
Abstract

A new study in Science led by Ailong Ke and Stan Brouns combines cryoelectron microscopy and biochemical analyses to show how “Craspase”—a Type III-E CRISPR complex bound to a caspase—couples RNA recognition with protease activation.
Type III CRISPR-Cas systems are reputed to be the most complex of the six CRISPR types, and now with the discovery of Type III-E, they are also proving to be the most versatile. Categorized as Class I, Type III systems launch a sophisticated and multilayered immune response to foreign RNA using effector complexes typically composed of multiple subunits. In addition to the core genes that code for the effector complex, Type III loci have been found to harbor an assortment of accessory genes that encode a variety of enzymes, including nucleases and proteases.1,2
Although accessory nucleases now have well-established mechanisms for recruitment and activation during the Type III immune response, whether and how a protease can become involved in immunity has remained elusive. Solving this puzzle, two teams led by Ailong Ke (Cornell University) and Stan Brouns (Delft University of Technology) combined their efforts to reveal the mechanism by which a Type III-E CRISPR-Cas system in complex with a protease triggers protein cleavage upon RNA recognition.3,4
Type III systems are divided into six subtypes (A–F) that have slightly different cas gene compositions (Table 1). Of these, Type III-E systems are among the newest additions to the family. 5 They were bioinformatically identified in metagenomic data sets as resembling Type III systems but have several features that set them apart.
Encodes the signature subunit for Type III systems.
Encodes repeat-associated mysterious proteins.
Encodes the small subunit in Type III systems.
Subtypes III-B and III-C are distinguished by the presence/absence of cas6, which is not a part of the effector complex.
First, although other Type III systems rely upon multisubunit effector complexes, Type III-E systems employ a single-subunit effector composed of a fusion of several domains. These include multiple domains homologous to Cas7, which belongs to the repeat-associated mysterious protein (RAMP) superfamily, as well as a domain resembling the small subunit Cas11 (Fig. 1). Accordingly, Type III-E effector complexes have been referred to as Cas7–11.6,7 They also go by the nickname “giant” RAMP (gRAMP).3–5
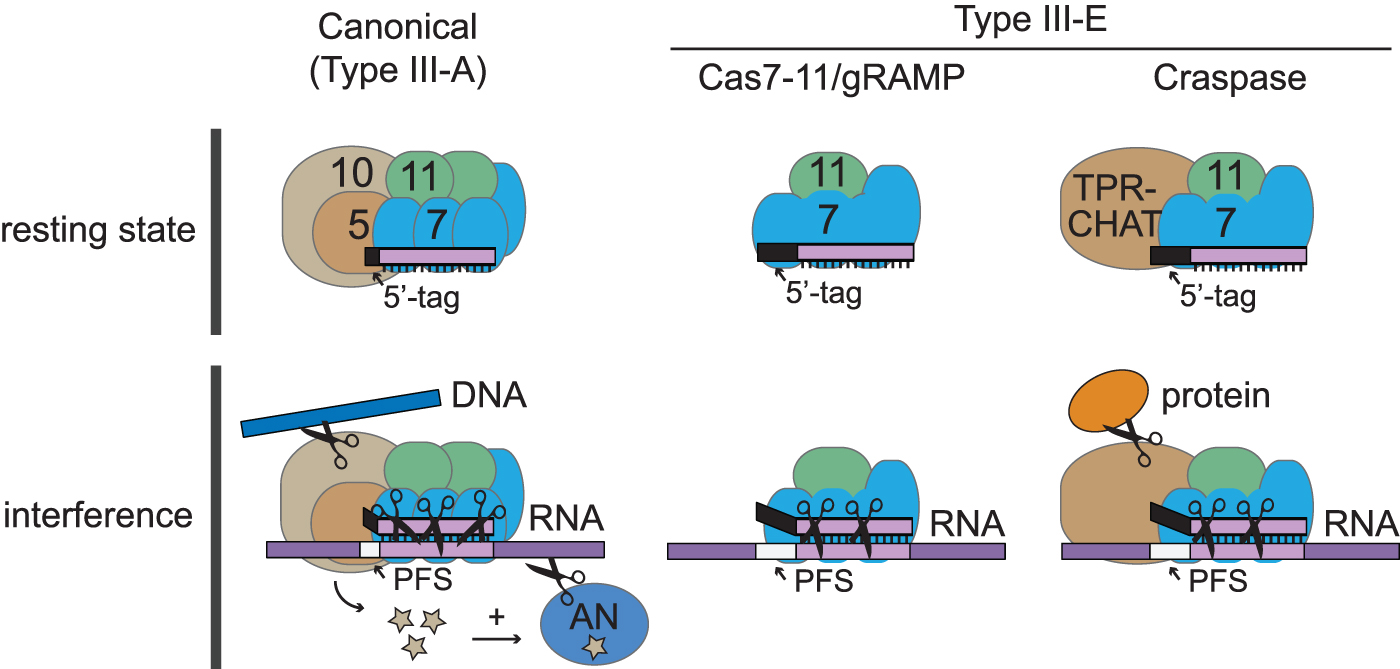
Comparison of effector complexes from a canonical Type III CRISPR-Cas system and the atypical Type III-E system. Colored ovals represent protein subunits and stars represent cyclic oligoadenylate signaling molecules. AN, accessory nuclease.
