
Abstract
Background:
Pathological displacement of teeth caused by periodontitis-related bone loss in patients with diabetes is often corrected with orthodontic treatments. However, recovery from orthodontic therapy is often delayed for unclear reasons. This study explored effects of streptozotocin-induced diabetes in rats on protein expression involved in remodeling of the periodontal ligament (PDL) and alveolar bone during orthodontic tooth movement.
Methods:
Forty-eight Sprague-Dawley rats were randomly divided into two experimental groups: “normal” and “diabetes” (n = 24 each). Diabetes was induced by a single dose of streptozotocin (65 mg/kg). Animals were euthanized at 3, 7, and 14 days after orthodontic induction. Changes in expression of collagen type I (Col-I), matrix metalloproteinase type 1 (MMP-1), and tissue inhibitor of MMP-1 (TIMP-1) were measured immunohistochemically in the pressure side. Col-I and collagen type III (Col-III) fibers were assessed by picrosirius red staining in the tension side. Osteoclasts were observed on the surface of the alveolar bone.
Results:
Diabetes increased expression of MMP-1 and Col-III and decreased expression of Col-I in PDL. After the orthodontic induction, osteoclast action was delayed, and higher Col-III/Col-I and MMP-1/TIMP-1 ratios persisted in the diabetes group compared with the normal group. The ratio of MMP-1/TIMP-1 in the diabetes group reached a peak on Day 7, whereas the ratio remained at near control levels in the normal group. The diabetes group appeared to have worse recovery from damage caused by orthodontic movement.
Conclusions:
Under mechanical forces, diabetes prolonged duration of degradation of PDL and remodeling of PDL and resorption of alveolar bone.
Introduction
Orthodontic tooth movement reflects dynamic, time-dependent changes to multiple components of the periodontium—periodontal ligament (PDL), neighboring bone, and gingiva—mediated through the expression of a variety of cytokines and proteases. 3,11,14 –20 Even appropriate orthodontic forces can cause mechanical injury to the PDL and alveolar bone resorption, 21 although this damage can resolve with remodeling of the PDL and alveolar bone.
Preclinical evidence indicates that numbers of osteoclasts increase under compression during orthodontic stress. 22 These large multinucleated cells originate by fusion of recently recruited preosteoclasts from the marrow (not local PDL cells). 17 PDL remodeling following application of force can be partly attributed to the modulation of fibroblast gene expression of proteases, such as matrix metalloproteinase (MMP) type 1 (MMP-1). 16 Animal models have also shown that orthodontic force significantly increases both levels of MMP-1 mRNA and the interstitial activity of MMP-1 in the gingiva, which can partly explain the clinically observed shrinkage and adaptation of the gingiva during tooth movement. 23
The balance of MMPs and tissue inhibitors of MMPs (TIMPs) is central to the stabilization of the extracellular matrix, with an MMP/TIMP imbalance associated with pathologic breakdown of the extracellular matrix in periodontitis. 24 –26 In diabetes, collagen is rapidly degraded by elevated MMP levels, 27,28 and fibroblast function is disturbed. 29 Because orthodontic treatment has been linked to remodeling of the collagens in the PDL, diabetes may complicate orthodontic treatment. Specific changes in the periodontium have been shown to be more pronounced after orthodontic movement in animal models of experimental diabetes as evidenced by slow osseous regeneration, weakening of the PDL, and microangiopathies in the gingival area. 30
Prior clinical research has largely focused on orthodontic treatment of patients with diabetes after controlling blood glucose levels. 12,13 However, previous reports have not explored the alteration of expression of collagens, MMPs, and TIMPs in the PDL after orthodontic treatment in uncontrolled diabetes. The present study explored the expression of collagen type I (Col-I) and type III (Col-III), MMP-1, and TIMP type 1 (TIMP-1) in the PDL of normal and rats with streptozotocin (STZ)-induced diabetes before and after application of orthodontic force.
Materials and Methods
Animals, experimental diabetes model, and orthodontic model
Forty-eight 7-week-old male Sprague-Dawley rats, weighing between 185 and 211 g were randomly divided into two experimental groups: “normal” and “diabetes” (n = 24 each group). Diabetes was induced in the diabetes group by a single intraperitoneal injection of 65 mg/kg of body weight STZ (Sigma Chemical Co., St. Louis, MO). Saline solution was injected in the normal group. Fasting blood glucose was evaluated prior to the injection of STZ and every day after 3 days post-injection using the Accu-Chek® Active monitoring system (Roche Diagnostics GmbH, Mannheim, Germany). Blood glucose was determined before orthodontic treatment and sacrifice of the study animals. STZ-injected rats were considered to have diabetes if fasting glucose levels were >300 mg/dL. Rats with diabetes and normal animals were treated identically except for injection of STZ or saline, respectively. Short-term diabetes was not expected to cause pathologic damage to periodontal tissues in rats with STZ-induced diabetes. 10 After injection, animals were housed in metabolic cages and fed standard rat chow and water ad libitum. Chlorhexidine-soaked tampons (Koutai®, South China Pharmaceutical Co. Ltd., Shenzhen, China) were used for biofilm plaque control once a week at the beginning of the study period and after STZ injection. After addition of orthodontic forces, oral care was carried out every 2–3 days. Oral care was focused on the area surrounding the experimental and control teeth with an emphasis on the supragingival dental cervical region.
