
Abstract

Ten years ago, Weizong Bao and Jerzy Jurka identified bacterial TnpB-like proteins (Fanzor [Fz]1 and Fz2) in diverse eukaryotic transposable elements and large double-stranded DNA (dsDNA) viruses infecting eukaryotes. 1 This finding triggered considerable excitement in the field, but that soon faded as the function of Fz proteins remained uncertain. The recent discovery that TnpB is an RNA-guided DNA nuclease and a precursor of the Cas12 family of type V CRISPR-Cas effectors2–4 renewed interest in Fzs, triggering speculation that they might serve as functional RNA-programmable nucleases in eukaryotic cells. Writing in Nature, researchers in Feng Zhang's laboratory demonstrate that eukaryotic Fz proteins are indeed RNA-guided DNA endonucleases. 5
By combining advanced structural and sequence profile mining, Saito et al. first identified more than 600 variants of curated Fz proteins encoded in eukaryotes, mostly in fungi, protists, plants, and eukaryotic viruses (Fig. 1A). 5 In agreement with the original study, 1 phylogenetic analysis revealed two main branches of Fz proteins, Fz1 and Fz2, which likely originated from diverse TnpB proteins that were horizontally transferred to and between eukaryotic hosts.
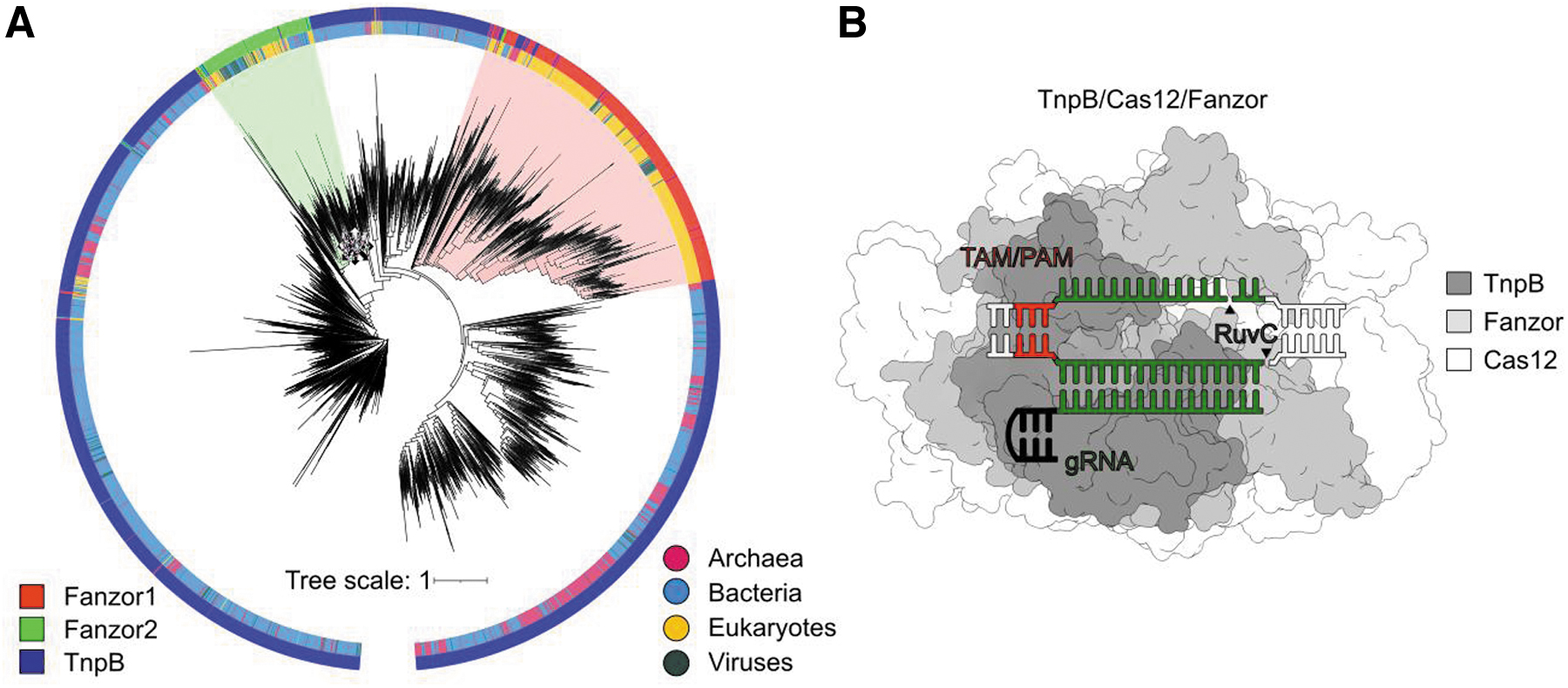
Identification of Fz proteins in a number of eukaryotic viruses, which are potential vectors for horizontal gene transfer, supports this hypothesis. Occasionally, some Fz variants reside on the same branches with TnpB in the phylogenetic tree and are identified in eukaryotic hosts that have close interactions with bacteria, suggesting actively ongoing TnpB transfer to eukaryotes. Overall, the phylogenetic analysis of Fz and TnpB proteins indicates that Fz arose in eukaryotes through the horizontal transfer of TnpB from prokaryotes and Fz gene transfer between eukaryotes.
