
Abstract
Many therapeutic applications of CRISPR-Cas9 gene editing rely on delivery using the highly versatile adeno-associated virus (AAV) vector. The smallest type II Cas9 ortholog—Cje1Cas9, derived from Campylobacter jejuni with <1,000 amino acids—is particularly attractive for AAV delivery. However, the complex protospacer adjacent motif (PAM) of Cje1Cas9 (N3VRYAC) greatly restricts the density of recognition sequences in human genome. In this study, we identify two compact CjeCas9 orthologs designated as Cje2Cas9 and Cje3Cas9, whose PAM-interacting residues are different from those of the well-known Cje1Cas9. They can induce efficient genome editing in human cells, and their simpler trinucleotide PAM (N4CYA) requirements expand the scope of targeting. Moreover, Cje3Cas9 efficiently disrupts the Tyr gene in mice after being micro-injected into zygotes with the corresponding sgRNA. It also successfully disrupts the Pcsk9 gene in 8-week-old mouse liver after delivery with an sgRNA using an all-in-one AAV delivery vehicle. The gene-edited mice showed lower cholesterol level than wild-type mice. Notably, the 8e-nCje3–ABE and an sgRNA targeting Pcsk9 were successfully packaged into a single AAV vector for genome editing in adult mouse liver, with editing efficiency up to 12%. Thus, simple PAMs and a compact size enable Cje2/3Cas9 to expand the target scope of CRISPR-Cas9 toolsets, exhibiting considerable potential for therapeutic applications.
Introduction
The CRISPR-Cas9 system is a revolutionary genome editing platform that has been widely used in biotechnology and medicine.1–4 Streptococcus pyogenes Cas9 (SpCas9) plays a dominant role in genome manipulation because it exhibits a high editing efficiency and recognizes a relaxed 5′-NGG-3′ protospacer adjacent motif (PAM). SpCas9-based base editors (BEs), comprising deaminases and Cas9 nickase, enable targeted C-to-T (CBE) or A-to-G (ABE) conversions in genomic DNA and are also powerful genome editing tools for therapeutic applications.5–7 However, the SpCas9 (1,368 aa)–sgRNA complex exceeds the cargo capacity (∼4.7 kb) of a single adeno-associated virus (AAV) vector for efficient delivery in vivo.8,9
There are a few solutions to overcome this limitation. On one hand, SpCas9 and its sgRNA can be packed and delivered separately, 10 with the BEs split into two smaller parts through intein-mediated protein transsplicing, delivered by a pair of AAV vectors.11–15 On the other hand, there are several small Cas9 orthologs that result in permanent genome editing and positive therapeutic outcome in vivo by single AAV-mediated delivery, including SaCas9 (1,053 aa), 8 Nme1Cas9 (1,082 aa), 16 Nme2Cas9 (1,082 aa), 17 St1Cas9 (1,121 aa), 18 Cje1Cas9 (984 aa), 19 and so on (Supplementary Table S1).
We have also optimized a small SpaCas9 (1,130 aa) that enables efficient editing in adult mice liver using single AAV-mediated delivery. 20 As with other small Cas9-based BEs, SpaCas9-BEs cannot be packaged into a single AAV vector for in vivo genome modification.
The smallest Cje1Cas9 is <1,000 aa, making it particularly suitable for all-in-one AAV delivery; even its BE systems hold potential for all-in-one AAV delivery. However, as with most small Cas9s recognizing complex PAM sequences, Cje1Cas9 recognizes a very intricate N3VRYAC (V = A/C/G, R = A/G, Y = C/T) PAM, which greatly restricts its targeting scope.19,21,22 Therefore, it is important to explore novel small CRISPR-Cas enzymes with simple PAMs to expand the scope of targeting.
It has been reported that Cas9 PAM-interacting domains (PIDs) evolve under natural selective pressure. As a result, closely related orthologs have distinct PAM requirements. 17 In this study, we identified two compact Cje1Cas9 orthologs—Cje2Cas9 and Cje3Cas9—by comparing sequence and structural information of Cje1Cas9 with that of orthologs. We demonstrate that Cje2Cas9 and Cje3Cas9 function efficiently at endogenous sites with a simple N4CYA PAM. Importantly, the combination of Cje3Cas9–sgRNA/8e-nCje3Cas9–sgRNA complex and a single AAV-mediated delivery efficiently induces gene editing in adult mice liver, holding potential for therapeutic applications.
Materials and Methods
Animals
C57/BL6 mice and ICR mice were provided by the Laboratory Animal Center of Jilin University (Changchun, China). All animal studies conformed to regulations issued by the animal welfare and research ethics committee at Jilin University.
Plasmid construction
Cje1Cas9 was bought from Addgene (#89752) and the plasmid encoding pmTmG reporter was provided by Dr. Feng Gu. 23 Codon optimization and synthesis of Cje2Cas9 and Cje3Cas9 were conducted by Genscript Biotech (China). Mutations were introduced into plasmids using the Fast Site-Directed Mutagenesis Kit (Tiangen, China) and primers are listed in Supplementary Table S2. (Plasmids sequences are shown in the Supplementary Sequence.)
Cell culture and transfection
HEK293T and N2a cell lines were maintained at 37°C under 5% CO2 in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS) (HyClone, China), 100 U/mL penicillin, 100 mg/mL streptomycin, and GlutaMAX. The cells were seeded in 24-well plates on day 1 and transfected on day 2 at 80–90% confluency using Hieff Trans™ Liposomal Transfection Reagent (Yeasen, China). A total of 500 ng DNA was seeded into each well of a 24-well plate. Three micrograms per milliliter puromycin (Meilunbio, China) was used to enrich the positive transfected cells 24 h after transfection.
