
Abstract
The development of clustered regulatory interspaced short palindromic repeats/CRISPR associated protein 9 (CRISPR-Cas9)-mediated gene modification has opened an exciting avenue of targeting genes to study the pathogenesis of diseases and to develop novel therapeutic concepts. However, as the effector protein Cas9 is of bacterial origin, unwanted side effects due to a host immune response against Cas9 need to be considered. Here, we used the syngeneic model of BAL17CNS-induced primary lymphoma of the central nervous system (PCNSL, CNS) in BALB/c mice to address this issue. Surprisingly, stable expression of Cas9 in BAL17CNS (BAL17CNS/Cas9) cells rendered them unable to establish PCNSL on intracerebral transplantation. Instead, they induced a prominent intracerebral immune response mediated by CD8 T cells, which lysed BAL17CNS/Cas9 cells via perforin. In addition, B cells contributed to the immune response as evidenced by serum anti-Cas9 antibodies in BALB/c mice as early as day 8 after transplantation of BAL17CNS/Cas9 cells. In athymic BALB/cnu/nu mice, NK cells mounted a vigorous intracerebral immune response with perforin-mediated destruction of BAL17CNS/Cas9 cells. Thus, in the CNS, perforin produced by NK and CD8 T cells was identified as a mediator of cytotoxicity against BAL17CNS/Cas9 cells. These observations should be taken into account when considering therapeutic CRISPR-Cas9-mediated tumor cell manipulation for PCNSL.
Introduction
Primary lymphoma of the central nervous system (PCNSL, CNS) is a unique diffuse large B cell lymphoma entity confined to the CNS with a less favorable prognosis as compared to extracerebral counterparts.1,2
Selective studies of tiny human PCNSL samples usually obtained by stereotactic neurosurgery have elucidated the genomic, transcriptional, and proteomic landscape of PCNSL. 3 Determination of the functional in vivo role of the individual factors identified now awaits analysis in preclinical animal models.
Establishment of the syngeneic PCNSL model in BALB/c mice using BAL17CNS lymphoma cells that reflects major characteristics of human PCNSL allowed functional studies to decipher PCNSL pathogenesis. 4 Combining this preclinical model with modern genome-editing techniques is an attractive tool to precisely dissect the relevance of individual genes and may contribute to development of novel therapeutic regimens in PCNSL.
The genome editing CRISPR-Cas9 (clustered regulatory interspaced short palindromic repeats/CRISPR associated protein 9) system in Streptococcus pyogenes (Sp), Staphylococcus aureus (Sa), or other Cas enzymes utilizes noncoding RNAs that guide Cas9 protein to generate double-strand breaks. DNA damage is repaired by the non-homologous end joining DNA repair pathway. This mechanism results in insertions and deletions that are used to disrupt gene function (for a review, see Katti et al 5 and Naldini 6 ). This elegant technology allows genetic manipulations, including addition and knock-out of genes in mammalian cells, and may yield in vivo preclinical mouse models easily and rapidly. It circumvents the generation of genetically manipulated mouse models from embryogenesis, which are less precise; however, they are more time- and animal-consuming and cost-intensive.
Despite these promising advantages, the CRISPR-Cas9 system, when applied in mammals in vivo, may be associated with unwanted side effects. Proper functioning of this technology requires Cas proteins, which are key components of an ancient bacterial adaptive immune system.5,7–10 Thus, one such pitfall is the potential induction of host immune reactions, since Cas proteins including SpCasp9 and SaCas9 may be immunogenic. Approximately 40% of the human population and 20% of school-aged children are colonized with Sa and Sp, respectively, and pre-existing adaptive immunity to the Cas9 orthologs derived from these bacteria is frequent in humans. 11
There is also evidence of memory T cells to Cas9 in the human population,11–15 and antigen-reactive T cells against SpCAs9 and SaCas9 were identified in 68% and 78% in a group of 18 healthy donors. 16
So far, it is unknown whether an immune response to Cas9 may play a role in vivo in murine models of CNS disorders. Regarding pre-existing immunity to Cas9, the situation of mice housed under specific pathogen free (SPF) conditions that preclude contact with Sp and Sa differs from that of humans; nevertheless, their microenvironment is not sterile. With respect to the CNS, the blood–brain barrier strictly restricts entry of immune cells and soluble factors into the brain; further, lack of transplant rejection has long been considered as a major hallmark of CNS immune privilege. Thus, one might expect that the CNS is particularly suitable for in vivo studies using the CRISPR-Cas9 system.
Here, we have generated BAL17CNS/Cas9 lymphoma cells as a prerequisite for functional studies of genes involved in the lymphomagenesis of PCNSL and transplanted them into the brain of BALB/c mice by stereotaxy. Surprisingly, in contrast to parental BAL17CNS lymphoma cells, Cas9-stably expressing BAL17CNS cells failed to induce PCNSL. Instead, a cytotoxic immune response mediated by CD8 T cells and—in the absence of CD8 T cells—by NK cells both of which expressed perforin granules in a polarized pattern to closely attached intracerebral BAL17CNS/Cas9 cells induced their lysis.
Materials and Methods
Lymphoma cells
The mammalian codon-optimized S. pyogenes Cas9 protein expression vector 17 was used in the ViraPower lenti-virus expression system (Invitrogen, Karlsruhe, Germany) according to the manufacturer's instructions. The vector LentiCas9-Blast was a gift from Feng Zhang (Addgene plasmid no. 52962; http://n2t.net/addgene:52962).
In brief, BAL17CNS lymphoma cells 4 were incubated with virus particles produced in 293FT cells. Selection of the bulk population started 48 h after transduction, followed by single clone selection, which yielded the Cas9-expressing cell line designated BAL17CNS/Cas9.
BAL17CNS and BAL17CNS/Cas9 cells were cultured in Iscove's Modified Dulbecco's Medium (Invitrogen, Karlsruhe, Germany) with 20% fetal calf serum. 293FT cells were cultured in Dulbecco's Modified Eagle's Medium (Invitrogen) with 10% fetal calf serum. The absence of Mycoplasma was tested in all cells under culture (Mycoplasma Gel Detection Kit, Biotools, Madrid, Spain).
