
Abstract
Background:
Tight glycemic control has the potential to reduce long- and short-term effects of diabetes mellitus. New and improved glucose sensors for short-term implantation in the subcutis offer an alternative to the classical self-monitored blood glucose concentration in the management of diabetes. The use of glucose sensors has an advantage over the latter due to its capacity to obtain continuous glucose measurements. However, instability of in vivo glucose sensor measurements has been reported, and this bioinstability is likely to be influenced by the inflammatory reaction to the implanted sensor. Gene expression analysis is now performed in an existing porcine model of subcutaneous glucose sensor implantation to investigate the time course of inflammation from a new perspective.
Methods:
Tissue surrounding glucose sensors was sampled to different time points (2 h, 24 h, 3 days, and 7 days) after implantation in the subcutis of pigs. From the tissue RNA was extracted, cDNA was synthesized, and real-time reverse transcription-polymerase chain reaction (RT-PCR) was performed for the quantification of immunoregulatory genes.
Results:
Genes coding for adhesion molecules, chemokines, cytokines, CD markers, and antigen presentation molecules were differentially expressed over time. Most of the investigated genes were significantly up-regulated 24 h and 7 days after implantation.
Conclusions:
The present study demonstrated that the technology for targeted multiple-gene expression by real-time RT-PCR is useful in the evaluation of the immune response to subcutaneously implanted glucose sensors and that the expression levels also seemed to correspond to the histomorphological observations over time.
Introduction
Self-monitored blood glucose concentration has become an integrated part in the management of diabetes, 3 and intensive self-monitoring of blood glucose is effective for the control and treatment of diabetes. 4,5 However, this method is noncompliant and only provides a snapshot of the glucose concentration. By contrast, continuous glucose monitoring delivers the information needed for optimal control of glucose levels. 6
Since the relation between blood and interstitial fluid glucose concentration was demonstrated, 7 numerous invasive techniques have been developed for continuous glucose monitoring. 8 –11 However, previous work demonstrates that continuous glucose measurements are inaccurate and unstable when sensors are implanted in subcutis. 6,12 –16 Suggested causes of the observed bioinstability of sensor measurements are protein/cellular biofouling at the membrane surface, tissue interferents affecting the electrode, enzymatic dysfunction, and unstable levels of oxygen. 12,17 –20 It seems that some of these problems are associated with the inflammatory reaction at the implantation site. However, only a few reports on the inflammatory reaction and biocompatibility of subcutaneously implanted glucose sensors for short-term implantation are available. 6,12,21 –30 These studies were only based on a histomorphological evaluation of the inflammatory reaction. Therefore, a thorough examination of the inflammation caused by subcutaneously implanted glucose sensors requires the use of additional assays and methods.
In order to further understand what happens at the tissue–sensor interface we characterized the histomorphological changes and examined the effect of sensor implantation on the mRNA expression over time in the subcutis of pigs. Specifically, we quantified different immunomodulatory genes important for the development of inflammation by real-time reverse transcription-polymerase chain reaction (RT-PCR).
Materials and Methods
Animals
Four female crossbred Danish Landrace-Yorkshire-Duroc pigs with a weight of approximately 75 kg were used. The pigs were acclimatized for 2 weeks in multiple pens before the experiment started. During the experimental period they were housed in single pens, at a room temperature of 18–22°C and a relative humidity of 30–70%. The pigs were fed twice daily with a commercial swine fodder (27501-A, Prima Antonio, Oestsjællands Andel, Slangerup, Denmark) and had free access to tap water. The experiment was carried out in accordance with animal welfare guidelines provided by the Animal Experiments Inspectorate, Ministry of Justice, Copenhagen, Denmark.