Orthodontic appliances were placed in all rats in both experimental groups 8 weeks after STZ or saline solution injections. Orthodontic tooth movement in rats with STZ-induced diabetes and normal rats was achieved by inserting a standardized coil spring composed of nickel-titanium wire between the upper incisors and the left upper first molar. The orthodontic appliance was fixed to teeth with a 0.2-mm stainless steel wire. To prevent detachment of the appliance from the incisors' surface, we made a shallow groove approximately 0.5 mm from the gingiva of the upper incisors and fixed the stainless wire on the upper incisors. The force level of the coil spring after activation was approximately 0.5 N in keeping with previous reports indicating that application of 0.5 N does not damage periodontal tissues. 11,26,31 The PDL on the mesial side (“Area M”) of the mesial root of the molar was under pressure and the distal side (“Area D”) was under tension (Fig. 1). The right upper first molar served as the control to check for interanimal variability.
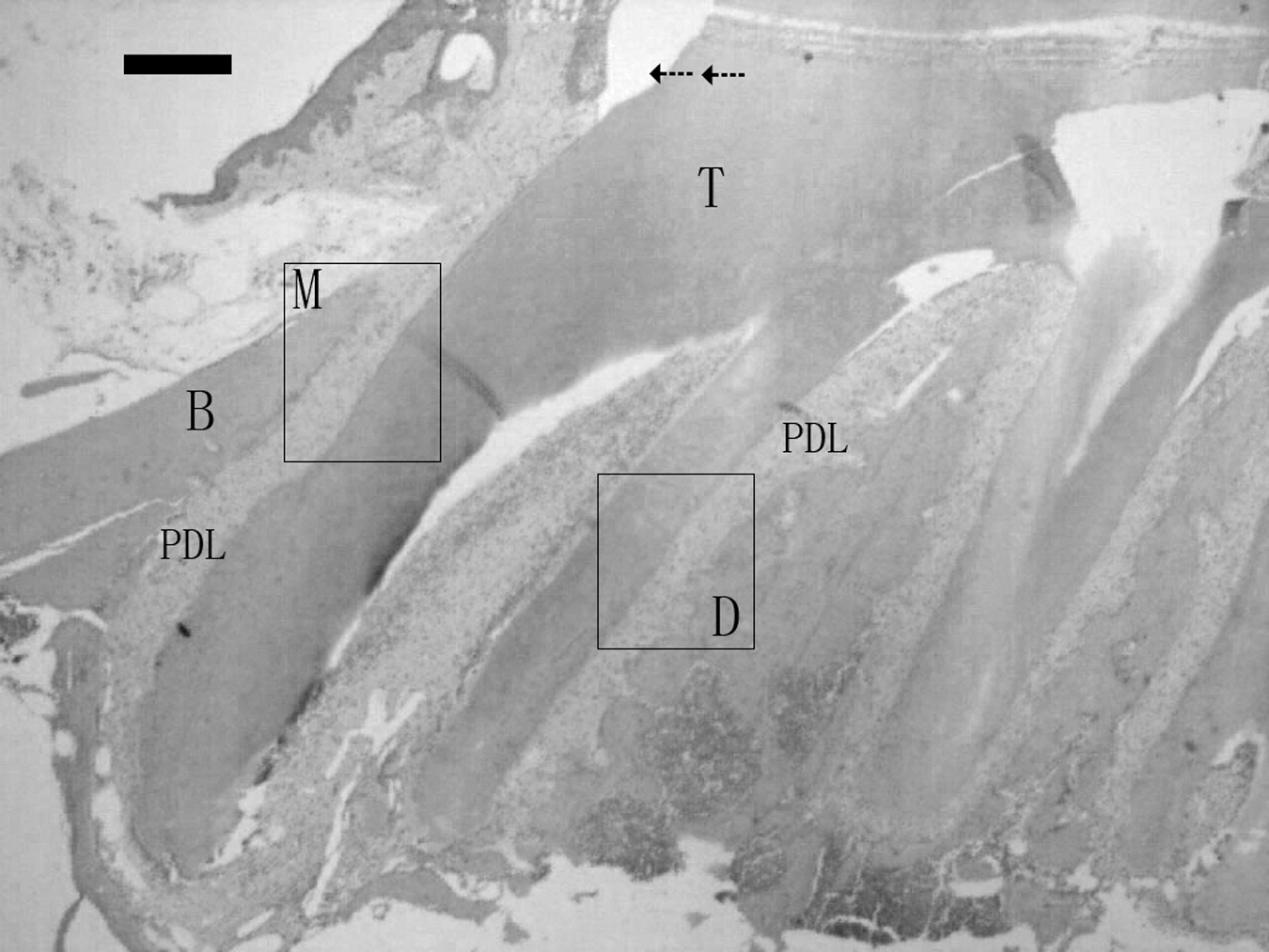
Histological appearance of alveolar bone (B) and PDL in normal rats. Direction of forces is indicated by arrows. Area M, PDL between the mesial surface of the proximal mesial root of the maxillary first molar root and B; Area D, PDL between the distal surface of the proximal mesial root of the maxillary first molar and B. T, teeth.