Subsequent biochemical analysis revealed that, similar to prokaryotic TnpBs,4,6,7 Fz proteins associate with a guide RNA mostly derived from the flanking DNA sequence downstream of Fz open reading frame DNA sequence, followed by a short stretch of the downstream flanking sequence. 5 Guided by RNA, Fz effectors recognize and cleave DNA targets near the target/transposon-adjacent motif, named TAM, which was previously identified for TnpB3,4 and is equivalent to the protospacer-adjacent motif recognized by Cas12.6,8–10
Cryo-electron microscopy (cryo-EM) structure of Fz1 variant from Spizellomyces punctatus with guide RNA and target DNA uncovered typical bi-lobed complex architecture characteristic for TnpB and Cas12 effectors (Fig. 1B).11–15 These results indicate that Fz effectors function in eukaryotes using the structural and molecular mechanism of target DNA recognition and cleavage similar to bacterial TnpBs and related Cas12 effector proteins of CRISPR-Cas systems. However, their biological function remains to be established.
Prokaryotic TnpB proteins that are precursors of Fzs are associated with widespread IS200/IS605 mobile genetic elements (MGEs) in prokaryotes.1,5 Typically, IS200/IS605 family MGEs encode tnpA and tnpB genes flanked by left-end and right-end elements. Although it has been well established that TnpA is a transposase responsible for transposon mobilization, through a single-strand circular DNA intermediate utilizing a “peel-and-paste” mechanism, 16 the possible role of TnpB in transposition has only emerged recently.
The discovery that TnpB functions as an RNA-guided DNA endonuclease that cuts dsDNA targets in the vicinity of the TAM sequence, which coincides with the sequence required for IS200/IS605 insertion/excision, indicates that the donor site generated after transposon excision from the lagging DNA strand becomes the target for TnpB. 3 Thus, after TnpA-mediated transposon excision, TnpB should be able to cleave the newly formed donor site promoting recombination that will reinstate the transposon copy at the position where it was excised, using a sister chromatid containing a transposon as a template. This “peel–paste–copy” mechanism, which is supported by preliminary data,3,17 would prevent transposon loss and could explain the increase of the transposon copy number observed in some strains.
Fz function
The first demonstration of the RNA-guided DNA cleavage activity of eukaryotic TnpB-like Fz proteins raises the big question of their biological function. The differences between eukaryotic and prokaryotic cells—notably the presence or absence of the nuclear envelope—imply that Fz effectors had to acquire additional functionalities to adapt to the eukaryotic cellular environment. Indeed, emerging evidence suggests that some of the Fz effectors contain nuclear localization sequences and introns, enabling Fz function in eukaryotic cells. 18
The localization of Fz effectors in the vicinity of other transposons, including Helitron, Mariner, IS4-like, and the presence of multiple copies of Fzs, suggests that their function could be similar to TnpB in the transposition of prokaryotic MGEs.1,5,18 Standalone Fz effectors occasionally present in the genomes can be coupled to transposases acting in trans or could perform additional functions that still need to be determined, suggesting that further Fz studies hold great potential for some exciting new discoveries.
The key discovery provided by the Zhang laboratory is the demonstration that Fzs can function as programmable genome editors 5 similar to Cas12 and TnpB effectors.3,4,8,19–21 Saito et al. show that the engineered SpuFz1 variant introduced ∼20% of insertions/deletions at the targeted genomic site in human cells, demonstrating the potential of eukaryotic RNA-guided DNA targeting Fz effectors as potential genome editing tools.
The noble pursuit of curiosity-driven research has yet again showcased its significance, yielding invaluable discoveries and nature-inspired tools that hold immense potential for shaping the landscape of molecular biology in the near future.
Footnotes
Disclaimer
V.S. is a chairman and co-founder of Caszyme.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
T.K. is supported by the Research Council of Lithuania (S-MIP-21-8).