Glucose sensors and implantation
Glucose sensors were fabricated for implantation after modifications of Moussy et al. 31 and as described previously. 30 In brief, the electrochemical sensor consisted of Kapton® (DuPont, Wilmington, DE) foil with screen-printed electrodes (Pt and an Ag/AgCl), one electrode on each side. The working electrode was covered with three membranes/polymeric layers: an inner membrane made of cellulose triacetate Selectophore (Fluka, Buchs, Switzerland) and Nafion® (5% perfluorinated, Sigma-Aldrich, St. Louis, MO), an enzyme layer of glucose oxidase (G7016, Sigma-Aldrich) cross-linked by glutaraldehyde (G5882, Sigma-Aldrich), and an outer membrane that consist of a mixture of silicone (Dow Corning, Midland, MI) and swellable polyurethane (Noveon, Cleveland, OH). The current flow in the amperometric glucose sensor is generated from the enzyme-catalyzed production of hydrogen peroxide as described recently. 32 –34 The sensors were sterilized at 25 kGy by electron irradiation, sterile-packaged, and stored at 4°C until use. Moreover, extracts of sensors were tested for the presence of endotoxins according to U.S. Pharmacopeia 26 Method 85 (Bacterial Endotoxins Test, Gel Clot Limit Test) with the Pyrogent Plus Gel-Clot Limulus amoebocyte lysate test kit (Bio Whittaker, Cambrex Bio Science, Copenhagen) having a lysate sensitivity of 0.125 EU/mL as described previously. 30
Four sensors (batch Hj001-006) were implanted for 2 h, 24 h, 72 h, and 7 days in the subcutis of each of the four pigs (n = 16 per time point). For every time point a sham procedure was performed, i.e., the implantation procedure was carried out without leaving a sensor in the subcutis (n = 4 per time point). The implantation of sensors in the subcutis was performed as described previously. 6,29,30 For histological and immunohistochemical evaluation tissue from all pigs and time points (n = 16) were assessed (sham n = 4). For real-time RT-PCR skin around two sensors and one sham procedure per time point in each of three pigs were assessed (n = 6) (sham n = 3).
Tissue sampling
The pigs were anesthetized with a mixture (1 mL/15 kg) of 25 mg/mL zolazepam/tiletamin (Zoletil ® 50 veterinary grade, Virbac International, Carros, France), 0.65 mg/mL medetomidine (Domitor ® veterinary grade, Orion, Espoo, Finland), 12.5 mg/mL ketamine (Ketaminol® veterinary grade, Intervet International B.V., Boxmeer, The Netherlands), and 2.5 mg/mL methadone (Metadon®, “DAK,” Nycomed, Roskilde, Denmark). With the animal under anesthesia a tissue cube (4 × 4 × 2 mm) was sampled around the implanted sensor tip and from sham-operated tissue. As the sensors were approximately 5 cm long and implanted at an angle of 45° to the skin surface, the sensor tip was consistently placed in the subcutaneous tissue and not in the dermis or muscle, i.e., all samples evaluated were from the subcutaneous compartment. Samples were collected in 1.0-mL cryotubes, immediately snap-frozen in liquid nitrogen, and stored at −80°C. Simultaneously, subcutaneous tissue was sampled for histology, i.e., samples were put in cryomolds, covered with OCT™ compound (Tissue Tek® OCT compound, Sakura FineTek, Værløse, Denmark), and snap-frozen in isopentane cooled on dry ice. Finally, the pigs were euthanized with an intravenous overdose of 30 mL of pentobarbital (200 mg/mL) (Pharmacy, Faculty of Life Sciences, University of Copenhagen).
For histopathological examination tissue adjacent to tissue sampled for RNA extraction was cut into 0.2- × 2- × 2-cm slabs containing the sensor, fixed for 30–36 h in 4% paraformaldehyde, and processed by standard histological procedures through graded concentrations of ethanol, Estisol (Estisol 220, Esti Chem, Koege, Denmark), and embedded in paraffin. Tissue sections were cut at 3–4 μm and stained with hematoxylin and eosin for routine histopathological evaluation. Microscopic evaluation of all tissue sections was performed with an Olympus (Ballerup, Denmark) AX70 light microscope.