Rats were anesthetized, perfused intracardially with 4% paraformaldehyde, and decapitated 3, 7, and 14 days after orthodontic induction. The maxillary jaw was fixed in the same fixing solution for 24 h at 4°C and decalcified for ≥4 weeks in EDTA at 4°C. The dimineralized tissues were dehydrated and embedded in paraffin. Tissue blocks were cut into 3-μm-thick mesiodistal serial sections and mounted on 3-aminopropyl-triethoxysilane-coated slides. The experimental protocol was reviewed and approved by the institutional animal ethics committee. All procedures adhered to the National Institutes of Health's Guide for the Care and Use of Laboratory Animals. 32
Histological and collagen fiber staining
Hematoxylin and eosin staining, picrosirius red staining, and circularly polarized light were used to examine the histochemistry of sections. 22,33 Collagen fibers were analyzed via polarizing microscopy after staining sections with picrosirius red for 30 min. In this technique, Col-I fibers appear red, and Col-III fibers appear green. The positive signal for Col-I and Col-III was calculated by image polarizing microscopy processing, and the ratio of Col-III/Col-I was determined. Matlab version 7.0 software 34,35 was used to calculate the proportion of positive signals in the PDL. Hematoxylin and eosin staining was used to identify osteoclasts. Each slide was examined by two independent blinded observers, and the average number of osteoclasts was recorded.
Immunohistochemical assay
Immunohistochemistry reagents were obtained from Wuhan Boster Biological Technology Ltd. (Wuhan, China). All sections were immersed in 3.0% H2O2 in methanol for 20 min to block endogenous peroxidase activity. Sections intended for analysis of MMP-1 and TIMP-1 were boiled in citrate buffer for 2 min and cooled naturally to indoor temperature. After three rinses in 0.01 M phosphate-buffered saline, the sections were incubated with blocking solution and then exposed to rabbit anti-Col-I, rabbit anti-MMP-1, or rabbit anti-TIMP-1 overnight at 4°C. The dilution ratio was 1:180, 1:100, and 1:80, respectively.
After a rinse with phosphate-buffered saline, sites with bound primary antibody were detected by incubating the sections with goat anti-rabbit secondary antibodies conjugated to horseradish peroxidase (1:600 dilution ratio) for 20 min. Sections were rinsed again and then immersed in a diaminobenzidine chromagen solution to visualize sites of antibody binding. Under microscopy, the diaminobenzidine reaction product generated good color formation discretely localized at horseradish peroxidase-labeled sites, providing high-resolution images of antigen distribution. Finally, sections were rinsed in distilled water, counterstained with hematoxylin solution, dehydrated, and coverslipped with a xylene-based mounting medium.
The positive signal of Col-I, MMP-1 and TIMP-1 was calculated by image processing (described above), and the ratio of MMP-1/TIMP-1 was determined. Immunoreactivity of Col-I, MMP-1, and TIMP-1 and the presence of Col-I and Col-III were confirmed by a blinded examiner. All the sections were examined by two individuals to select the positive signals, and the mean image elements ranges of positive signals were confirmed.
Measures of outcome
Pairwise comparison between the normal and diabetes groups was carried out for the right first molar (control) at baseline and the left first molar at Days 3, 7, and 14 of orthodontic induction for the following outcomes: osteoclast count and expression of Col-I, Col-III, MMP-1, and TIMP-1. Observation of osteoclasts was focused at the middle one-third of the PDL of the upper first molar close to the side of the alveolar bone in Area M. Area M was evaluated for Col-I, MMP-1, TIMP-1, and MMP-1/TIMP-1 ratio, and Area D was evaluated for Col-I, Col-III, and Col-III/Col-I ratio.
Statistical analysis
In general analysis, data were expressed as mean with SE. Repeated measurement analysis was performed for identifying the interaction between time and group effects through the generalized linear model (GLM) procedure. Furthermore, a Bonferroni post hoc test was used to evaluate the difference between groups at each time point, and within-group differences at a given time point were assessed through two-way analysis of variance test. All statistical assessments were under the significant level of 0.05 for overall comparison and of 0.05/3 = 0.0167 for post hoc comparison. Data were analyzed using SAS version 9.0 (SAS Institute Inc., Cary, NC).
Results
Histological findings
Application of orthodontic force stimulated osteoclastogenesis and formation of Howship's lacunae in both the normal and diabetes groups (Fig. 2). The number of osteoclasts in the normal and diabetes groups increased markedly after orthodontic induction and peaked at Day 7 (Fig. 3). There was a significant difference between the experimental groups at Day 14 (P < 0.05) reflecting a large decline in osteoclast number in the normal group versus a small decrease in the diabetes group.
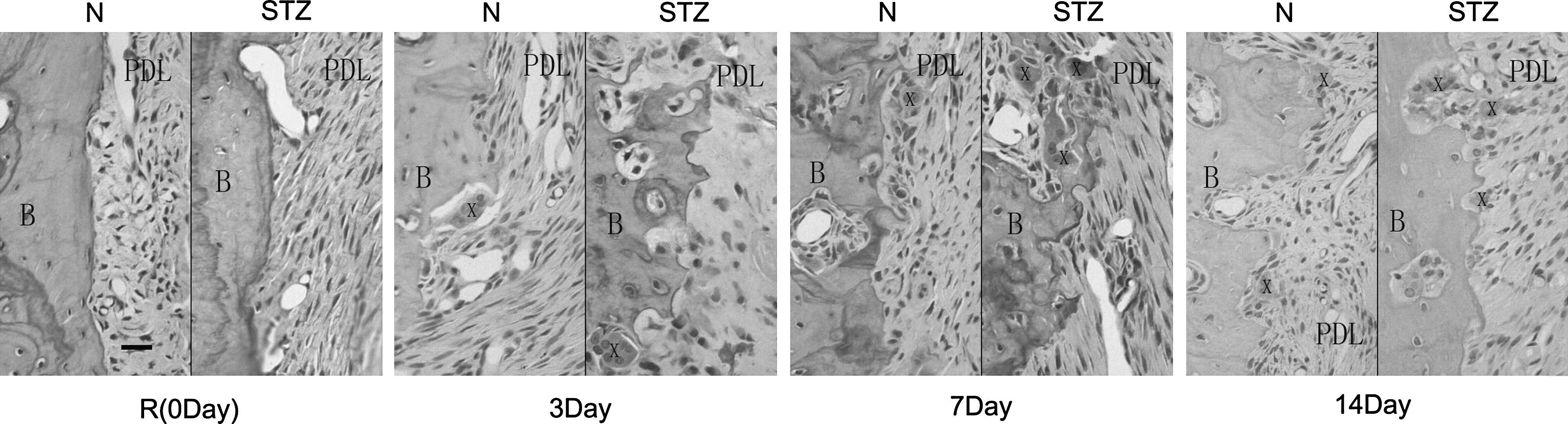
Histological appearance (hematoxylin and eosin-stained sections) of alveolar bone (B) and PDL in normal and diabetes groups under control conditions and Area M (pressure side) after orthodontic induction. X, osteoclasts.