Second, as another distinctive feature, gRAMP lacks domains associated with the Type III signature subunit Cas10. In canonical systems, Cas10 confers DNase activity upon foreign RNA recognition, and some homologs also generate cyclic-oligoadenylate molecules, second messengers that bind and activate a variety of accessory nucleases that are not a part of the complex but typically encoded nearby. 8 Instead of accessory nucleases, many Type III-E loci were found to encode a putative accessory protease with tetratricopeptide repeats (TPRs) and a Caspase HetF Associated with Tprs (CHAT) domain, which is typically found in caspases (i.e., proteases involved in programmed cell death). 5 In light of the many idiosyncrasies of Type III-E systems, the first challenge in their functional characterization has been to determine whether and how the gRAMP alone senses and destroys nucleic acids.
Taking on this challenge, the Brouns group had previously characterized biochemically the gRAMP encoded in Candidatus Scalindua brodae (Sb-gRAMP). 3 Similar to other Type III systems, Sb-gRAMP was found to copurify with processed crRNAs and cleave complementary RNA in a metal-dependent manner. In addition, RNA cleavage was found to persist in the presence of complementarity between the protospacer flanking sequence (PFS) and the repeat-derived sequence on the 5′-end of the crRNA (i.e., 5′-tag). In canonical Type III systems, mismatches between the PFS and 5′-tag are not required for RNA cleavage but are essential to signal the detection of “nonself” nucleic acids and activate Cas10 catalysis.
As Type III-E systems are devoid of domains resembling Cas10 and corresponding accessory nucleases, the investigators tested for physical interactions between Sb-gRAMP and the TPR-CHAT protease encoded nearby. Indeed, coexpression of Sb-gRAMP and TPR-CHAT resulted in the production of a stable 1:1 complex. This complex was given the name “Craspase” (CRISPR-guided caspase), and in light of the parallels with other Type III systems, the authors proposed a model for Craspase-mediated immunity in which target RNA recognition allosterically triggers protease activity in the complex.
On the Move
In their seminal follow-up study in Science, Ke, Brouns, and colleagues proved the validity of this model by generating a series of cryoelectron microscopy snapshots of gRAMP and Craspase in different functional states, morphing the images into movies to track the structural changes, and testing mechanistic predictions with functional assays. 4 The investigators first solved the structure of Sb-gRAMP on its own in three different states—resting (i.e., unbound), target RNA-bound, and post-RNA cleavage.
Within the RNA-bound state, two types of targets were investigated—an RNA with a PFS that matches the crRNA 5′-tag and an RNA with a mismatching PFS to mimic nonself RNA detection. These structures revealed the presence of a long flexible loop that, upon target RNA binding, propagates conformational changes within gRAMP to align the ribonuclease catalytic residues. This flexible linker, termed the “gating loop,” causes similar structural changes in the presence of both target RNAs (PFS matched and PFS mismatched), suggesting that rather than detecting self versus nonself, the gating loop may play a role in recognizing appropriate targets.
To test this, the investigators mutated the gating loop, purified the wild-type and mutant gRAMP complexes, and performed functional assays. Unlike wild type, the mutant gRAMP was found to promiscuously bind and cleave an RNA substrate with only nine nucleotides of complementarity with the crRNA. These observations confirmed that the gRAMP gating loop is critical for sensing the presence of a bona fide target (∼16–18 nucleotides in length) and licensing RNA cleavage.
The investigators next revealed the mechanism of RNA-triggered protease activity by solving three different Craspase structures—unbound and bound to two RNA targets with PFS sequences that match and mismatch the 5′-tag, respectively. Strikingly, TPR-CHAT was found to occupy a position in the Craspase complex proximal to the crRNA 5′-tag, analogous to the position of Cas10 in canonical Type III effector complexes (Fig. 1). The authors noticed that in the unbound complex, the active site residues of the CHAT domain are positioned too far apart to catalyze proteolysis. However, in the target-bound Craspase complex, dramatic conformational changes bring the active site residues in proximity. These changes are conveyed by a long “switch helix” connecting the TPR and CHAT domains.
Notably, the structural changes are more pronounced when Craspase is bound to the PFS mismatched RNA target, suggesting that the protease only becomes active when bound to nonself RNA. To corroborate these predictions, the authors went on to identify the natural target of proteolysis—Csx30 encoded in the CRISPR locus—and reconstitute its Craspase-mediated cleavage. Indeed, Csx30 cleavage was observed only in the presence of a PFS mismatched/nonself RNA target. Although the function of Csx30 remains to be determined, the authors speculate that its cleavage perpetuates an immune response that may lead to cell dormancy/death.
Beyond expanding our understanding of the mechanistic diversity of Type III CRISPR-Cas systems, this and other studies are paving the way for a variety of new technologies. Already, recent studies have characterized Cas7-11 from Desulfonema ishimotonii (DiCas7-11) and repurposed it as an RNA knockdown/editing tool in eukaryotic cells.6,7 It bears mentioning that DiCas7-11 is devoid of nonspecific nuclease activity and nontoxic to mammalian cells, potentially making it more attractive than other CRISPR-based RNA-targeting tools.
Undoubtedly, Craspase's remarkable capacity to catalyze RNA-guided protein cleavage now opens up a new world of possibilities in bioengineering and medicine.