Western blot analysis
Expression of Cas9 in BAL17CNS/Cas9 lymphoma cells was verified by Western blot. As primary antibody, mouse anti-Cas9 monoclonal antibody (Millipore, Darmstadt, Germany) was used, followed by peroxidase-conjugated rat anti-mouse Ig (Dianova, Hamburg, Germany) and the Pierce™ ECL Western Blotting-Substrate (Thermo Fisher Scientific, Dreieich, Germany). The chemiluminescent signal was detected by the ChemiDoc™ XRT+ system (Bio-Rad, Göttingen, Germany) using the Image Lab™ software (Bio-Rad).
Animals
Female BALB/c and BALB/cnu/nu mice at the age of 6–8 weeks were obtained from Charles River (Sulzfeld, Germany). Animals were housed in cages up to five mice each in a controlled microenvironment (SPF conditions excluding the presence of hemolytic Streptococci group A and Sa) with a temperature range of 22 ± 2°C and a relative humidity of 45–65% on a 12/12 h light/dark cycle. They were supplied with water and food ad libitum. Experiments were performed according to the Institutional Animal Care and Use Committee (IACUC) under permits from the Government of North-Rhine-Westphalia approved by the responsible ethical committee (AZ 84-02.04.2015.A356 and AZ 81-02.04.2020.A496).
Experimental in vivo procedure
Tumor cells were implanted intracerebrally (i.c.) under general anesthesia with 100/10 mg/kg body weight ketamine/xylazine (Ketaset®; Zoetis, Berlin, Germany; Serumwerk, Bernburg, Germany). Parental or genetically modified 5 × 105 BAL17CNS lymphoma cells suspended in a total volume of 3 μL phosphate-buffered saline (PBS) were implanted stereotactically into the right frontal white matter (1.5 right lateral, 1 mm anterior, 3 mm depth from bregma) using a Hamilton syringe fixed in a holder on a rodent stereotaxic frame.
Tumor cells were applied very slowly, and the needle was removed after 5 min. The wound was closed with absorbable suture material (vicryl 4-0; Ethicon, Norderstedt, Germany). For postoperative analgesia, mice were treated with tramadol (Tramal®; Grünenthal GmbH, Aachen, Germany) at a concentration of 0.5 mg/mL in drinking water for 3 days. Animals were sacrificed at days 1, 2, 3, 4, 5, 8, and 16 after injection (p.i.), respectively.
For all experiments, groups consisted of six mice each. Body weight was determined daily and calculated as percentage of the body weight determined immediately before i.c. transplantation. Mice were monitored daily for clinical symptoms according to a clinical score (Supplementary Table S1).
Histopathology, immunohistochemistry, and immunofluorescence
I.c. tumor growth was evaluated in all six mice per group. Snap-frozen brains were cut entirely into 10 μm horizontal cryostat sections (360–410 sections per mouse). Paraffin-embedded brains were cut into serial 1 μm sections; starting anteriorly at the level of the lateral ventricle, 320–360 sections were cut per mouse. Every 10th section was stained by hematoxylin and eosin (HE). Sections adjacent to HE-stained frozen and paraffin sections, respectively, were used for immunohistochemistry and immunofluorescence (for antibodies see Supplementary Table S2).
Determination of serum anti-Cas9 antibodies
For analysis of anti-Cas9 antibodies, serum was isolated by centrifugation of blood samples obtained by cardiac puncture. The wells of 96-well microtiter plates were coated with monoclonal rabbit anti-Cas9 capture antibody (5 μg/mL, clone JM11-55; Novus Biologicals, CO, USA) at 4°C overnight. After blocking with 1% bovine serum albumin in PBS, 100 μL Cas9 solution (5 ng/μL Cas9 protein in reconstitution solution; Sigma, Taufkirchen, Germany) was added. As controls, reconstitution solution without Cas9 was used.
Mouse sera (150–200 μL volume) adjusted to a total volume of 1.2 mL with PBS were added to Cas9-coated and control wells in triplicates. Incubation was followed by 0.25 μg/mL goat anti-mouse IgM+IgG+IgA (H+L) (Millipore). Reactivity was detected with the Alkaline Phosphatase Yellow Liquid Substrate System (Sigma), terminated by addition of 20 μL 0.4 M NaOH. Absorbance at 405 and 620 nm was determined by use of the Multiskan FC ELISA reader (Thermo Fisher Scientific). Data were corrected for background values (405 nm value minus 620 nm value) assessed in triplicate.
The mean OD value for the Cas9 protein and control reaction was calculated for each serum sample, and the OD ratio (ODCas9/ODControl) was determined for each serum sample.
Statistical analysis
Differences in clinical score and body weight between mice that have been transplanted with BAL17CNS/Cas9 or BAL17CNS cells were tested by two-way ANOVA with Geisser-Greenhouse correction. All statistical analyses were carried out with GraphPad PRISM 7.0 (La Jolla, CA, USA).
Results
Characterization of the BAL17CNS/Cas9 cell line
Clones derived from Cas9-transduced BAL17CNS cells were analyzed for expression of Cas9. The clone that showed the maximal Cas9 expression in Western blot studies, that is, E5, was used for all experiments as BAL17CNS/Cas9 cell line (Fig. 1A; Supplementary Fig. S1A).