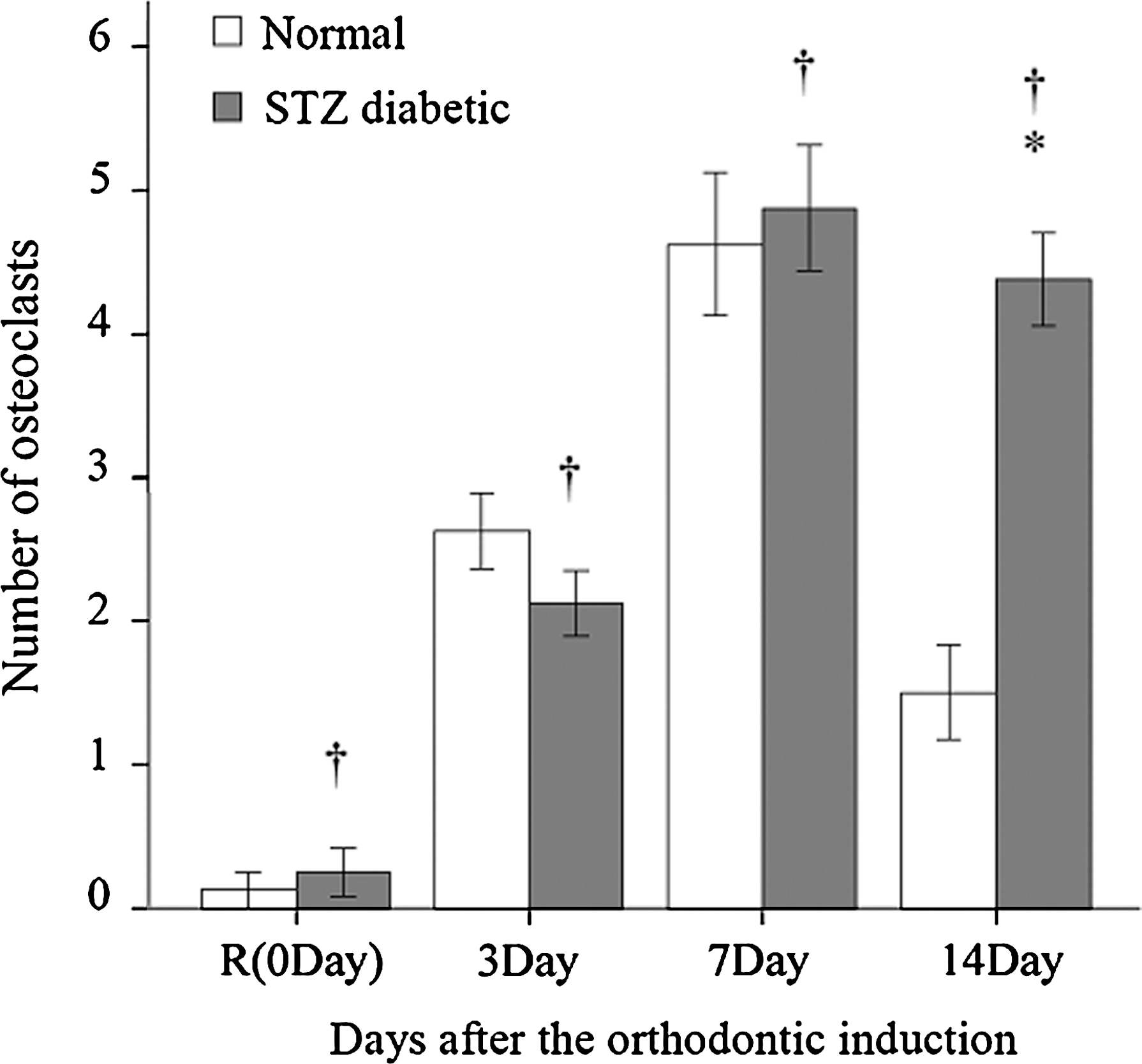
Osteoclast count in normal and diabetes groups with control treatment and after orthodontic induction. Data are mean ± SE values by group. †Before (0 Day) and after orthodontic induction within-group (normal and STZ-induced diabetes) means were significantly different (adjusted P < 0.05/3 through paired sample t test). *Normal and diabetes group means were significantly different (P < 0.05 through independent sample t test).