RNA isolation, cDNA synthesis, and real-time RT-PCR
The tissue sample was ground to a fine powder in the presence of liquid nitrogen with a pestle and mortar. The powder was immediately transferred to a 15-mL tube and homogenized with a Polytron PT 3100 (Kinematica AG, Lucerne, Switzerland) in 4 mL of Trizol® reagent (Invitrogen, Life Technologies, Carlsbad, CA) for lysis of cells.
Extraction of RNA was performed by adding 200 μL of chloroform to Eppendorf tubes (Eppendorf, Hamburg, Germany) per 1,000 μL of Trizol reagent. Tubes were shaken vigorously by hand for 15 seconds and incubated for 2 min. Eppendorf tubes were then centrifuged with a tabletop centrifuge at 12,000 g for 20 min at 4°C, and the aqueous phase was removed and mixed gently with 500 μL of isopropanol in an Eppendorf tube to precipitate the RNA. After incubation for 10 min the Eppendorf tubes were centrifuged for 10 min (12,000 g in the centrifuge) at 4°C, and the liquid was poured off. The pellet was then washed in 75% ethanol, followed by centrifugation for 10 min (9,000 g in the centrifuge) at 4°C, and then the ethanol was poured off and pipetted out if necessary. Finally, the pellet was dissolved by adding 5–15 μL of RNase-free water (catalog number D5758, Sigma Chemical Co., St. Louis) depending on the size of the pellet. The RNA concentration and integrity were measured using an Agilent 2100 Bioanalyzer according to the manufacturer's instructions (RNA 6000 Nano Assay, Agilent Technologies, Palo Alto, CA).
First-strand cDNA synthesis was made from a total of 5 μg of RNA using SuperScript™ II reverse transcriptase and oligo(dT) primers according to the manufacturer's instructions (Invitrogen, Life Technologies) and as previously described.
35
Real-time RT-PCR was performed using the cDNA as a template for target-specific amplification to evaluate a panel of genes related to inflammation, i.e., cytokines, chemokines, and cell surface molecules as shown in Table 1. All probes and primers for real-time RT-PCR were designed using the Primer Express (Applied Biosystems, Foster City, CA, USA) software package and nucleotide sequences obtained from GenBank or the TIGR porcine EST database.
36
These sequences are available online at
Data were obtained from searches in the online databases
SLA, swine leukocyte antigen.
Immunohistochemistry
For immunohistochemical detection of CD3ɛ, CD4a, and CD8b, three serial cryostat sections approximately 5 μm thick from all tissue samples were cut onto SuperFrost®Plus glass slides (Menzel-Gläser, Braunschweig, Germany) and air-dried. The cryostat-cut sections were fixed in cold acetone (4°C) for 10 min and air-dried. In all the immunohistochemical protocols the alkaline phosphatase-conjugated Powervision + poly/AP anti mouse/rabbit kit (ImmunoVision Technologies, Daly City, VA) was used as the detection system according to the manufacturer's instructions. All washings and dilutions were done in Tris-buffered saline (0.05 M Tris and 0.15 M NaCl, pH 7.6) except for the primary antibodies, which were diluted in 0.1% bovine serum albumin (catalog number A7906, Sigma, Brøndby, Denmark) in Tris-buffered saline. The primary antibodies used were all mouse monoclonal antibodies specific for porcine CD3ɛ (clone PPT-3 [catalog number 4510-01], SouthernBiotech, Birmingham, AL), porcine CD4a (clone MIL17 [catalog number MCA1749], AbD Serotec, Kidlington, UK), or porcine CD8b (295/33-25, Labor Dr. Glatthaar, Reutlingen, Germany). All incubations were performed at room temperature apart from the incubation of the primary antibodies, which was overnight at 4°C. Development was done with Fast Red (catalog number 4210, Kem-En-Tec, Taastrup, Denmark), and Mayer's hematoxylin was used as a counterstain. Cryostat-cut sections of a porcine lymph node were included as a positive control, and negative controls included incubation without primary antibody and substitution of the primary antibody with nonsense antibody of matching isotype.
All immunostainings were scored semiquantitatively: no (0), few (1), moderate (2), many (3), or massive (4) number of stained cells in the cellular infiltrate surrounding the implanted sensor.