DNA isolation from cells
Seventy-two hours after transfection, cells were washed twice with phosphate-buffered saline (PBS) before lysed using One Step Mouse Genotyping Kit (Vazyme Biotech, China) at 55°C for 25 min, followed by enzyme inactivation at 95°C for 5 min. Cell genomic DNA was then used as templates for polymerase chain reaction (PCR) amplification. Amplicons were subjected to Sanger sequencing and analyzed by tracking of indels by decomposition (TIDE) 24 or EditR. 25 Sequences of all target sites and primers used to amplify target sites are given in Supplementary Tables S3 and S4.
PAM characterization using PAM Definition by Observable Sequence Excision
We performed the PAM Definition by Observable Sequence Excision (PAM-DOSE) assay as reported in a previous study. 23 In brief, HEK293T cells were cotransfected with plasmids encoding SpCas9/Cje1Cas9/Cje2Cas9/Cje3Cas9, corresponding gRNA expression plasmids, SaCas9 and its gRNA expression plasmids, and pmTmG library plasmids at the ratio of 1:1:1:1:1 using Hieff Trans Liposomal Transfection Reagent (Yeasen, China). Seventy-two hours after transfection, cell genomic DNA was extracted using TIANamp Genomic DNA Kit (Tiangen, China) and subjected to PCR amplification of sequences flanking the CRISPR target locus.
Amplicons were isolated by agarose gel electrophoresis before analyzed by deep sequencing. The paired-end deep sequencing of PCR amplicons was conducted by Sangon Biotech (Shanghai, China), using an Illumina MiSeq. PAM regions without indel within three bases near PAM were extracted. PAMs were counted and used to generate sequence logos and PAM wheels. 26
pmTmG reporter assay
Plasmids encoding Cje2Cas9/Cje3Cas9, gRNA, and pmTmG reporter were transfected into HEK293T cells at the ratio of 1:1:1 using Hieff Trans Liposomal Transfection Reagent (Yeasen, China). The fluorescent images of cells were captured using a microscope (ts100; Nikon, Tokyo, Japan), followed by editing quantification by flow cytometry. All editing frequencies were quantified as (number of green fluorescence-containing cells)/(number of fluorescence-containing cells).
mRNA and gRNA preparation
We linearized plasmids encoding Cje3Cas9, rA1-nCje3Cas9–CBE, and 8e-nCje3Cas9–ABE with NotI at 37°C for 12 h. The linearized plasmids were purified using TIANquick Midi Purification Kit (Tiangen Biotech, China) before being transcribed in vitro using the HiScribe™ T7 ARCA mRNA kit (NEB). The in vitro transcribed mRNAs were stored at −80°C after purification using the RNeasy Mini Kit (Qiagen). Aliquoting was necessary for convenience and longer storage. The sgRNA oligos were synthesized, annealed, and then inserted into the pUC57-Cje gRNA expression vector that contains a T7 promoter.
Sequences containing T7 promoter, target sites, and scaffold v2 were then amplified from pUC57-Cje gRNA expression vector and purified before transcribed in vitro using the MAXIscript T7 kit (Ambion). The in vitro transcribed sgRNAs were purified using the miRNeasy Mini Kit (Qiagen) according to the manufacturer's protocol.
Microinjection of mouse zygotes and genotyping
The cytoplasm of pronuclear-stage zygotes was injected with Cje3Cas9, rA1-nCje3Cas9–CBE, or 8e-nCje3Cas9–ABE mRNA (50 ng/μL) and sgRNA (30 ng/μL). Groups of ∼15 zygotes were injected with different Cje3 effectors and corresponding sgRNAs to identify gene editing efficiencies of the effectors. The zygotes were then cultured in potassium simplex optimized medium at 37°C with 5% CO2 and 100% humidity. The zygotes were collected individually when developed into blastocyst stage to analyze gene editing efficiencies.
After lysis with embryo lysis buffer (1% NP40) at 56°C for 60 min and then at 95°C for 10 min, they were subjected to PCR amplification. Amplicons containing target sequences were analyzed by Sanger sequencing. Newborn mice were toe clipped for genomic DNA extraction. Sequences containing target sites were amplified from genomic DNA and identified by Sanger sequencing. All primers used for genotyping are given in Supplementary Table S4.
Off-target assay
COSMID (https://crispr.bme.gatech.edu/) 27 was used to predict potential off-target sites for each sgRNA. For each sgRNA, we selected the top 5 potential off-target sites to test off-target editing. Off-target sites for all sgRNAs used in this study were amplified and analyzed by deep sequencing. Primers amplifying off-target sequences are given Supplementary Table S5.
AAV injection and processing
Construction of AAV–Cje3Cas9 and AAV–8e-nCje3Cas9 expression cassettes and viral production were conducted by Packgene Biotech (China). The titer of AAV–Cje3Cas9 was 2 × 1012 genome copies (GCs)/mL. The titer of AAV–8e-nCje3Cas9 was 2 × 1012 GCs/mL and 2 × 1013 GCs/mL. All dosages of AAV were adjusted to 200 μL with sterile PBS before the injection. Then, 2 × 1011 GCs AAV–Cje3Cas9 targeting Pcsk9-1 site was delivered to an 8-week-old female ICR mouse intravenously through lateral tail vein injection. For AAV–8e-nCje3Cas9 targeting Pcsk9-2 site, the titers used in each mouse were 2 × 1011 GCs and 4 × 1012 GCs, respectively.
All mice were maintained at animal facility with standard diet and housing. We fasted mice for 12 h before collecting blood from the mandibular vein at 0 and 30 days. The levels of serum total cholesterol, albumin (ALB), and alanine aminotransferase (ALT) were measured using a Catalyst One Chemistry Analyser (IDEXX) following the manufacturer's protocol. Mice were euthanized 30 days after vector administration, and liver tissues were collected for analysis by deep sequencing. Liver tissues were fixed in 4% formalin overnight, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (H&E).