Immunoreactivity of Col-I, MMP-1, and TIMP-1 in Area M
In the control animals, experimental diabetes significantly decreased expression of Col-I (P < 0.05) and increased expression of MMP-1, with little difference in TIMP-1 between the normal and diabetes groups (Fig. 4).
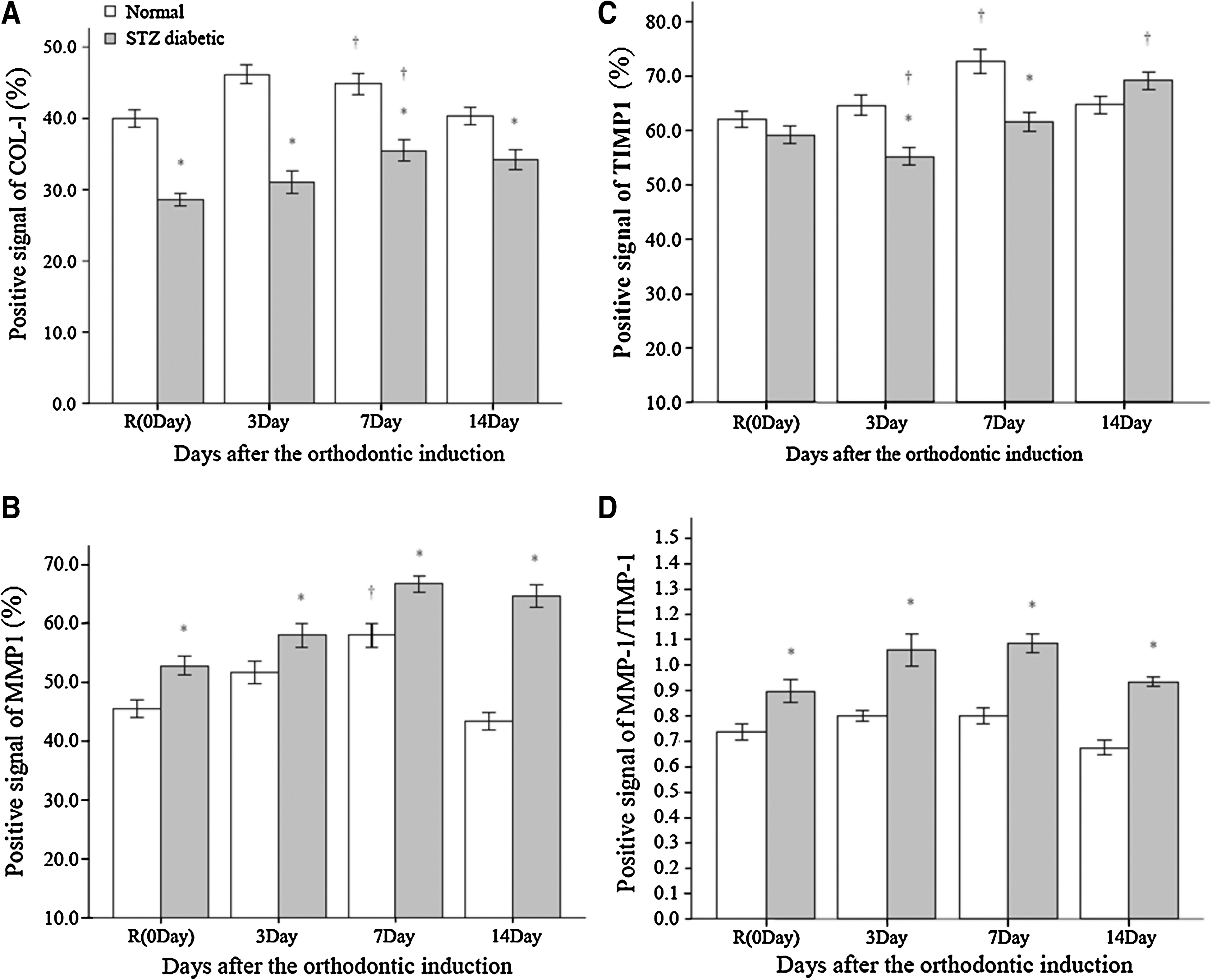
Expression of (
After orthodontic induction, the expression of Col-I in the PDL under pressure (Area M) in the normal group increased at Day 3 and plateaued at Day 7, after which Col-I declined to the level of controls by Day 14 (Fig. 4A). In the diabetes group, Col-I expression was virtually unchanged for the first 3 days, but increased at Day 7 before declining slightly at Day 14. Significant differences between the experimental groups were seen at all time points (all P < 0.05).
Expression of MMP-1 increased in both the normal and diabetes groups after orthodontic induction, reaching a peak at Day 7 (Fig. 4B). MMP-1 declined to pre-induction levels at Day 14 in the normal group but remained elevated in the diabetes group at Day 14 (P < 0.05). Significant differences between the experimental groups were seen at all time points (all P < 0.05). TIMP-1 levels were relatively stable in the normal group from Day 3 to 14, whereas a transient decrease in TIMP-1 inmmunoreactivity was observed in the diabetes group at Day 3 followed by a trend to higher values until Day 14 (Fig. 4C). There was a significant difference in expression of TIMP-1 between the normal and diabetes groups at Days 3 and 7 (P < 0.05).