Statistical analysis
Statistical difference in subcutaneous mRNA expression (Ct value) between sensor implantation and sham operation was evaluated by analysis of variance with Fisher's least significant as a post hoc test for individual comparisons to the time points 2 h, 24 h, 72 h, and 7 days post-implantation (PI). P < 0.05 was considered statistical significant, and power calculations were performed with a minimal acceptable cutoff of 0.8.
Results
The Limulus amoebocyte lysate test showed an endotoxin concentration of less than 1.25 EU per sensor, which is below levels for clinical testing of implantable medical devices. The histomorphological characterization of skin around sensors implanted from 2 h to 7 days corresponded to previous published data with a similar in vivo setup. 29,30 In brief, after 2 h few infiltrating neutrophils were observed, and fibrin lined the sensor cavity. After 24 h this fibrin layer was occasionally intermingled with basophilic layers of nuclear material possibly originating from necrotic neutrophils. The inflammatory cells in the vicinity of the sensor surface were primarily composed of neutrophils and macrophages, and fibrin exudation/hemorrhage was a consistent finding. Immunohistochemistry revealed few CD3-, CD4-, or CD8-positive cells (Fig. 1). After 3 days, the extent of the hemorrhage/fibrin exudation and infiltration of inflammatory cells increased, i.e., a mixed infiltration of neutrophils and macrophages. Hemorrhage together with edema was often observed at the margin of the sensor cavity. Occasionally, formation of granulation tissue, a few T lymphocytes (Fig. 1), and/or a foreign body reaction with giant cells and epitheloid cells was seen. At day 7, the infiltration of inflammatory cells (macrophages, lymphocytes [Fig. 1]), granulation tissue formation, and foreign body reaction was more pronounced. The sham procedure did in general not result in an inflammatory reaction. However, mild hemorrhage could be observed at the insertion site after the sham procedure was performed.