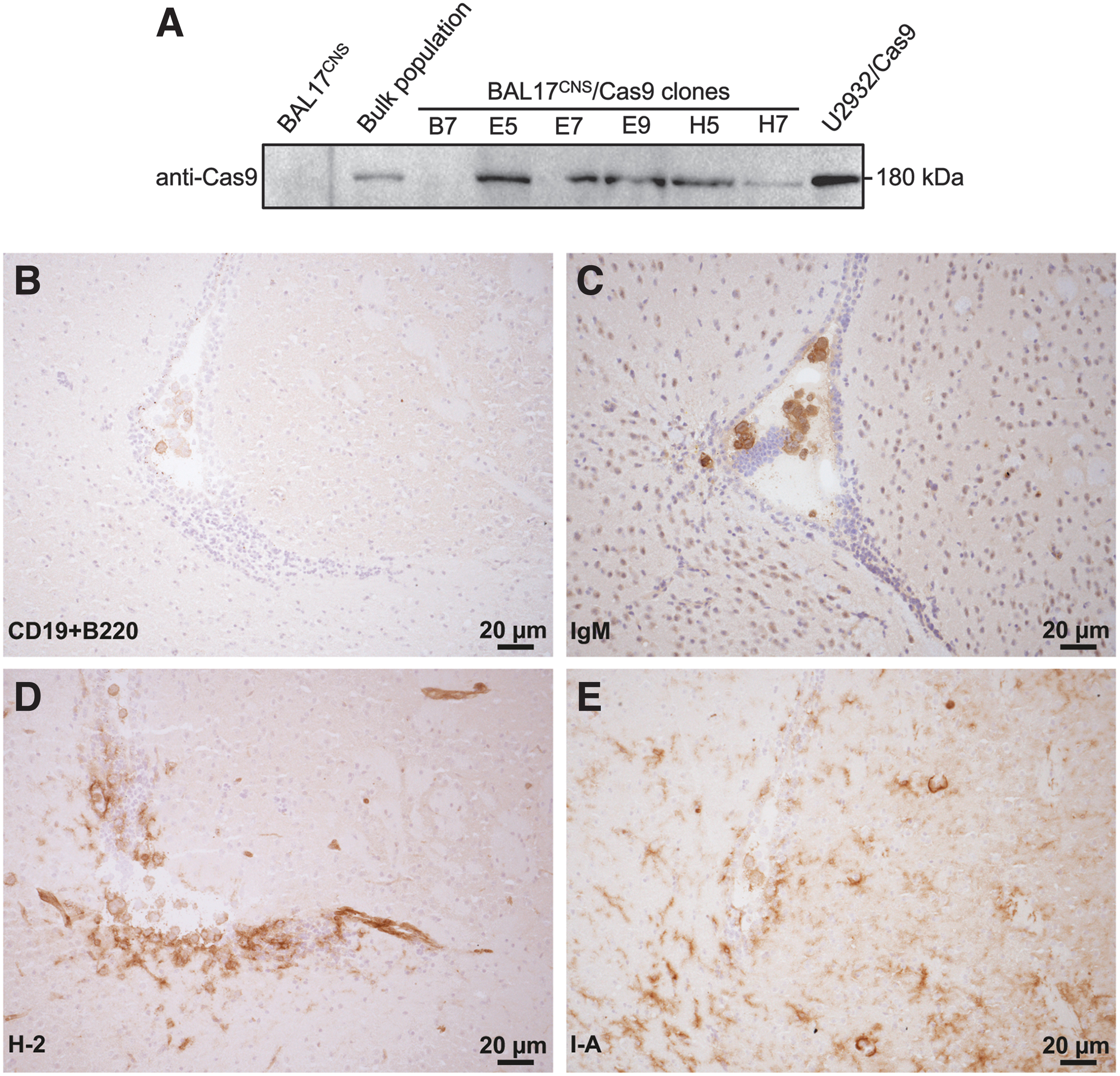
Characterization of BAL17CNS/Cas9 cells.
BAL17CNS/Cas9 cells were confirmed to carry the same functional immunoglobulin heavy chain rearrangements such as the parental BAL17CNS cells (Supplementary Fig. S1B). BAL17CNS/Cas9 cells were not impaired in their proliferative activity in vitro (Supplementary Fig. S1C). The CD19+ B220+ IgM+ MHC class I+ MHC class II+ immunophenotype of BAL17CNS/Cas9 cells (Fig. 1B–E) was identical to BAL17CNS cells. 4
Thus, BAL17CNS/Cas9 cells are similar to their parental BAL17CNS cell line.
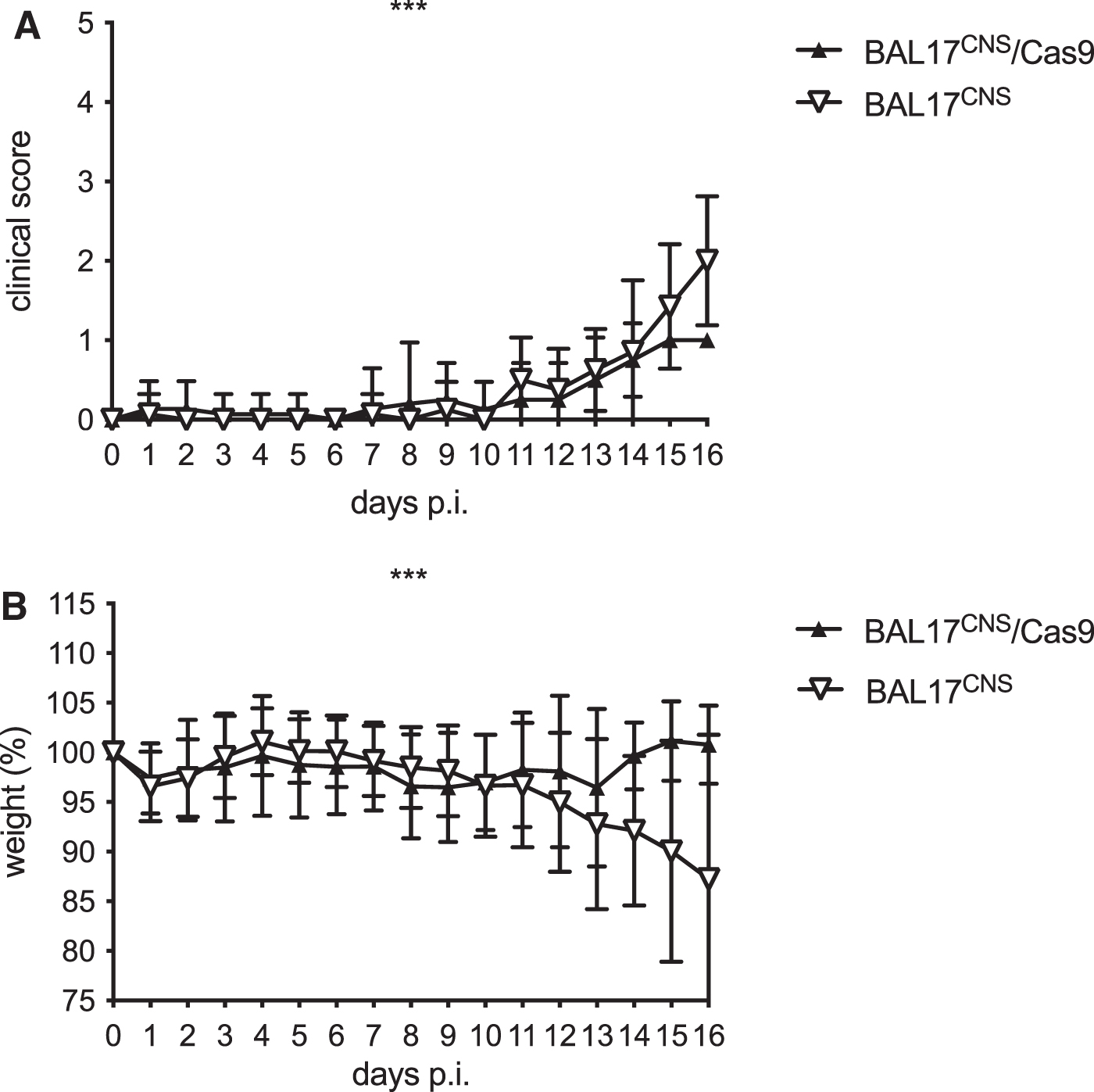
Clinical course of BALB/c mice after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 cells or BAL17CNS cells
Mice of both experimental groups lost body weight and developed clinical symptoms on the operation procedure, from which they recovered (Fig. 2A, B). Thereafter, body weight remained stable in mice that had received BAL17CNS/Cas9 lymphoma cells (Fig. 2B). These animals developed mild clinical symptoms that persisted until day 16 p.i. (Fig. 2A). In contrast, mice that had been transplanted with BAL17CNS lymphoma cells developed progressive clinical symptoms in association with steadily declining body weight from day 12 to 16 p.i. (Fig. 2A, B).

