
Abstract

How do you control CRISPR? This is a critical question in the gene editing field, but humans aren't the only organisms who need CRISPR to cut in the right place at the right time. Prokaryotes that use CRISPR as an anti-phage defense system also need to keep CRISPR in check and prevent “autoimmunity”—a potentially lethal event where a CRISPR system targets its own genome for destruction. Indeed, the evolutionary success of CRISPR-Cas systems themselves also rests on their ability not to kill their prokaryotic hosts.
In an elegant new study in Cell, 1 Rachel Workman, Josh Modell, and colleagues used an unbiased transposon screen to find regulators of CRISPR-Cas9 activity in Staphylococcus aureus. By treating a mutagenized population of bacteria with a deadly phage, they selected for the bacterial mutants with the strongest and fastest immune response. Very surprisingly, the highest-performing mutants had transposon insertions in CRISPR locus itself. Specifically, the transposon insertions were concentrated in the promoter and 5′ end of an enigmatic RNA called tracr-L.
The trans-activating CRISPR-RNA (tracrRNA) is a critical part of type II CRISPR-Cas systems. 2 The tracrRNA binds to CRISPR-RNAs (crRNA), facilitating their processing by RNaseIII. Cas9 is then loaded with this dual tracrRNA:crRNA structure. For simplicity, many CRISPR applications use a chimeric RNA molecule called a single-guide RNA, in which the processed forms of the tracrRNA and crRNA are linked in a single molecule. 3
When the tracrRNA was first discovered by Emmanuelle Charpentier's group in 2011, they noted an oddity. The unprocessed tracrRNA was present in two forms: an 89-nucleotide “short” form (tracr-S) and a 171-nucleotide “long” form (tracr-L). As the short form alone is sufficient to facilitate crRNA processing and formation of the loaded Cas9 ribonucleoprotein (RNP), the role of the long form was left a mystery.
Ten years later, we now know the role of the long-form tracrRNA. When tracr-L is disrupted, expression of CRISPR-Cas components is massively increased, giving anti-phage immunity a huge boost. This establishes that tracr-L is a negative regulator of CRISPR-Cas expression and activity. We also now know why tracr-L is used to dampen CRISPR-Cas: loss of tracr-L activity leads to increased CRISPR-Cas autoimmunity and death for the bacterial host.
So, how does tracr-L do its job? Intriguingly, Workman et al. found that tracr-L contained an 11-nucleotide stretch that matched the DNA sequence at the Cas9 transcriptional start site. Furthermore, they spotted an “NGG” positioned in exactly the right place to be a protospacer adjacent motif. Could tracr-L be mediating “natural CRISPRi”?
CRISPR interference, or CRISPRi, is a technique used for programmed transcriptional repression.4,5 It generally uses a full-length 20-nucleotide spacer with a catalytically dead Cas9. However, the authors were able to show that this 11-nucleotide match sequence was able to hit the sweet spot of recruiting a catalytically active Cas9 to bind, but not cleave, its own promoter. Remarkably, the structure and function of the tracr-L RNA is quite similar to the engineered single-guide RNAs used in CRISPR applications today (Fig. 1).