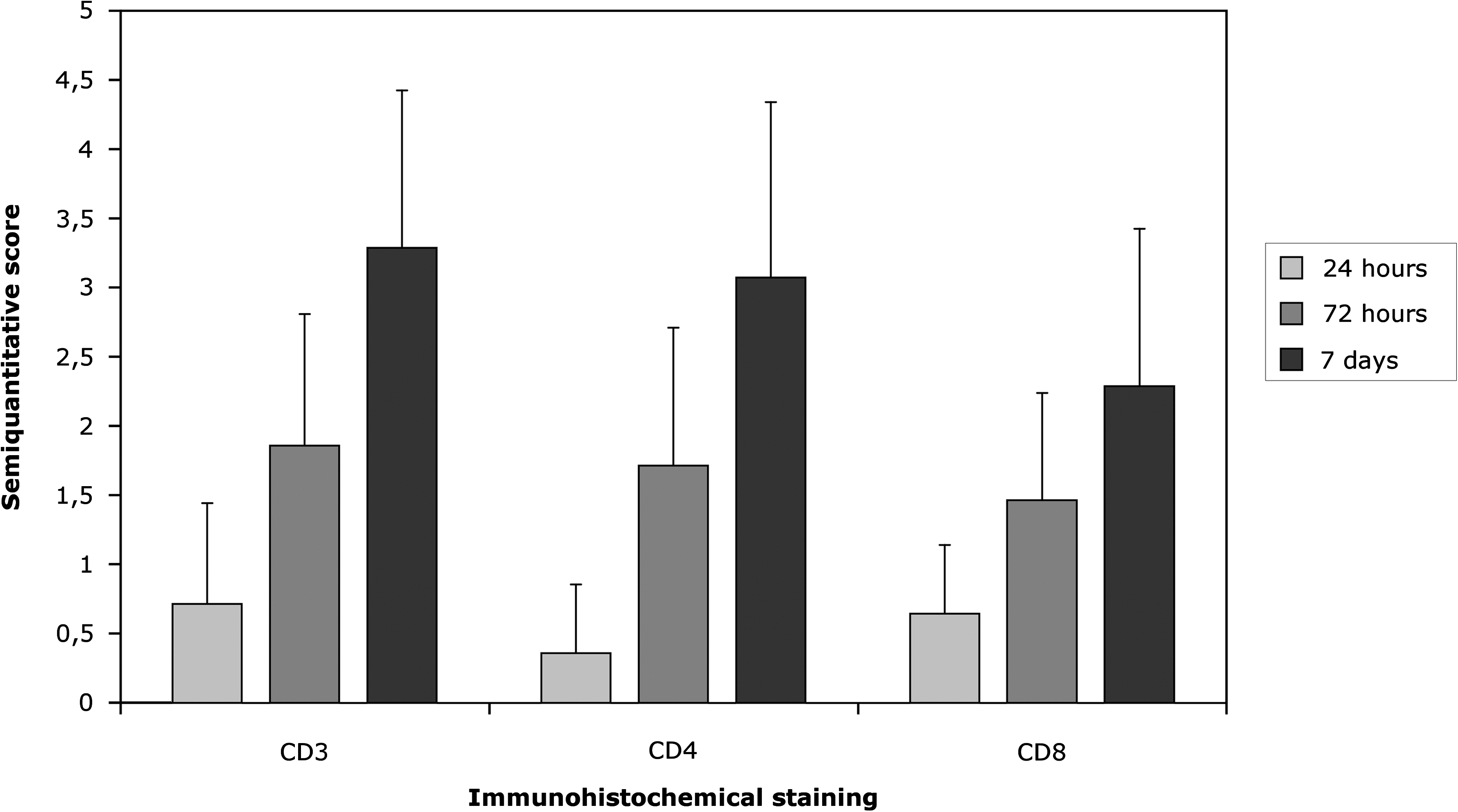
Semiquantitative score (mean + SD) of the extent of protein expression of CD3ɛ, CD4a, and CD8b as assessed by immunohistochemistry in the tissue around implanted sensors from 2 h to 7 days after implantation (n = 16 per time point). No positive staining was observed 2 h post-implantation and in sham controls (n = 4).
The 27 evaluated immunomodulatory genes and one housekeeping gene are described in Table 1, and their expression over time after sensor implantation in subcutis is presented in Figure 2. In brief, the expression of mRNA was significantly up-regulated at the following time points after implantation: 2 h PI for interleukin (IL) 6; 24 h PI for intercellular adhesion molecule 1 (ICAM1), integrin β2 antigen (ITGB2), monocyte chemoattractant protein 1 (MCP-1) (CCL2), IL8, IL1B, IL6, tumor necrosis factor (TNF), colony-stimulating factor (CSF) 2, triggering receptor expressed on myeloid cells (TREM) 1, TREM2, CD4, CD86, immunoglobulin heavy constant μ (IGHM), swine leukocyte antigen (SLA)-DM α chain (DMA), SLA-DQ α chain (DQA1), SLA-DR α chain (DRA), IL2, and IL5; 72 h PI for ICAM1, CCL2, IL8, IL1B, IL6, TREM1, TREM2, and IL5; and 7 days PI for ICAM1, ITGB2, CCL2, IL8, IL1B, IL6, TNF, lymphotoxin α (LTA), CSF2, TREM1, TREM2, CD3E, CD4, CD8A, CD80, CD86, IGHM, SLA-DMA, SLA-DQA1, SLA-DRA, IL2, IL4, IL5, and interferon-γ (IFNG). Nitric oxide synthase (NOS) 2 mRNA expression remained unchanged at all time points. Moreover, the regulatory genes IL10 and transforming growth factor β1 (TGFB1) were significantly up-regulated at 24 h, 72 h, and 7 days PI. In summary, the expression of inflammatory-associated genes was up-regulated in the subcutis with implanted glucose sensors compared to sham operation.