With the control treatment, the ratio of MMP-1/TIMP-1 was slightly higher in rats with diabetes than in normal rats. During orthodontic induction, this ratio was little changed in the normal group, although with a slight decrease at Day 14 (Fig. 4D). At Day 3, the MMP-1/TIMP-1 ratio increased in the diabetes group (P < 0.05) and remained elevated at Day 7, but declined and approached pre-induction levels at Day 14.
Picrosirius red staining of collagen fibers in Area D
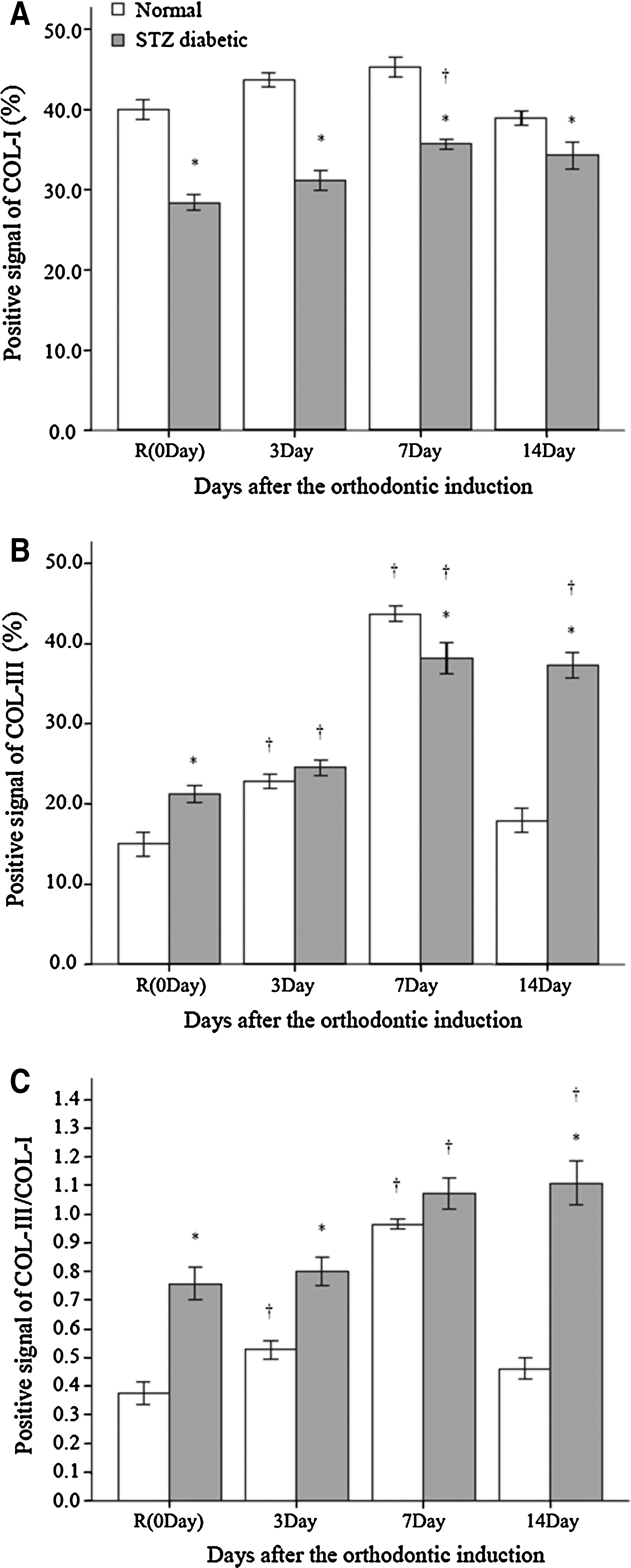
Figure 5 illustrates collagen bundles in picrosirius red-stained sections in normal and diabetes groups with control treatment and at Days 3, 7, and 14 after orthodontic induction. With the control treatment, expression of Col-I was significantly higher in normoglycemic rats than in rats with STZ-induced diabetes (P < 0.05) (Fig. 6A). When the PDL was under tension, Col-I and Col-III increased in both experimental groups until Day 7, with significantly different expressions of Col-I and Col-III in the experimental groups at all time points (Fig. 6A and B). Col-III decreased to pre-induction levels by Day 14 in the normal group, whereas Col-III remained near peak levels at Day 14 in the diabetes group.