Clinical score and body weight of BALB/c mice after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 or BAL17CNS cells.
Upon i.c. transplantation of 5 × 105 BAL17CNS/Cas9 cells, BALB/c mice develop an i.c. immune response, resulting in rejection of lymphoma cells
To identify the cause underlying the discrepant clinical course of disease, a detailed morphological analysis was carried out.
Noticeably, none of the mice transplanted with BAL17CNS/Cas9 cells showed evidence for solid lymphoma growth in the brain parenchyma (Fig. 3A). Only a few tumor cells were present at day 8 p.i. (Fig. 3A), decreasing up to day 16 p.i. In striking contrast, in BALB/c mice that had received BAL17CNS cells, brain parenchymal lymphoma was prominent at day 8 p.i. (Fig. 3B), further progressing to day 16 p.i.

BAL17CNS/Cas9 lymphoma cells fail to induce PCNSL in BALB/c mice.
Although a few BAL17CNS/Cas9 cells were associated with a highly cellular inflammatory infiltrate, predominantly located in the lateral ventricle and the adjacent periventricular brain parenchyma (Fig. 3C, D), inflammation was absent from mice that had received BAL17CNS cells (Supplementary Fig. S2A, B). In BAL17CNS/Cas9-transplanted mice, inflammatory infiltrates consisted of large granular lymphocytes (Fig. 3D) corresponding to CD335+ NK cells (Fig. 4A), prominently activated F4/80+ MHC class I+ MHC class II+ microglia/macrophages (Fig. 4B–D), CD11c+ myeloid cells (Fig. 4E), and CD8 T cells (Fig. 4F), which increased up to day 16 p.i. and expressed the CD62L antigen (Fig. 4H, I).
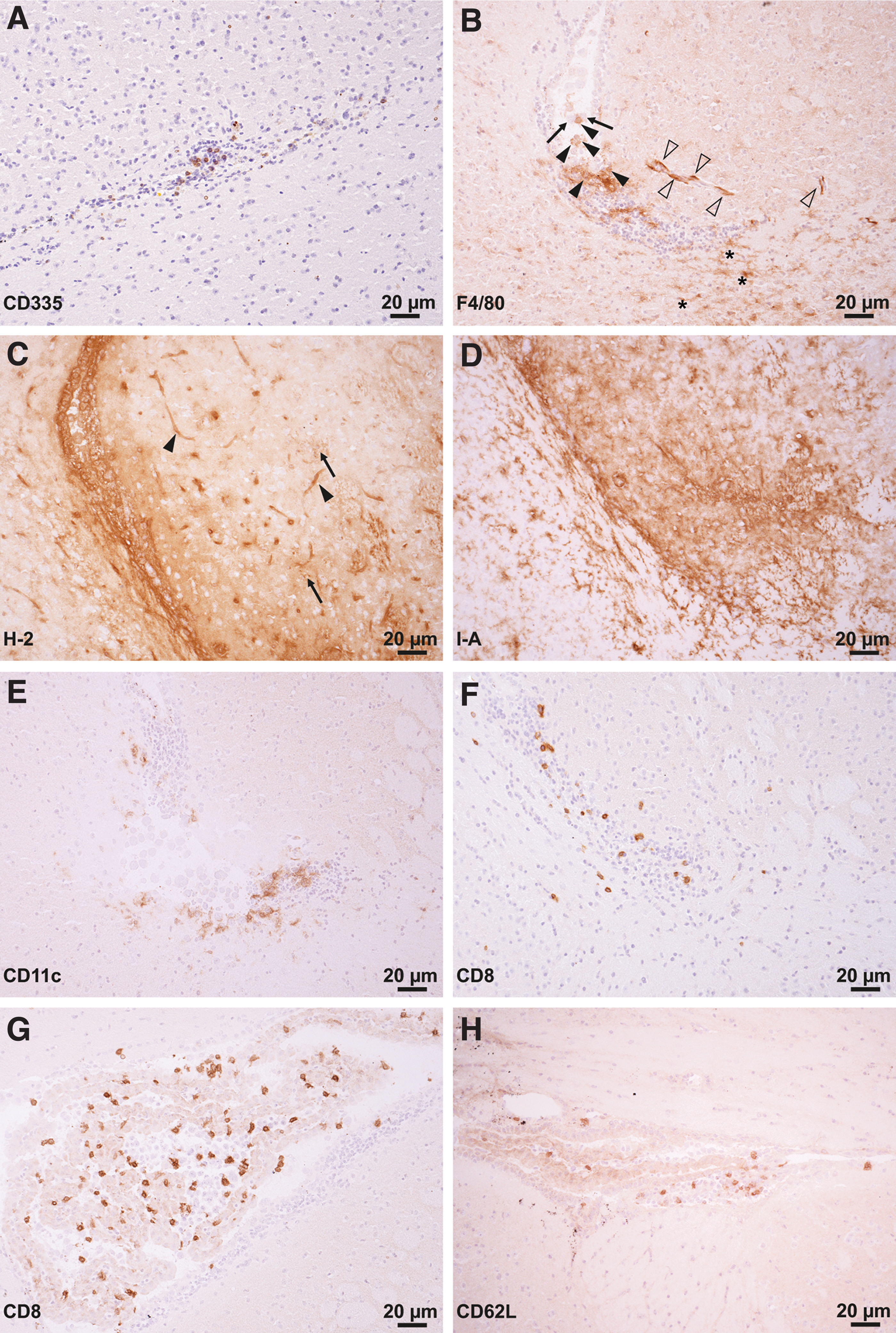
Phenotypic composition of the inflammatory infiltrates in the brains of BALB/c mice at day 8 and 16 after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 cells.