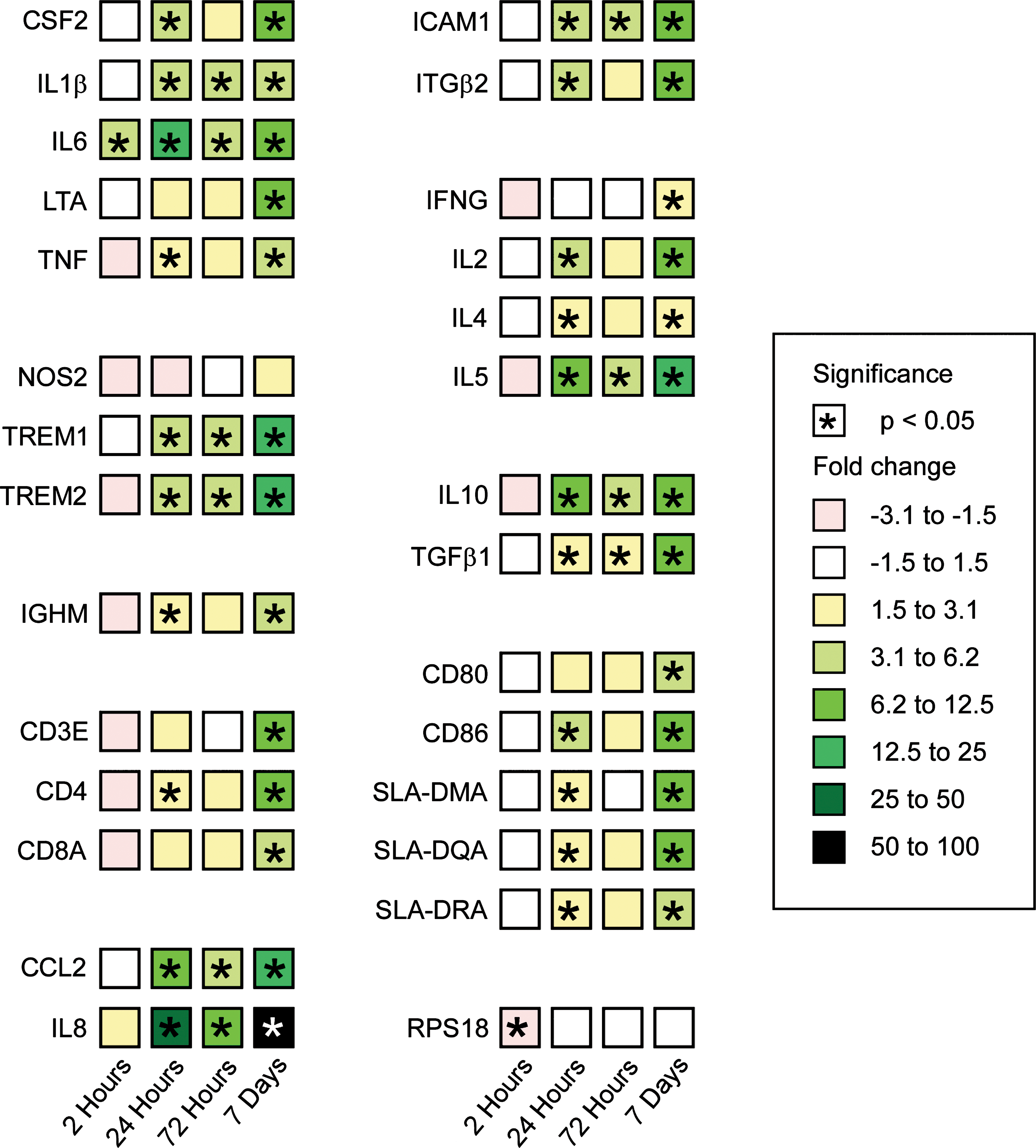
Real-time reverse transcription-polymerase chain reaction analysis of gene expression in subcutis 2 h (n = 6), 24 h (n = 6), 72 h (n = 6), and 7 (n = 6) days post-implantation of glucose sensors in the subcutis. The data represent the mean fold change in gene expression compared to sham operation (n = 3). Squares are color-coded to denote either up- or down-regulation of gene expression. See Table 1 for gene abbreviations. *Significant difference at the level of 0.05.