Quantitative real-time PCR
Total RNA of edited mouse liver was isolated with TRNzol-A+ reagent (Tiangen, China) according to the manufacturer's instructions. Then, cDNA was synthesized using FastKing gDNA Dispelling RT SuperMix (Tiangen, China). Primers used for quantitative PCR (qPCR) are listed in Supplementary Table S4. The qPCR detection was carried out using SuperReal PreMix Plus (Tiangen, China) with Bio-Rad IQ5 multicolor real-time PCR detection system. The relative Cje3Cas9 expression was normalized to Gapdh.
Statistical analysis
All data are expressed as mean ± s.e.m. of at least three replicates for all experiments. Student's t-test through GraphPad prism software 8.0.1. was used to compare editing efficiencies of different Cas enzymes in HEK293T. The probability value <0.05 (p < 0.05) indicates statistical significance. *p < 0.05, **p < 0.01, ***p < 0.001.
Results
Two CjeCas9 orthologs with key mutations in PAM-interacting residues recognize different PAMs
Although CjeCas9 is the smallest of all effectors, it recognizes a complicated PAM (N3VRYAC), as do most small Cas9 nucleases such as Nme1Cas9 (N4GATT) and SaCas9 (NNGRRT). This greatly restricts the choice of targets available for editing. 19 In addition, three small orthologs characterized recently—Nme2Cas9, SauriCas9, and SlugCas9—require short PAM sequences (N4CC and NNGG), indicating that smaller Cas9s with simple PAMs can be discovered from natural orthologs.9,17,28 We also intended to explore new CjeCas9 homologs having distinct PAM requirements to further expand the targeting scope. The crystal structure of CjeCas9 indicates that key residues (T913 and S915) determine the fourth and fifth bases of the NNNVRYAC PAM specificity. 21
First, we searched for all related orthologs of Campylobacter jejuni Cas9 in NCBI identical protein groups (Fig. 1A). Then 96 Cas9s with divergent residues at 913 or 915 were selected for further analysis after protein sequence alignment with CjeCas9 from strain NCTC 1168, known previously as CjeCas9 but referred to hereafter as Cje1Cas9 (Fig. 1A and Supplementary Figs. S1 and S2). Interestingly, we noticed that a class of CjeCas9 orthologs harbor a T to D change at residue 913, similar to H to D change at key residue 1028 in NmeCas9 orthologs (Supplementary Figs. S2 and S3). 17 The D1028 residue of Nme2Cas9 has been demonstrated to determine the strong requirement for a C at the fifth base of the NNNNCC PAM. 29

Three closely related Campylobacter jejuni Cas9 orthologs have distinct PAMs.
Structural homology modeling shows that Cje1Cas9 is 32% identical with Nme2Cas9, and T913 residue in Cje1Cas9 corresponds to D1028 residue in Nme2Cas9 (Fig. 1B and Supplementary Fig. S4). Therefore, we speculated that the T to D change at residue 913 of Cje1Cas9 orthologs may convert the N4G PAM into an N4C PAM (Fig. 1B). Based on these considerations, we selected two CjeCas9 orthologs with D913 residue from C. jejuni strain PNUSAC007013 and PNUSAC009442 for further analysis, referred to as Cje2Cas9 (987 aa, 96% of sequence similarity to Cje1Cas9) and Cje3Cas9 (1000 aa, 60% of sequence similarity to Cje1Cas9), respectively (Fig. 1C and Supplementary Fig. S4).
The Cje2/3Cas9 locus contains an array composed of 36-bp direct repeats (DRs), which were interspaced by 30-bp spacers in the proximity of the Cas gene operon, similar to that of Cje1Cas9 locus (Fig. 1C).
Next, we utilized a positive screening system designated as PAM-DOSE to identify whether these CjeCas9 orthologs recognize distinct PAMs. 23 In brief, a pool of pmTmG plasmids consisting of a randomized 8-bp sequence (5′-NNNNNNNN-3′) and a fixed SaCas9 target site was generated. Only when Cas enzymes of interest (SpCas9, Cje1/2/3Cas9 in this study) recognize a functional PAM sequence and SaCas9 cleaves the fixed target site simultaneously can the CAG promoter drive the expression of the EGFP gene. 23
Therefore, the efficacy of Cas enzymes of interest is visible under the fluorescent microscope or through flow cytometry analysis (FCA), whereas identifying specific PAM requirements entails PCR amplification and deep sequencing analysis (Fig. 1D). As expected, the conventional SpCas9 and Cje1Cas9 recognized NGG and N3VRYAC as PAMs, respectively, consistent with previous studies (Fig. 1E and Supplementary Fig. S5). Notably, Cje2Cas9 (N4CYAT) and Cje3Cas9 (N4CYA) recognize the N4C PAM, confirming the previous speculation (Fig. 1E and Supplementary Fig. S5).
To examine the propensity for PAM recognition extending beyond position 8 (the length of randomization in our PAM library), the spacer targeting the PAM library was also shifted forward from 5′ by 2 nt. This allowed PAM identification to be extended to the 10th base. As a result, all three CjeCas9s showed no obvious preference at the 9th and the 10th positions (Supplementary Fig. S6). Overall, these results suggested that CjeCas9 orthologs with key mutations in PAM-interacting residues exhibited novel PAM specificities.
Determination of efficient PAMs required by Cje2/3Cas9-mediated gene editing
We quantitatively compared the functional PAM sequences of Cje2/3Cas9 by transfecting HEK293T cells with Cje2/3Cas9-encoding plasmids, corresponding gRNA and pmTmG reporter plasmids consisting of different PAM sequences based on an optimal GTACCTAT PAM (Fig. 2A). The editing efficiency of different PAMs was quantified by calculating the ratio of EGFP-positive cells to fluorescence-positive cells sorted by FCA (Fig. 2A). As a result, both Cje2Cas9 and Cje3Cas9 showed strong preference for a C at the fifth base and T/C at the sixth base of the PAM (Fig. 2B–D).

Detailed PAM sequence analysis of Cje2/3Cas9.