Highly activated, blastic CD8 T cells expressed perforin in a polarized manner toward intimately associated lymphoma cells (Fig. 5A–F). Lymphoma cells mostly were in various stages of cell death, ranging from increased nuclear basophilia with condensed, clumped chromatin, cytoplasmic eosinophilia to cells with apoptotic bodies and, finally, cellular debris (Fig. 5G). Severe inflammation even resulted in brain tissue necrosis (Figs. 3A, C and 5A, C).
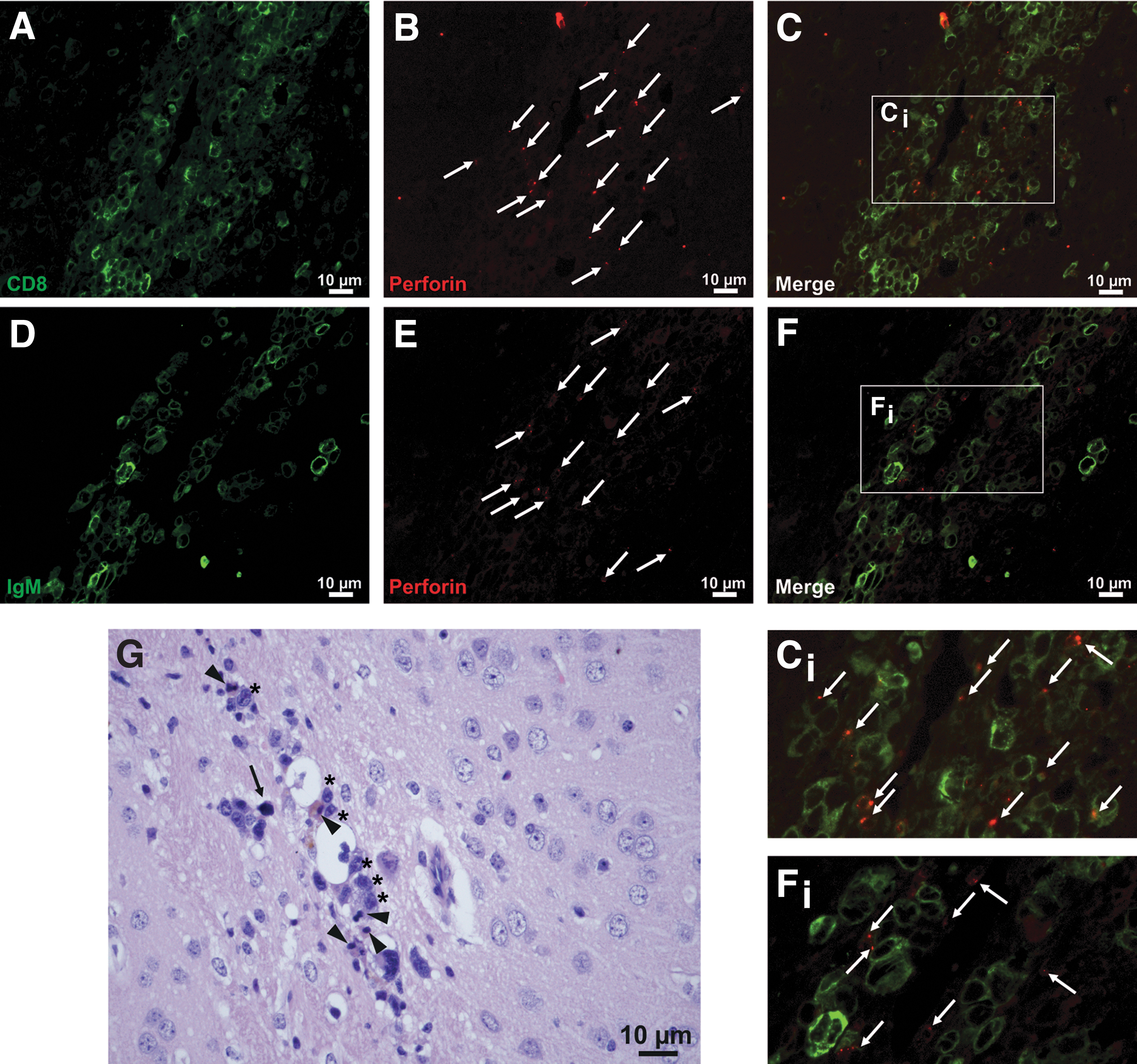
CD8 T cells kill i.c. BAL17CNS/Cas9 lymphoma cells via perforin in BALB/c mice.
These experiments reveal that CD8 T cells mount a cytotoxic reaction against BAL17CNS/Cas9 lymphoma cells in the brain. This prominent immune response with perforin-mediated tumor cell death prevented the development of parenchymal lymphoma in the brain of BALB/c mice.
Clinical course of BALB/cnu/nu mice after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 cells or BAL17CNS cells
To address the question whether the anti-BAL17CNS/Cas9 lymphoma cell immune response is mediated exclusively by CD8 T cells, BALB/cnu/nu mice were i.c. transplanted with BAL17CNS/Cas9 or BAL17CNS cells.