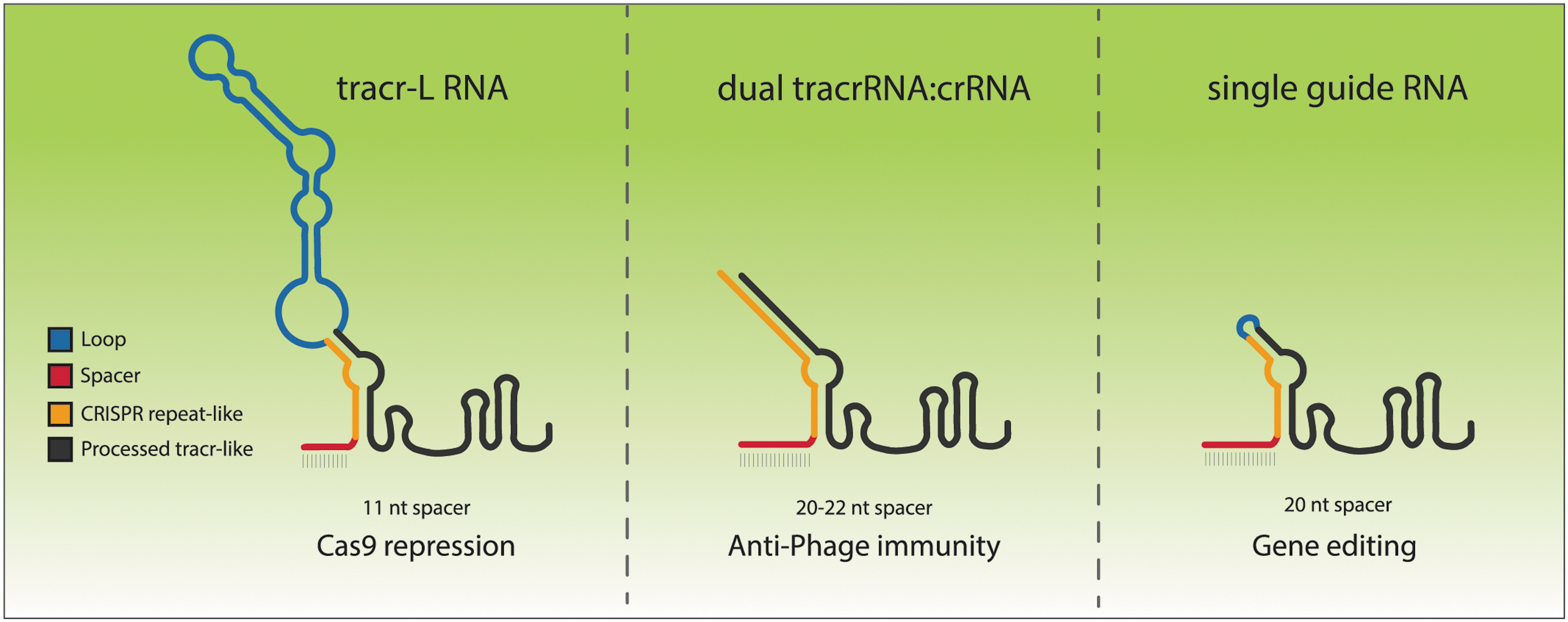
Comparison of different RNAs that can program Cas9. Some bacteria use the single-molecule RNA tracr-L to direct transcriptional repression of the Cas9 promoter (left panel), while a processed dual RNA tracrRNA:crRNA controls anti-phage immunity (middle). Single-guide RNAs are chimeric molecules commonly used in gene editing (right), where a crRNA-like sequence (red, orange) is attached to a processed tracr-like sequence (black) with a small loop (blue).
The balancing act between CRISPR anti-phage activity and CRISPR autoimmunity has been studied in some type I CRISPR-Cas systems and, to a lesser degree, type III CRISPR-Cas systems. CRISPR-Cas systems are disseminated by horizontal transfer, and in almost all studied cases, CRISPR-Cas transcriptional regulation is achieved by integrating CRISPR into a preexisting transcriptional network (as recently published by Smith et al. 6 and extensively reviewed by Shivram et al. 7 ).
The advantage of using a component of the CRISPR locus itself as a regulatory mechanism is clear: intrinsic control of CRISPR-Cas activity should facilitate its safe acquisition by new hosts. Indeed, Workman et al. show wide dissemination of tracr-L elements in bacteria that encode Cas9. Nonetheless, more than half of CRISPR-Cas loci surveyed did not have a tracr-L element, suggesting the existence of other regulatory mechanisms that control Cas9.
What remains a fascinating question is the activator(s) of CRISPR-Cas immunity in the cell. The CRISPR-Cas system is regulated by a negative feedback loop in which its own activity dampens its expression. To break this loop, CRISPR-Cas expression levels and tracr-L RNP repressor activity must be decoupled. This can plausibly occur through several scenarios, such as selective destruction of tracr-L. The authors also showed that by increasing the number of spacers, they were able to increase Cas9 levels by outcompeting the repressive tracr-L RNA with bona fide CRISPR-RNAs.
Finally, and perhaps most exciting, is the possibility that phage-encoded Cas9 inhibitors called anti-CRISPRs could activate Cas9 expression by preventing the tracr-L RNP from binding the Cas9 promoter. Anti-CRISPR function is highly dose dependent,8,9 and overexpression of Cas9 in response to phage inhibition could be the perfect anti-anti-CRISPR response to shift the balance in favor of bacterial immunity. Indeed, the bifunctional anti-CRISPR AcrIIA1 uses a similar strategy to counter CRISPR-Cas immunity. By autoregulating its own transcription, it tunes its expression to match Cas9 levels. 10
Looking forward, we still have much to learn about our favorite gene editing tools from the prokaryotic world. Workman et al. have shown that bacteria deploy single-guide RNAs to enable CRISPRi and regulate Cas9, using spacer length to toggle between DNA cleavage and gene repression. There is an ever-expanding universe 11 of new CRISPR-Cas systems within an enormous diversity of bacteria, archaea, and even phages.12,13 Each one of these organisms has the potential to evolve new and fascinating ways of using and controlling their immune systems, informing the advancement of CRISPR tools for years to come.