At the seventh base, Cje2Cas9 efficiently recognizes both T and A, whereas Cje3Cas9 only favors an A (Fig. 2B, C). Moreover, Cje2Cas9 exhibited moderate preference for a T at the eighth base and Cje3Cas9 was compatible with all bases (Fig. 2B, C). These quantitative comparison results were consistent with the observations of PAM identification from pmTmG library.
Taken together, these data demonstrated that Cje2Cas9 and Cje3Cas9 can efficiently cleave target sites containing N4CYWT (Y = C/T, W = A/T) and N4CYA PAMs in human cells, respectively. Compared with intricate N3VRYAC PAM of conventional Cje1Cas9, the trinucleotide PAM of Cje3Cas9 is more universal, expanding genome targeting range and significantly increasing the number of editable sites.
Improved efficiency of Cje2/3Cas9 with optimized sgRNA
It has been reported that Cje1Cas9 exhibited relatively low and variable activity in human cells, as most of type II-C Cas9 orthologs.19,21 Therefore, we attempted to improve editing activities of Cje2/3Cas9 by optimizing the gRNA. We noticed the DR and tracrRNA sequences of Cje2Cas9 are almost identical to those of Cje1Cas9 while different from those of the Cje3Cas9 (Fig. 3A). In addition, there is a poly Us in previously used gRNA scaffold v1 of Cje1Cas9, which could potentially inhibit transcription efficiency since it is the pause signal for RNA polymerase III (Fig. 3A).19,20 Thus, we engineered the gRNA scaffold attempting to enhance editing efficiencies.
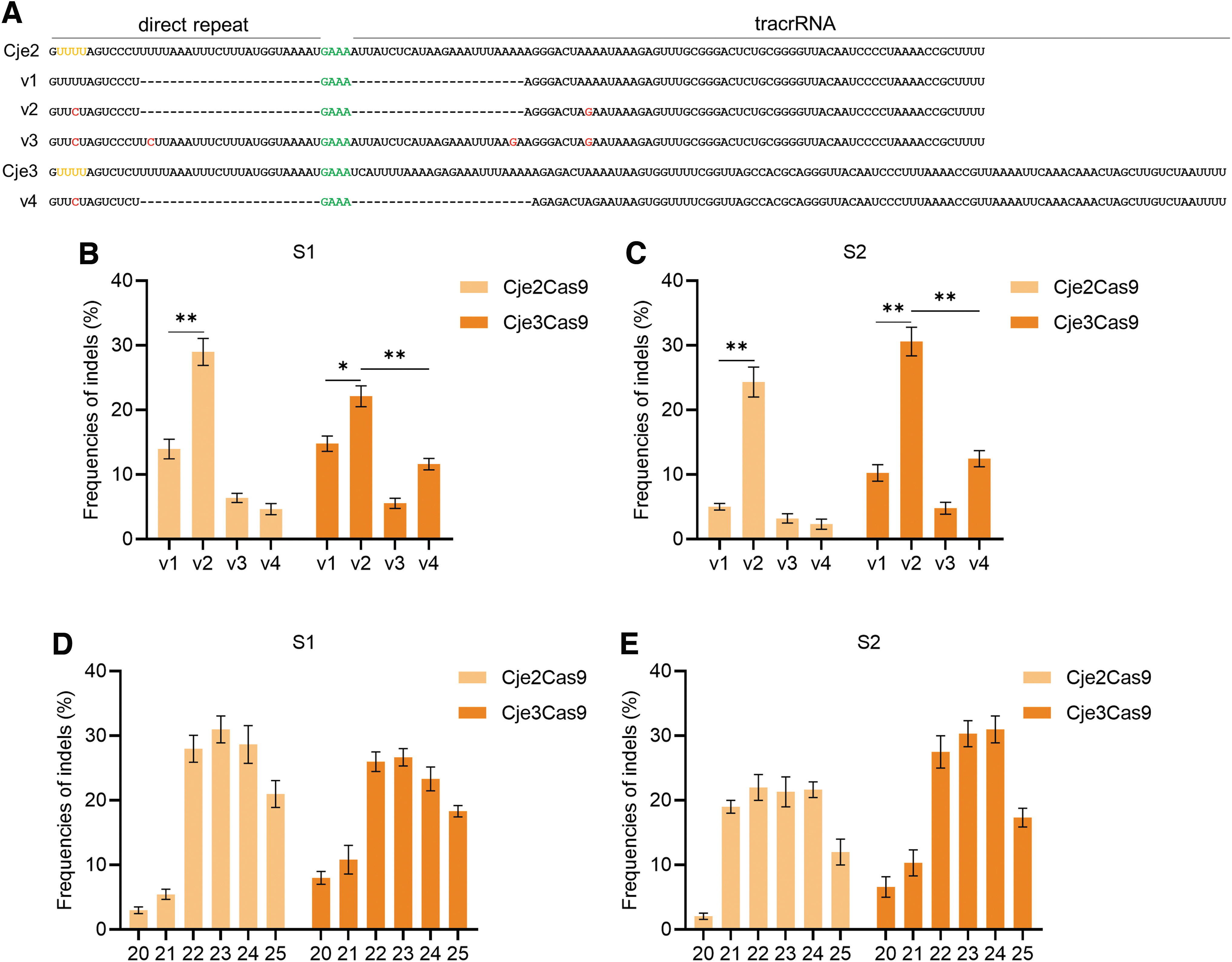
Engineered Cje2/3Cas9 gRNA augments editing frequency in human cells.
We disrupted poly Us in the original Cje2 and Cje3 gRNA scaffolds by swapping U-A base pair with C-G base pair (v2 and v4) (Fig. 3A and Supplementary Fig. S7). Moreover, a full-length Cje2 gRNA scaffold without consecutive Us (v3) was also constructed (Fig. 3A and Supplementary Fig. S7). It was shown that the gRNA scaffold v2 significantly improved editing efficiencies of both Cje2Cas9 and Cje3Cas9, whereas the full-length gRNA scaffold v3 showed low efficiency (Fig. 3B, C). Interestingly, Cje3Cas9 functions more efficiently when using scaffold v2 from Cje2 than using its original scaffold v4 (Fig. 3B, C). These results collectively indicated that gRNA scaffold v2 is the best performing architecture for both Cje2Cas9 and Cje3Cas9.