Clinically, BALB/cnu/nu mice transplanted with BAL17CNS/Cas9 cells developed less severe clinical disease as compared with BALB/cnu/nu mice that had received BAL17CNS cells (Fig. 6A). Postoperatively, up to day 16 p.i., mice that had been transplanted with BAL17CNS/Cas9 cells fully regained body weight, whereas mice transplanted with BAL17CNS cells showed a steady decline of body weight (Fig. 6B).
Clinical score and body weight of BALB/cnu/nu mice after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 or BAL17CNS cells.
NK cells contribute to the i.c. immune response against BAL17CNS/Cas9 lymphoma cells in BALB/cnu/nu mice
The clinical course of BALB/cnu/nu mice transplanted with either BAL17CNS or BAL17CNS/Cas9 cells was reflected by i.c. development and growth of PCNSL (Figs. 6 and 7). Surprisingly, BALB/cnu/nu mice that had received BAL17CNS/Cas9 cells did not develop PCNSL (Fig. 7A, C, and E). In contrast, BAL17CNS cells had induced bulky lymphoma involving large areas of the supratentorial brain, including the hypothalamus, the caudate-putamen, the internal capsule, as well as the frontal and parietal cortex in BALB/cnu/nu mice without inflammation (Fig. 7B, D, and F; Supplementary Fig. S3).
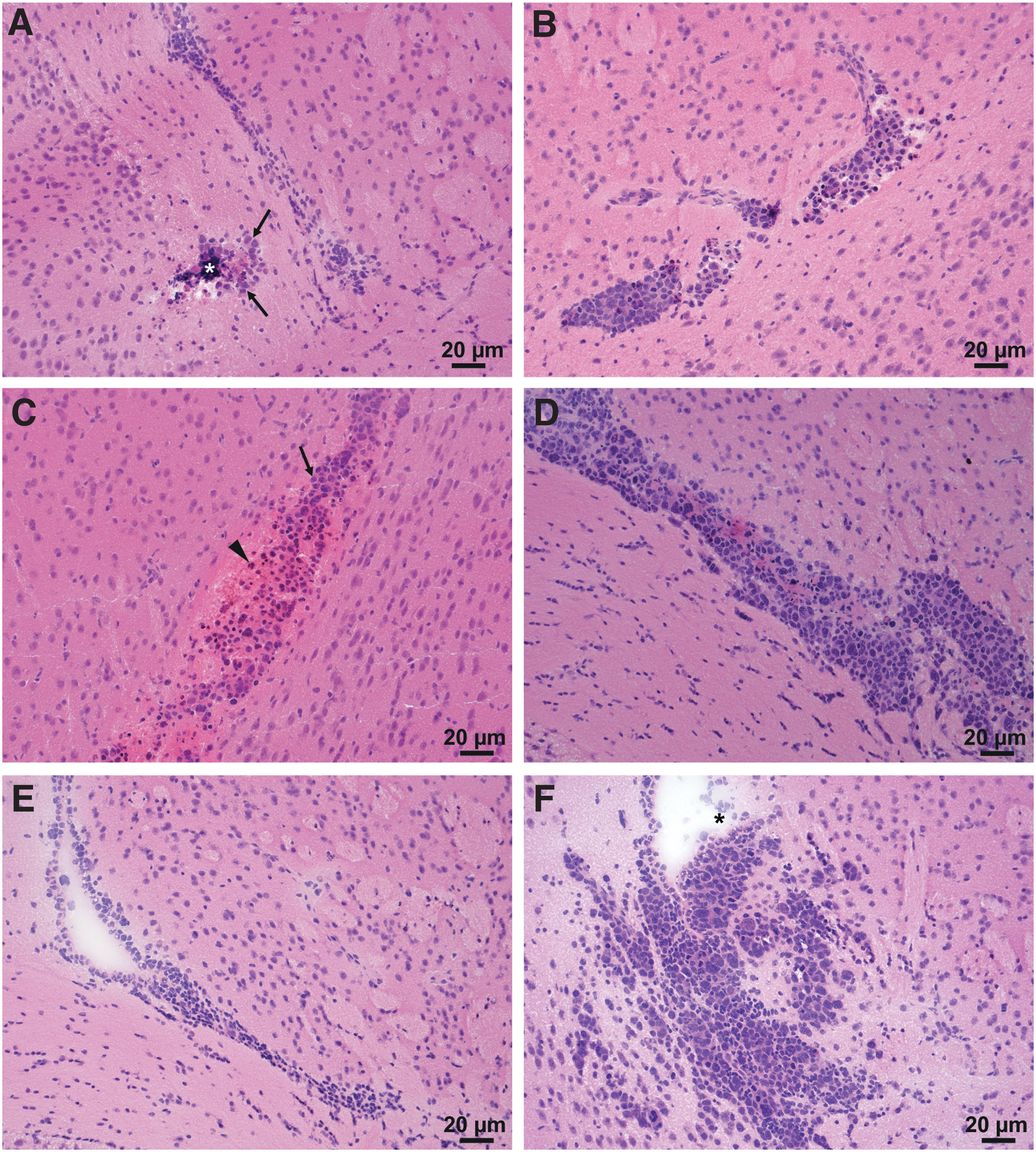
BAL17CNS/Cas9 lymphoma cells fail to induce PCNSL in BALB/cnu/nu mice.
As early as day 1 p.i., inflammation was initiated in the brains of BALB/cnu/nu mice that had received i.c. BAL17CNS/Cas9 cells, increasing thereafter (Fig. 7A, C, and E). Inflammatory infiltrates mainly involved the lateral ventricle and the neighboring subependymal brain parenchyma (Fig. 7A, C, and E). CD335+ NK cells (Fig. 8A), F4/80+ macrophages/activated microglia (Fig. 8B), and CD11c+ myeloid cells (Fig. 8C) were associated with single tumor cells or small clusters of BAL17CNS/Cas9 cells (Fig. 8A–D). In addition to microglia, cerebral endothelial cells were activated and had also upregulated the MHC class I antigen (Fig. 8D). Perforin was secreted close to BAL17CNS/Cas9 lymphoma cells (Fig. 9A–C), resulting in the death of tumor cells.