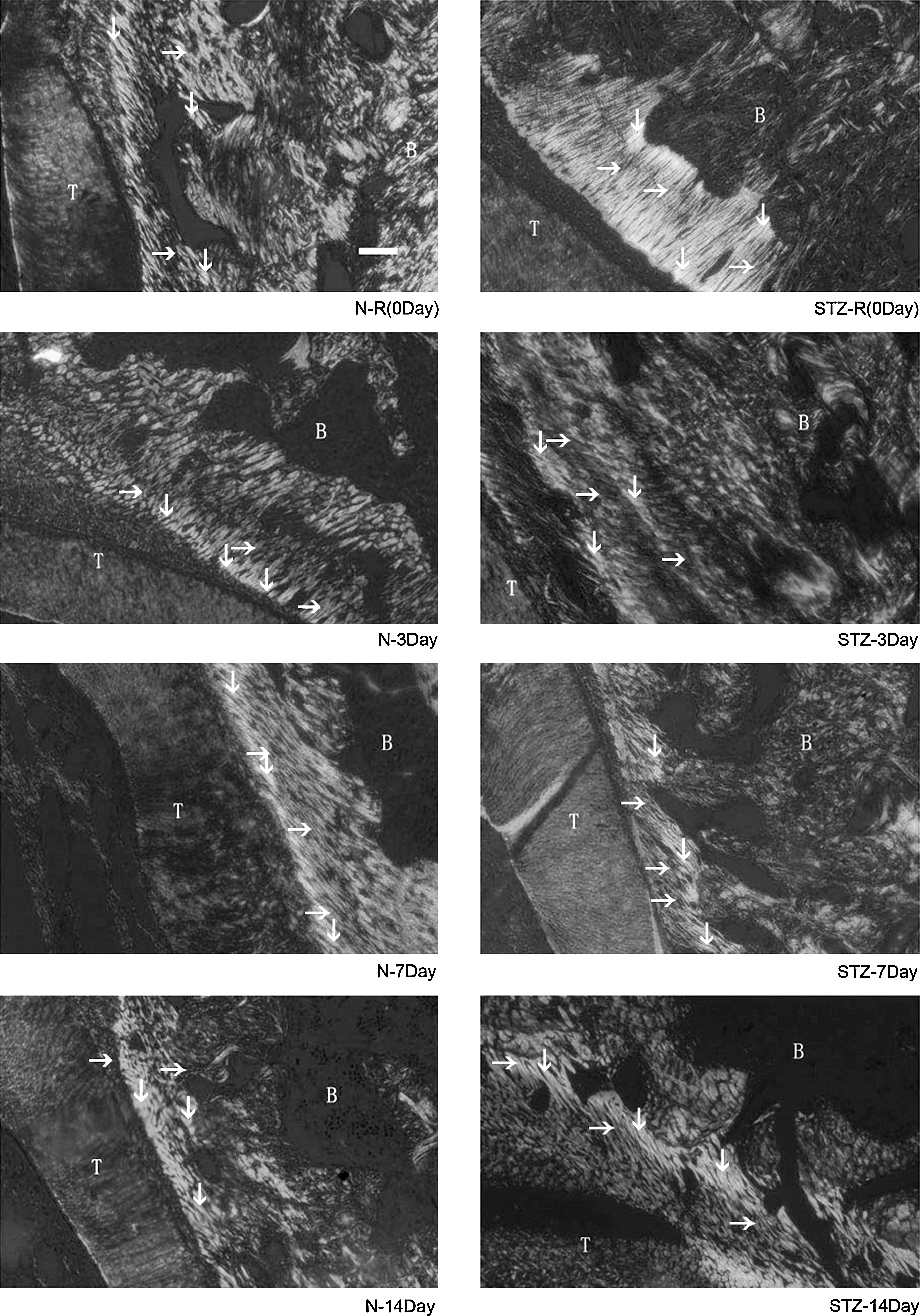
Histological appearance of collagen fibers (picrosirius red staining) in normal and diabetes groups under control conditions and in Area D (tension side) after orthodontic induction in the PDL. ↓, Col-I; →, Col-III; T, teeth; B, alveolar bone.
Expression of (
The ratio of Col-III/Col-I was greater in rats with diabetes than in normoglycemic rats with control treatment and for the duration of orthodontic induction (Fig. 6C). At Day 14, the ratio of Col-III/Col-I returned to pre-induction levels in the normal group but remained elevated in the diabetes group (P < 0.05) at nearly two times pre-induction levels.
Discussion
This study explored osteoclast activity, expression of specific collagen types, and expression and inhibition of MMP-1 in the PDL of normoglycemic rats and animals with STZ-induced diabetes before and after orthodontic induction. Results showed that experimental diabetes is associated with a persistent increase in the expression of MMP-1 and Col-III, whereas the expression of Col-I in the PDL is significantly lower than in normoglycemic animals. After orthodontic induction, the diabetes state prolonged orthoclastic activity and enhanced MMP-1 expression as reflected in persistently higher Col-III/Col-I and MMP-1/TIMP-1 ratios in rats with diabetes. The ratio of MMP-1/TIMP-1 in rats with STZ-induced diabetes reached a peak on Day 7, whereas that in normal rats remained nearly constant.
Orthodontic tooth movement within the jaw is based on the ability of surrounding bone and PDL to react to a mechanical stimulus with remodeling processes. 31 Osteoclasts, derived from preosteoclasts recruited from the bone marrow, increase in the PDL during orthodontic stress. 17,31 PDL and gingival remodeling induced by orthodontic force has been partly attributed to modified gene expression of MMPs and their inhibitors. 10,16,23 Diabetes, which increases the risk of periodontal disease, 3 can increase the expression of MMP-1 in periodontal tissues, 36 thus creating an environment conducive to destruction of collagen. 28 This metabolic disorder is also associated with alterations in immune response, inflammation, and extracellular matrix synthesis. 37 –39 In addition, hyperglycemia, a characteristic feature of diabetes, inhibits the proliferation and differentiation of PDL fibroblasts, which can contribute to the delayed periodontal regeneration and healing seen in patients with diabetes mellitus. 29,40 Collagen molecules are also more easily glycated under the hyperglycemic conditions prevailing in diabetes, which can lead to excessive cross-linking of these important structural proteins.