In addition, Cas9 orthologs prefer different optimal spacer length, and editing efficiencies vary as spacer length changes.8,19,20 To test the impact of varying spacer length on editing activities of Cje2/3Cas9, we designed six gRNAs of variable lengths from 20 to 25 nt for each endogenous site. All gRNAs contain an additional guanine at the 5′-terminal sequence to facilitate U6 promoter initiating transcription. 20 We illustrated at two endogenous sites that Cje2/3Cas9 function effectively when using 22- to 24-nt spacer, and activity decreased upon other lengths (Fig. 3D, E). For convenience, we recommended 23-nt spacer, and all target sites used in this study were 23 nt in length.
Collectively, these results illustrated that Cje2Cas9 and Cje3Cas9 generate highest editing efficiency with scaffold v2 and the optimal spacer length is 23 nt.
Cje2/3Cas9 mediated efficient genome editing and base editing in human cells
We assessed the efficacy of Cje2/3Cas9 for genome editing at 24 endogenous sites in HEK293T cells (Fig. 4A). As a result, Cje2Cas9 generated efficient indels at target sites with N4CYAK (K = T/G) PAMs, while showed lower efficiency at target sites with N4CYAM (M = A/C) PAMs (Fig. 4B). In contrast, Cje3Cas9 exhibited excellent compatibility with all N4CYAN PAMs, although it was slightly less efficient at N4CYAK PAM sites than Cje2Cas9 (Fig. 4B). Cje2Cas9 also cleaves at target sites with N4CYTT PAMs (Fig. 4B). These editing results in human cells were basically consistent with observations of PAM identification, which proved that Cje2Cas9 and Cje3Cas9 work effectively in human cells with corresponding N4CYWK and N4CYA PAMs, respectively.

Cje2/3Cas9-induced genome editing and base editing in human cells.
Despite the high degree of sequence similarity, conventional Cje1Cas9 could not recognize Cje2/3Cas9 PAMs (N4CYA) in human cells, highlighting the PAM specificity and applicability of Cje2/3Cas9 (Fig. 4B). In addition, Cje1Cas9–T913D (harboring the key T913D mutation) showed low efficiency in 3 of the 24 tested sites, demonstrating the importance of T913D mutation in PAM recognition (Fig. 4B).
BEs, mainly consisting of deaminases and Cas enzymes, install C.G-T.A or A.T-G.C transitions at specified DNA sites without requiring DNA double-strand breaks or a donor template.6,7 However, successful base editing entails a proper PAM adjacent to targeted sites, which limits the scope of genome editing.5,30 To test whether Cje2/3Cas9 are compatible with widely used deaminases to construct BEs, we constructed rA1-nCje2-CBE and rA1-nCje3–CBE by swapping the SpCas9 nickase in BE4max with Cje2Cas9 nickase (D8A) and Cje3Cas9 nickase (D7A) (Fig. 4C).
We found that rA1-nCje2/3-CBE showed low efficiency at tested sites in general (Fig. 4C). We suspected that Cje2/3Cas9, like other small Cas9 systems, may not be highly compatible with rA1 deaminase.20,31 Therefore, we applied eAID deaminase, which is more efficacious and compatible with effectors, to create eAID-nCje2-CBE and eAID-nCje3–CBE (Fig. 4C).31,32 Notably, the eAID-nCje2/3-CBE can induce more potent C-to-T conversions than rA1-nCje2/3-CBE, and eAID-nCje3–CBE had the highest editing efficiencies at most target sites (Fig. 4C and Supplementary Fig. S8).
We also generated 8e-nCje2/3-ABE, which proved efficient at inducing A-to-G conversion in genomic DNA (Fig. 4D and Supplementary Fig. S9). Through a side-by-side comparison at targeted sites with overlapping PAMs, we demonstrated that the Cje2/3Cas9 generated comparable indels and C-to-T base editing efficiency to the classic small Nme2Cas9 (Supplementary Fig. S10).
Taken together, these data further confirmed that Cje2/3Cas9 are potent gene editing tools that expand the scope of genome editing.
AcrIIA5, AcrIIA16, and AcrIIC1 can be used as off switches for Cje2/3Cas9
Bacterial CRISPR-Cas systems defend against bacteriophage infection with sequence-specific RNA-guided nucleases. 33 In response to the bacterial war on phage infection, numerous phages produce anti-CRISPR (Acr) proteins to block the function of CRISPR-Cas systems.33,34 Those natural brakes for CRISPR-Cas9 technologies have been playing an important role in manipulating genome modification in mammalian cells and organisms. 34 To clarify inhibitors blocking Cje2/3Cas9-mediated genome editing, we transfected HEK293T cells with Cje2/3Cas9, an sgRNA targeting S1 and 14 known Acr proteins (AcrIIA1–6, 11, 13, 16, and AcrIIC1–5).
Consequently, only AcrIIA5, AcrIIA16, and AcrIIC1 significantly inhibited DNA cleavage by both Cje2Cas9 and Cje3Cas9 (Fig. 5A, B), with the strongest inhibition presented by AcrIIC1 (Fig. 5A, B). Moreover, inhibitory effects of all three Acrs on Cje2/3Cas9 were positively correlated with dose (Fig. 5C). The three Acr proteins also inhibited the function of eAID-nCje2/3-CBE and 8e-nCje2/3-ABE systems, showing the potential of regulating nCje2/3-mediated base editing (Fig. 5D). The results demonstrated that the AcrIIA5, AcrIIA16, and AcrIIC1 proteins are also natural brakes for Cje2/3Cas9.
