriguingly, the spacers associated with the CRISPRs of these truncated systems most frequently match sequences derived from plasmids and other mobile genetic elements. This suggests that the crRNA-guided surveillance complex functionally complements TnsE, although it is not clear if or how this system provides an advantage over TnsE-mediated transposition. It is possible that crRNA-guided insertion of the transposon may inhibit replication of the incoming mobile genetic element, which could provide a defensive function for the host, but may limit the spread of the transposon.
In short, it is unclear whether the transposon domesticated the CRISPR, the CRISPR domesticated the transposon, or if the partnership is somehow mutually beneficial.
The Peters et al. paper concludes provocatively, noting that “transposon encoded CRISPR-Cas systems could be harnessed for genome-engineering applications.” This statement resonated with Samuel Sternberg's group. Working with a Tn7-like transposon encoding a variant I-F system, Sternberg's group showed last year that the crRNA-guided surveillance complex targets DNA transposition 47–51 bp downstream of the crRNA guide. 3 Showcasing its versatility, they engineered this system to deliver various genetic payloads to 16 different chromosomal locations with 95% or better on-target integration.
Hell Breaks Loose
Eric Sontheimer was recently quoted as saying, “Like everything else in the CRISPR world, the thin edge of the wedge comes in, and the next thing you know all hell breaks loose.” 4 Sontheimer's statement was in reference to the use of anti-CRISPRs for applications in genome engineering, but his words apply to most research in this field, including crRNA-guided transposons. In the past few months, four papers have been published on structures of transposon-encoded CRISPR-Cas systems. These include Sternberg's group in collaboration with Israel Fernández in Nature last December 5 (5 months after their preprint was posted on bioRxiv) and three other teams writing in Cell Research.6–8
Collectively, these papers report a total of 10 structures including crystal structures of TniQ, and cryo-EM structures of TniQ in association with the I-F crRNA-guided surveillance complex from Vibrio cholerae (i.e., VcCascade-TniQ), both before and after binding to a DNA target. The minimal I-F crRNA-guided surveillance complex maintains a crescent-shaped backbone, comprised of six Cas7 subunits, and is capped on either end by a Cas6 “head” and a Cas8/5 “tail” (Fig. 1B). This overall shape is similar to what has been observed in structures of I-F surveillance complexes involved in phage defense (Fig. 1B).9–13 Moreover, crRNA-guided DNA binding triggers an elongation in the backbone.10,12 In the I-F defense systems, elongation of the backbone coincides with a dramatic rearrangement of a helical bundle on Cas8. 12 This rearrangement locks the complex onto the DNA target and presents a landing pad for nuclease recruitment.
In the Tn7-like I-F CRISPR system, Cas8 and Cas5 are fused (Cas8/5) and the corresponding helical bundle is positioned like a bridge that connects the Cas6 head to the tail of the complex. Density for this region of the protein is weak, and the importance of this position remains unclear, but its location suggests that it may be involved in stabilizing the displaced strand of DNA (R-loop) and/or coordinated recruitment of transposon-associated proteins. Contacts between the helical bundle of Cas8/5 and TniQ, which forms a homodimer that is prominently positioned at the head of the complex, offer additional support for a role in coordinated recruitment of transposon-associated proteins (Fig. 1B).
While this work has revealed specific interactions between VcCasacade and TniQ, additional structures that include TnsABC will help clarify the mechanism of transposon recruitment. We anticipate that additional mechanistic insights will help develop these systems for creative applications in genome engineering. Once these and other transposon-associated CRISPR systems 14 are used to deliver a therapeutic payload to a specific genetic address in the human genome, then “all hell” is really going to break loose.
Footnotes
Acknowledgments
We thank Halpin-Healy et al., Li et al., and Jia et al. for providing atomic coordinates ahead of release by the PDB.
Author Disclosure Statement
B.W. is the founder of SurGene, LLC, and is an inventor on patent applications related to CRISPR-Cas systems and applications thereof.
Funding Information
Research in the Wiedenheft lab is supported by the National Institutes of Health (R35GM134867) and the Montana State University Agricultural Experimental Station (USDA NIFA).