Results from this study were expected to demonstrate differences in protein expression between the experimental groups, and especially in the effects of STZ-induced diabetes on osteoclastic activity and protein expression involved in PDL remodeling. Indeed, our findings showed that osteoclastic activity is initially blunted in diabetes, but significantly more osteoclasts are present in the PDL (at the border of the alveolar bone) of rats with STZ-induced diabetes compared with normoglycemic rats 14 days after orthodontic induction. This finding is consistent with earlier studies in similar rat models of experimental diabetes showing slow osseous regeneration after orthodontic movement. 30
Rapid bone resorption and marked PDL destruction are features of orthodontic movement in animal models of diabetes (and in the clinical setting). The pathobiology demonstrated in the current experimental model provides a basis for better understanding of these phenomena. Bone absorption occurs at the induction of appropriate orthodontic force but is soon replaced by bone remodeling in subjects without diabetes, resulting in orthodontic movement. Our study found a transient increase in osteoclasts in the PDL at the edge of the alveolar bone, which returned to a normal level gradually in the normal group. However, in study animals with diabetes, the elevated numbers of osteoclasts were sustained for a longer time under the same force, leading to extended period of the bone destruction.
Earlier studies have also demonstrated weakening of the PDL in experimental diabetes. 30 The PDL derives its strength from Col-I fibers, whereas Col-III creates more delicate fibrils responsible for tissue elasticity. 41 The presence of Col-III helps mitigate tension placed on the PDL during orthodontic treatment. This effect is also seen in remodeling of the periodontium due to occlusal force. 42 It is believed that Col-III accumulation is increased relative to Col-I in the earlier phases of collagen remodeling, 41 especially under tension force. During PDL remodeling, Col-III, which is prevalent during proliferation, is gradually replaced by Col-I until a normal ratio of Col-III/Col-I is achieved. 41 In our study, there was a progressive accumulation of Col-III on the tension side in rats with diabetes, culminating in a significantly higher level in the PDL at Day 14 compared with the normoglycemic group. In contrast, the accumulation of Col-I was comparatively blunted and delayed (on both tension and pressure sides) in rats with STZ-induced diabetes. These results suggest that the PDL is more “vulnerable” in diabetes, and the PDL requires more time to remodel because a higher ratio of Col-III/Col-I persists. At a later stage (i.e., 14 days after orthodontic induction), the differences in osteoclatic activity and Col-III/Col-I ratio are significant, indicating that rats with STZ-induced diabetes maintained high levels of destruction of alveolar bone and degradation of the PDL, whereas these indices in normoglycemic rats declined to near control levels.
In addition to collagens, MMP-1 and TIMP-1 play an important role in remodeling of the PDL during orthodontic tooth movement. A strong correlation between mechanical force and induction of the MMP-1 gene has been reported, 23 with MMP-1 causing the degradation of collagen fibers and other extracellular matrix components. 43 This proteolytic enzyme, which can induce a rapid and almost complete breakdown of extracellular matrix collagen, 44 is countered by TIMP-1. 45 A balance of MMPs and TIMPs is central to the stabilization of the extracellular matrix. 24 In this study, elevated expression of MMP-1 was found in the pressure side of the PDL of rats with STZ-induced diabetes. Furthermore, the ratio of MMP-1/TIMP-1 in rats with STZ-induced diabetes reached a peak on Day 7, whereas the ratio in normal rats remained at near control levels.
This elevated ratio of MMP-1/TIMP-1 in experimental diabetes, which reflects increased MMP-1 and decreased TIMP-1 expression, can disrupt the delicate balance of collagen degradation and remodeling in the PDL and cause prolonged degradation of the PDL compared with normal rats. In normoglycemic rats, the ratio of MMP-1/TIMP-1 changed little mainly because of simultaneous increases in both MMP-1 and TIMP-1. Thus, the degradation and remodeling of collagens remained in dynamic equilibrium even under orthodontic pressure. In contrast, the disturbed collagen metabolism in diabetes manifested in the altered expression of MMP-1 and TIMP-1 and MMP-1/TIMP-1 ratio.
Overall, in normoglycemic rats, levels of MMP-1, TIMP-1, and Col-III and the ratio of Col-III/Col-I peaked on Day 7 and declined thereafter, reflecting initial remodeling and subsequent restoration of the physiologic balance and stabilization of the extracellular matrix. Conversely, disturbed collagen metabolism persisted in rats with STZ-induced diabetes, resulting in extended remodeling and reconstruction of the PDL, which are disadvantages in the setting of orthodontic force.
Orthodontic tooth movement results from the resorption and formation of alveolar bone and the degradation and remodeling of the PDL. Osteoclastogenesis and dynamic changes to the collagenous composition of the PDL are key processes. Our findings indicate these events are disturbed and prolonged in diabetes so that rats with STZ-induced diabetes have compromised recovery from damage caused by orthodontic stress. Extrapolating these findings to the clinical setting suggests that the course of orthodontic treatment would be longer in patients with diabetes than in normoglycemic individuals. This pilot study is the only investigation to date to observe the longitudinal influence of hyperglycemia on the alveolar bone and PDL during orthodontic force. Further studies should be carried out to explore the time required for the alveolar bone and PDL recover to normal levels under orthodontic force in experimental diabetes and the potential impact of pharmacotherapy aimed at controlling blood glucose level on improving the degradation and remodeling of the PDL and resorption and formation of alveolar bone.
Footnotes
Acknowledgments
The authors acknowledge the help of Prof. R.B. Zhao, Department of Pathology of Harbin Medical University, Harbin, China, for performing the immunohistochemistry and her continuous support and encouragement. The authors also thank Prof. G.Z. Fang, Harbin University of Science and Technology, for performing the image processing. This work was supported, in part, by grant 2007RFXQS032 from the Harbin Science and Technology Bureau.
Author Disclosure Statement
No competing financial interests exist.