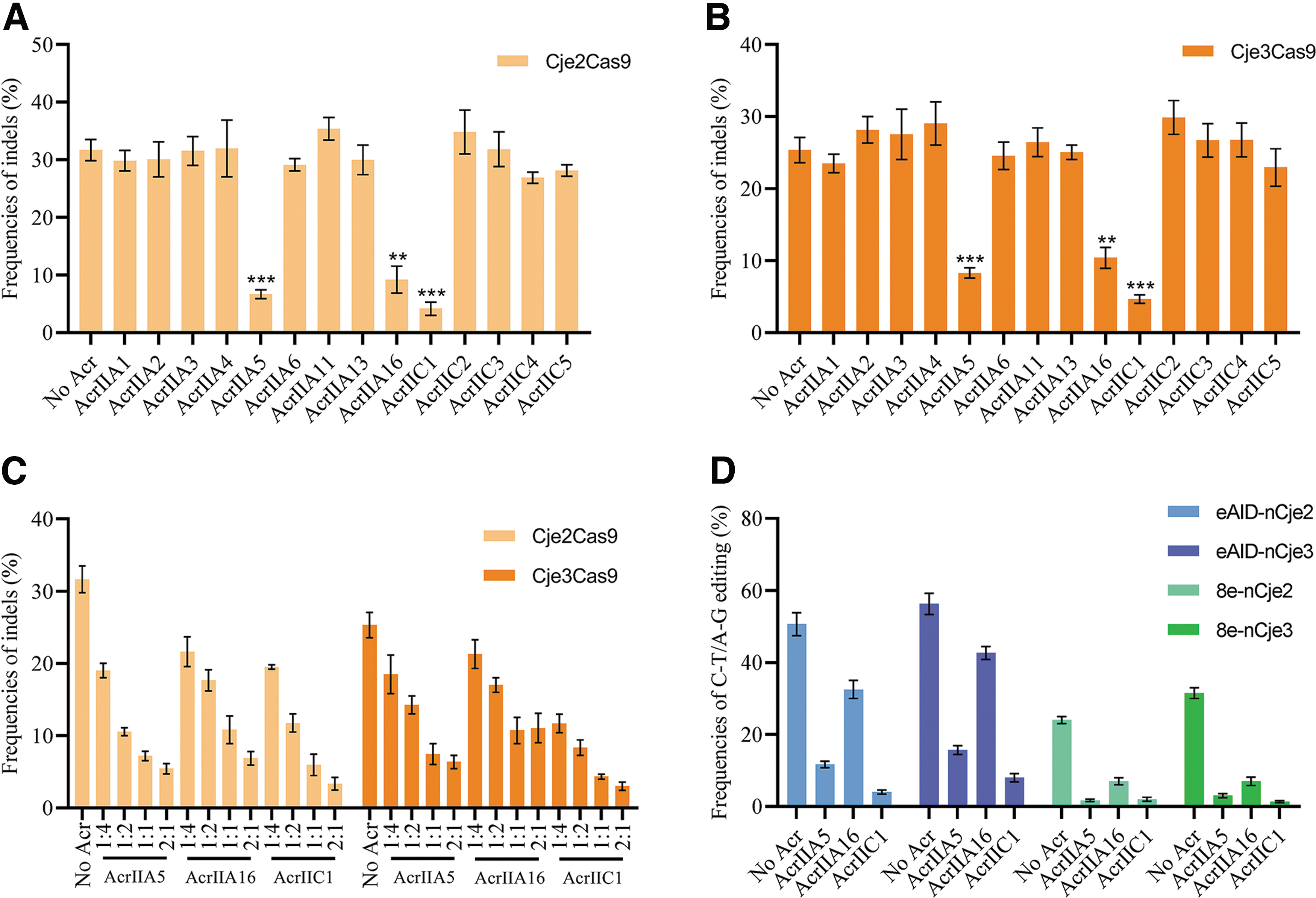
Acr proteins block the function of Cje2/3Cas9 in human cells.
Efficient gene editing in mice mediated by Cje3Cas9
In this study, we focused on Cje3Cas9 due to its simple N4CYA PAM and excellent compatibility with both CBE and ABE systems. We first tested the cleavage activity of Cje3Cas9 at six genomic sites with N4CYA PAMs in mice using sgRNAs targeting Pcsk9 and Tyr gene. After microinjection of Cje3Cas9 mRNA and gRNAs into mouse zygotes as previously reported,35,36 mouse zygotes were cultured in vitro and collected individually at the blastocysts stage for genotyping (Fig. 6A). At all sites tested, Cje3Cas9 exhibited high editing efficiencies ranging from 35.3% to 83.5% (Fig. 6A).