Phenotypic composition of the inflammatory infiltrates in the brains of BALB/cnu/nu mice at day 3 after i.c. transplantation of 5 × 105 BAL17CNS/Cas9 cells.
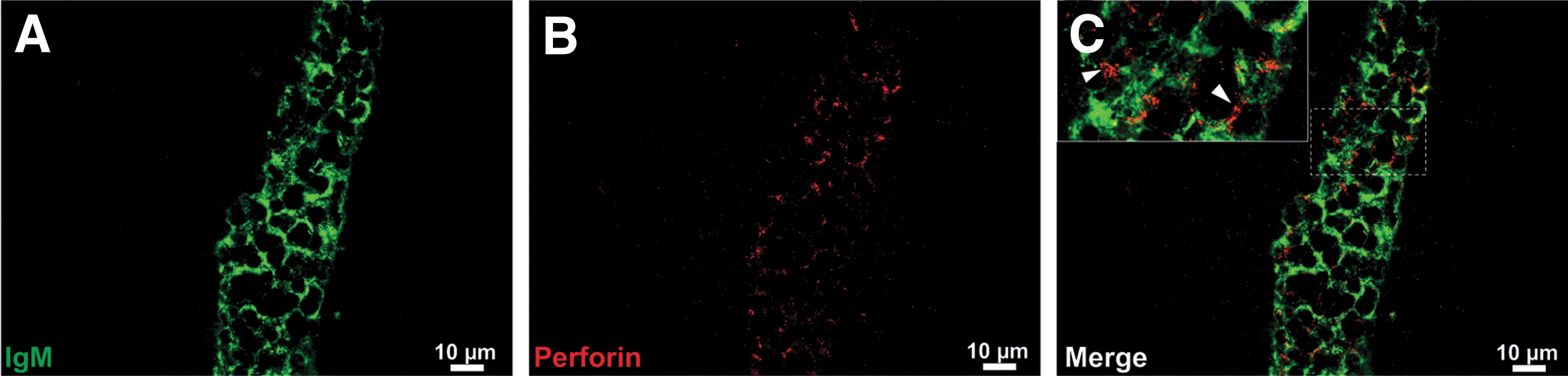
Perforin-mediated killing of BAL17CNS/Cas9 cells in the brains of BALB/cnu/nu mice at day 3 p.i.
These experiments demonstrate that NK cells also contributed to the i.c. immune response against Cas9-expressing BAL17CNS cells and induced BAL17CNS/Cas9 lymphoma cell death via secretion of perforin.
BALB/c mice produce serum anti-Cas9 antibodies upon i.c. transplantation of 5 × 105 BAL17CNS/Cas9 lymphoma cells
In BALB/c mice having been transplanted with BAL17CNS cells, serum reactivity against Cas9 was determined by enzyme-linked immunosorbent assay (ELISA); the mean + 3 × SD was defined as a cut-off (Fig. 10A). Compared with this threshold, 50% (3/6) of BALB/c mice that had received i.c. BAL17CNS/Cas9 cells had detectable serum anti-Cas9 antibodies at day 8 p.i. (Fig. 10A). At day 8 p.i., BALB/Cnu/nu mice transplanted with BAL17CNS/Cas9 cells showed an anti-Cas9 serum reactivity similar to that of BALB/Cnu/nu mice transplanted with BAL17CNS cells that was defined as cut-off (mean +3 × SD; Fig. 10B).

Production of serum anti-Cas9 antibodies in BALB/c, but not BALB/cnu/nu mice at day 8 p.i. having been i.c. transplanted with 5 × 105 BAL17CNS/Cas9 lymphoma cells.
Thus, i.c. transplantation of BAL17CNS/Cas lymphoma cells induced serum anti-Cas9 antibodies in immunocompetent BALB/c mice but—as expected—not in athymic nude BALB/cnu/nu mice.
Discussion
This study demonstrates that BAL17CNS lymphoma cells with stable expression of Cas9 (BAL17CNS/Cas9) transplanted into the brain of BALB/c mice fail to induce PCNSL. Instead, they induced a prominent i.c. host immune reaction against Cas9 with CD8 T cells perforin-mediated destruction of BAL17CNS/Cas9 cells. In the absence of T cells, a vigorous NK cell response accounted for the perforin-induced killing of BAL17CNS/Cas9 cells in the brain.
To the best of our knowledge, an immune response against Cas9 in the immune-privileged CNS has not yet been described. In our PCNSL model using BAL17CNS/Cas9 cells, the adaptive immune response was mediated by strongly activated CD8 T cells. The kinetics of the i.c. CD8 T cell response with establishment at day 8 p.i. and a further increase up to day 16 p.i. is in line with that described for mice immunized with SaCas9 in which hepatocytes were targeted by adenovirus-associated CRISPR-Cas; in the latter, a hepatic CTL response completely eliminated genome-edited cells.18,19
In general, CD8 T cells can be stimulated via cross-presentation or via peptides presented by MHC class I. Interestingly, in healthy human donors, two immunodominant SpCas9 T cell epitopes for HLA-A*02:01 were recently identified. 14 In mice, Cas9 peptides associated with MHC class I molecules that activate CD8 T cells have—to the best of our knowledge—not yet been described. Their identification will provide an important clue to elucidate the spectrum of immune responses to Cas9 orthologs.
Regarding the in vivo consequences of Cas9 expression in our PCNSL model in syngeneic immunocompetent BALB/c mice, we identified the cytotoxic molecule perforin secreted by effector CD8 T cells as functionally relevant mechanism for the lysis of Cas9-expressing lymphoma cells. Polarized expression of perforin and granzyme B toward targets that subsequently undergo cell death is indicative of cell death induction.20,21
So far, perforin-induced CD8 T cell-mediated destruction has not been reported in the host immune response against Cas9. In some, but not all B cell lymphoma entities an immune escape mechanism with bcl-2-mediated inhibition of perforin-dependent cytotoxicity has been described.22–25 The strong bcl-2 expression by BAL17CNS/Cas9 cells (Supplementary Fig. S4A, B) indicates that perforin-induced death of BAL17CNS/Cas9 cells was not due to a lack of this anti-apoptotic protein.
Noticeably, the immune reaction was aggressive enough to induce prominent necrosis of the brain tissue harboring Cas9-expressing lymphoma cells and the adjacent parenchyma, which was absent from the brain of BALB/c mice transplanted with BAL17CNS cells. These data add to previous in vitro observations that SpCas9-specific effector T cells lyse Cas9-transfected lymphoblastoid cell line in a dose-dependent manner. 15
In healthy humans, CD8 T cell immunity was interpreted to likely reflect a memory response; however, even in the absence of a pre-existing immune response, the expression of Cas9 in naive individuals may trigger a T cell response. 14 In our experimental system, mouse were housed under SPF conditions that preclude Sp species; however, one cannot exclude contact with a cross-reactive Cas ortholog since CRISPR systems exist across a wide range of bacterial species (for a review see Katti et al 5 ). The perforin+CD62L+ effector memory phenotype of the i.c. CD8 T cells in our model extends in vitro observations by Wagner et al, 15 who identified most human SpCAs9 reactive CD8 T cells as terminally differentiated effector memory cells.
Recently, in humans, 22 SaCas9 peptides have been identified that are presented by MCH class II molecules and elicit CD4 T cell proliferation. 26 However, their precise role in the anti-Cas9 immune response has not yet been defined. Our observations of only few CD4 T cells with a resting, unstimulated phenotype do not support their crucial role in the i.c. immune response to Cas9. If among the few i.c. CD4 T cells were regulatory cells, they are functionally irrelevant as they did not impede CD8 T cell mediated killing of transfected cells. In line with these data, Li et al 18 identified Cas9-reactive regulatory CD4 T cells in the liver of Cas9-targeted hepatocytes, which did not prevent the killing of Cas9-expressing target cells. 18
In our model, Cas9 was strongly and permanently expressed in BAL17CNS cells. Interestingly, when Cas9 levels were reduced in renal carcinoma cells that failed to grow subcutaneously (s.c.) due to Cas9-transfection, growth was similar to the parental cell line, 27 suggesting a dose-effect of Cas9 expression on the induction of a host immune response.
Collectively, there is cumulative evidence for an anti-Cas9 directed T cell response, although one cannot formally exclude the possibility that the transfection process yielded an immunogenic neo-antigen that may contribute to stimulation of the immune response. In this regard, the recent identification of an immune reaction not confined to Cas9 epitopes but also to other vector components is of note. 28
The absence of CD8 T cells was not sufficient to allow PCNSL establishment by BAL17CNS/Cas9 cells as evidenced in BALB/cnu/nu mice, in which genome-edited lymphoma cells also failed to establish PCNSL. This situation is quite different from a model of s.c. transplanted Cas9-expressing pancreatic tumor cells in which CD4 and CD8 T cell depletion enabled tumor growth. 29
In our model, the rapid recruitment of NK cells to i.c. BAL17CNS/Cas9 lymphoma cells was likely fostered by antigenic peptides deliberated from disintegrating tumor cells that stimulate toll like receptor-mediated signaling pathways and, thereby, create a pro-inflammatory microenvironment. The remarkably strong activation of resident brain cells including microglia and blood vessel endothelial cells also contributed to the pro-inflammatory milieu. The interaction of NK cells and CD11c+ myeloid cells associated with i.c. BAL17CNS/Cas9 l cells may have induced their reciprocal activation.
This cross-talk has likely fostered NK cell effector function that ultimately resulted in the efficient lysis of BAL17CNS/Cas9 lymphoma cells in the brain of BALB/cnu/nu mice. Interestingly, a relatively low number of NK cells, assessed by immunohistochemistry of CD335, an antigen uniformly expressed by rodent and human NK cells,30–32 was sufficient for the prominent and widespread secretion of death-inducing perforin. In immunocompetent BALB/c mice, NK cells may also play an important role in support of the i.c. adaptive immune response.
In addition to NK cells and CD8 T cells, B cells also contributed to the anti-Cas9 immune response in PCNSL. As early as day 8 p.i., when the T cell response was established, a humoral immune response was initiated as evidenced by the presence of serum anti-Cas9 antibodies in 50% (3/6) of immunocompetent BALB/c mice, which—as expected—were absent from BALB/cnu/nu mice. The production of specific anti-Cas9 antibodies further supports the Cas9-specificity of the CD8 T cell reaction.
In immunocompetent mice s.c. implanted with Cas9-expressing pancreatic carcinoma cells, anti-Cas9 serum antibodies were present in the majority of mice at day 13 of the immune response. 29 Nevertheless, the potentially supportive contribution of antibodies in the immune response against Cas9 still remains to be elucidated.
Conclusion
Thus, in conclusion, the data of our study indicate that caution is warranted when using Cas9-transfection in the CNS, for example, for therapeutic purposes, as Cas9 induces a prominent i.c. immune response to which activated resident brain cells as well as NK and CD8 T cells recruited to the brain contribute. Ultimately, rejection of Cas9-expressing BAL17CNS/Cas9 lymphoma cells mediated by aggressive NK and CD8 T cells that secrete perforin lysed their target cells in the brain. Regarding safety and efficacy of CRISPR-based therapy, the recent engineering of Cas9 variants with reduced immunogenicity 14 may offer an attractive tool to prevent such serious side effects of the i.c. immune response.
Footnotes
Acknowledgments
The authors thank Mariana Carstov and Elena Fischer for their expert technical assistance.
Authors' Contributions
Conceptualization: M.D., M.M.-R., R.S., M.S.-R., and A.B. Methodology: M.D., M.M.-R., M.S.-R., A.B., S.S., and C.W. Validation: M.D., M.M.-R., M.S.-R., A.B., S.S., and C.W. Formal analysis: M.D., M.M.-R., R.S., M.S.-R., A.B., and S.S. Investigation: M.D., M.M.-R., M.S.-R., A.B., S.S., and C.W. Data discussion and interpretation: M.D., M.M.-R., M.S.-R., A.B., S.S., R.S., and C.W. Writing: M.D. Visualization: M.D., M.M.-R., M.S.-R., A.B., S.S., and C.W. Funding acquisition: M.D., R.S., and M.M.-R.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a grant from the Deutsche Krebshilfe (70112052 and 70112053) to M.D. and R.S. and by grants from the Wilhelm-Sander Stiftung (2011.092.2, 2020.149.1) to M.D. and M.M.-R.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.