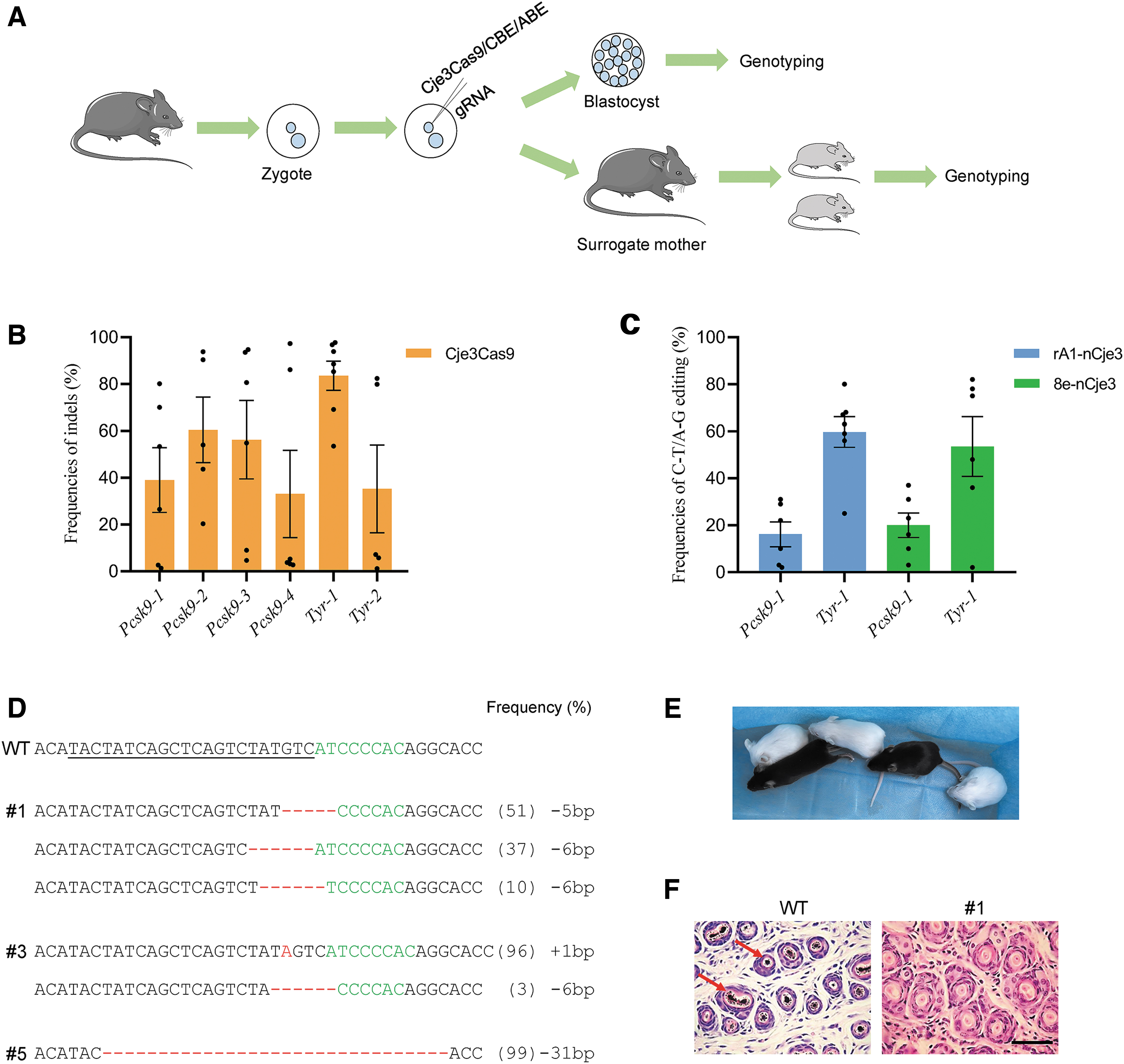
Cje3Cas9 efficiently induces genome editing in mice.
We also tested the gene editing activity of nCje3–BEs in mouse zygotes. Our results showed that both nCje3–CBE and nCje3–ABE efficiently induced base transitions at target sites with C-T efficiency of 16.2% to 59.7% and A-G efficiency of 20.9% to 53.5% (Fig. 6C), respectively.
Encouraged, we further attempted to generate mice models with mutations in Tyr using Cje3Cas9 to simulate human oculocutaneous albinism (OCA). 37 After microinjection, we transferred zygotes into surrogate mice. Of five pups obtained, three (60%) contained the expected editing outcomes at Tyr-1 site (Fig. 6D). They were completely albino mice in comparison with the wild-type (WT) mice (Fig. 6E). Histological H&E staining further confirmed the absence of melanin in the skin of a mutant but not in the WT mouse (Fig. 6F).
In addition, deep sequencing revealed that the average frequency of off-target editing in the mutant mice remained low (<1%) at possible off-target sites (POTs) (Supplementary Fig. S11A). These data demonstrated that the mouse model exemplified human OCA disease symptoms, which indicated that the Cje3Cas9 system is potent in gene editing and producing mouse models.
In vivo genome editing using all-in-one AAV–Cje3Cas9
The small size of Cje3Cas9 makes it potentially permissive to be delivered into adult mice by a single AAV vector. To test its potential for gene therapy, we constructed an AAV plasmid that drives Cje3Cas9 expression by Lp1b promoter and drives gRNA by U6 promoter (Fig. 7A). After examining the efficiencies of Cje3Cas9 to generate indels in N2a cells, we selected Pcsk9-1 site for in vivo editing because the editing efficiency at the site was high (Supplementary Fig. S12A), and disrupting Pcsk9 results in low blood cholesterol levels, indicating a low risk of cardiovascular disease.38,39

Cje3Cas9-mediated efficient in vivo genome editing through an all-in-one AAV delivery.
Using hepatotropic AAV serotype 8, we injected two groups of mice with 2 × 1011 GCs of AAV–Cje3Cas9 targeting Pcsk9-1 or PBS as a negative control (Fig. 7B). We collected mice serum before injection to test cholesterol level. Thirty days after injection, all mice were euthanized after collecting of serum to examine editing efficiency (Fig. 7B). Total liver DNA and RNA were isolated to analyze the expression of Cje3Cas9 and indels frequencies.
The qPCR confirmed the expression of Cje3Cas9 in mouse liver and deep sequencing confirmed the average indel frequencies at the Pcsk9-1 site were 16.4% in total liver, which is comparable with the efficiency of previously reported type II-A SpaCas9 in the same conditions (Fig. 7C and Supplementary Fig. S13A). 20 Deep sequencing revealed that off-target editing at POTs in the edited mice was low (Supplementary Fig. S11B).
In addition, the serum cholesterol levels in mutant mice treated with AAV–Cje3Cas9 significantly decreased in comparison with the PBS group 1 month later (Fig. 7D). Moreover, the serum ALT or ALB levels remained normal in the AAV–Cje3Cas9 group, from which we inferred that AAV–Cje3Cas9 expression did not cause liver damage (Fig. 7E, F). Histological examination of the liver by H&E staining further confirmed no signs of inflammation (Fig. 7G). Overall, these results confirmed that Cje3Cas9 can be delivered by an all-in-one AAV vector to induce robust genome editing in vivo.
In vivo adenine base editing using all-in-one AAV–8e-nCje3Cas9
Encouraged by the pilot study, we further tested whether the 8e-nCje3Cas9–ABE system could be packaged into a single AAV. Importantly, the small 8e-nCje3Cas9 and its gRNA just meet the AAV packaging limits (4.7 kb) and can be delivered by an all-in-one AAV vector (Fig. 8A). Targeted A-to-G editing in conserved RNA splicing signal of Pcsk9-2 site would disrupt Pcsk9 gene expression by inducing mRNA splicing defect (Supplementary Fig. S12B, C). 12 Similarly, 2 × 1011 GCs AAV-8e–nCje3Cas9 targeting Pcsk9-2 site were injected into mice tail veins and then editing efficiency was detected 30 days later (Fig. 8B).

8e-nCje3Cas9-induced in vivo base editing through all-in-one AAV delivery.
Deep sequencing revealed that the average A-to-G editing efficiency of Pcsk9-2 in the liver is 2.3% (Fig. 8C). In addition, no significant decrease in serum cholesterol levels was observed in the AAV-8e–nCje3Cas9 group (Fig. 8D). It has been reported that low titers (1011 GCs) of AAV is less efficient than high titers (1012 GCs) when using AAV–BE system in mouse liver. 40 Therefore, we speculated that higher doses of AAV could improve the efficiency of AAV-8e–nCje3Cas9. Subsequently, 4 × 1012 GCs AAV-8e–nCje3Cas9 were injected into mice tail veins (Fig. 8B).
The qPCR detection showed that the expression of 8e-nCje3Cas9 in 4 × 1012 GCs group significantly increased compared with 2 × 1011 GCs group (Supplementary Fig. S13B). Notably, the average 12.1% A-to-G editing efficiency of 4 × 1012 GCs group was much higher than that of 2 × 1011 GCs group (Fig. 8C). More importantly, serum cholesterol levels in 4 × 1012 GCs group significantly reduced (Fig. 8D). Only low off-target mutations were observed at POTs in the edited mice (Supplementary Fig. S11C).
These results confirmed that 8e-nCje3Cas9–ABE system can be delivered by an all-in-one AAV vector to induce efficient base editing in vivo.
Discussion
In this study, we have characterized two compact Cas9 orthologs termed Cje2Cas9 and Cje3Cas9, derived from C. jejuni, and efficiently induced genome modification in vivo. Importantly, the N4CYA PAMs recognized by Cje3Cas9 are simpler compared with the complex N3VRYAC PAMs of conventional Cje1Cas9, holding a potential for expanding the targeting scope.
By disrupting poly Us in sgRNA scaffold and using 23-nt spacer, we enhanced the editing efficiency of Cje2/3Cas9. We also validated that Cje2/3Cas9 and Cje2/3Cas9-based BEs potently mediated gene editing in both human cells and mice. AcrIIA5, AcrIIA16, and AcrIIC1 proteins block the function of Cje2/3Cas9, having potential to precisely regulate Cje2/3Cas9-mediated gene editing. Moreover, Cje3Cas9 or 8e-nCje3Cas9 and its sgRNA can be packaged into a single AAV vector to induce effective in vivo genome editing.
This study comprehensively characterized Cje2/3Cas9 and attested their versatility and solidity in vivo genome editing. Recently, the miniature Cas12f and TnpB nucleases have been engineered for efficient editing in human cells, further enriching the toolsets of small gene editing nucleases.41–45
In this study, Cje2Cas9 and Cje3Cas9 were successfully excavated by exploring natural mutations in pivotal PAM-interacting residues based on structural information. In our previous study, we verified that rationally swapping PIDs of SpaCas9 with those of its orthologs broadens targeting scope. 20 These approaches can be applied to other Cas9 orthologs, such as SaCas9 and St1Cas9 to further expand their targeting range. Thus, structural information of known Cas enzymes can be a valuable resource for identifying new Cas9 orthologs that recognize relaxed PAMs.
We have validated that Cje3Cas9-mediated ABE8e can be packaged into single AAV vector and induce A-to-G base editing in vivo. However, most Cas9-mediated BEs are limited by the cargo capacity of AAV, including glycosylase BEs,46–48 dual BEs,49–52 and prime editors. 53 It has been reported that Cas9 tolerates large single deletions in the REC2, REC3, HNH, and RuvC domains while maintains function. 54 The truncated version of Cas9s might enable those BEs to be delivered by an all-in-one AAV vector for in vivo genome editing and gene therapy.
Conclusion
In summary, we developed two new small Cas9 orthologs, Cje2Cas9 and Cje3Cas9, that efficiently induced genome editing in vivo. The simple N4CYA PAMs of Cje3Cas9 significantly broaden the targeting range of CjeCas9 orthologs. Importantly, the small size of Cje3Cas9 makes gene editing and base editing in adult mice a reality. Cje2Cas9 and Cje3Cas9 are promising tools for therapeutic applications.
Footnotes
Acknowledgments
We thank Peiran Hu and Nannan Li for their efforts in microinjection and zygotes transferring.
Authors' Contributions
Z.Q.L., L.L., and Z.J.L. conceived and designed the experiments. S.C. and Z.Q.L. performed the experiments. S.C., Z.Q.L., W.X., and H.Y. analyzed the data. Z.Q.L. and S.C. wrote the article. All authors have read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financially supported by the National Key Research and Development Program (Grant No. 2018YFB1404205, L.X.L.) of China Stem Cell and Translational Research (Grant No. 2017YFA0105101, Z.J.L.) and The National Natural Science Foundation of China (Grant Nos. 32170543 and 31970574, Z.J.L.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.