Statistically, the effects of the major players described were large, i.e., the great majority the analysis of variance was P < 0.0001, and the study was adequately powered as all but two of the power calculations exceeded 0.8, and the great majority are 1.000. The two that failed were housekeeping genes that did not appreciably change in expression (data not shown).
Discussion
From this and other studies toxic substances, 30 endotoxins in the glucose sensor, and massive needle trauma 30 induced at insertion of the glucose sensor can be ruled out as major influencing factors of the observed tissue reaction caused by sensor implantation. Therefore, it is likely that in this study the observed inflammatory reaction, i.e., the histological reaction and gene expression profiles, is primarily related to the physical and chemical properties of the sensor.
Specifically, ICAM-1 is a molecule on endothelial cells and binds the integrin leukocyte functional antigen 1 (LFA-1 or CD18). LFA-1 is expressed by different immune cells and is important in the transendothelial migration. The role of the chemokines IL8 and MCP-1 is twofold. First, they trigger changes in the conformation of integrins, stabilizing adhesion. 38,39 Second, both proteins direct the migration of leukocytes through a concentration gradient. IL8 and MCP-1 mainly attract and activate neutrophils and monocytes, respectively. IL8 production may be induced by a variety of stimuli, including pro-inflammatory cytokines (e.g., IL1, TNF-α). 40 Three genes in our study encode for pro-inflammatory cytokines, i.e., TNF-α, IL1β, and IL6. These cytokines mediate a variety of actions, among which induction of acute inflammation and their central role for the innate immunity are important. IL6 is known for its role in the acute-phase response in experimental lung infections in pigs, 41 and IL1α and TNF-α have previously been detected in situ by immunohistochemistry 1 h and 24 h, respectively, after implantation of sensors in the subcutis of pigs. 29 Moreover, LTA may play a role in the inflammatory reaction to sensors 7 days PI. LTA mediates a large variety of inflammatory, immunostimulatory, and antiviral responses. Granulocyte-monocyte CSF primarily stimulate growth and differentiation myelomonocytic cell lineages, and its production can be induced by TNF-α and IL1. 42 TREM1 and TREM2 are newly discovered receptors: TREM1 activates and stimulates degranulation of IL8 and myeloperoxidase from neutrophils and secretion of MCP-1, IL8, and TNF-α from monocytes, 43,44 whereas TREM2 is expressed on macrophages and dendritic cells but not on neutrophils and monocytes, suggesting a role in the chronic inflammatory reaction. 44 Thus, distinct TREM receptors may regulate acute and chronic inflammation through the action of myeloid cells. The NOS2 gene encoding inducible NOS remained unchanged during our experiment. Its role in bacterial infections has previously been elucidated in pigs, 45 and therefore this finding indicates that bacterial contamination is minimal in our model as previously shown. 30
Genes encoding receptors on T lymphocytes (CD3ɛ, CD4, and CD8α), receptors on B cells (immunoglobulin M [IgM]), receptors on antigen-presenting cells (APCs) (CD80, CD86, SLA-DMA, SLA-DQA1, and SLA-DRA), and cytokines (IL2, IL4, IL5, and IFNγ) were differentially up-regulated (Table 1). CD3ɛ constitutes a part of the CD3 complex expressed on T cells, is associated with the T-cell antigen receptor, and required for signal transduction through T-cell antigen receptors. 46 CD4 are expressed on T-helper cells 1 (TH1) and 2 (TH2), but also on monocytes and macrophages, and is a co-receptor for major histocompatibility complex class II molecules (SLA in pigs). CD8α constitutes a part of the CD8 molecule expressed on cytotoxic T cells. CD8 is a co-receptor for major histocompatibility complex class I molecules. 39 Thus, one could speculate that 7 days PI the immune response to sensors are partially directed toward a more specific cellular component as all three molecules were significantly up-regulated at that time. However, as porcine natural killer cells and macrophages express CD4 and CD8, respectively, the innate immune system might also be the primary contributor to the expression of these CD molecules. Therefore, more determinative staining would have to be performed to elucidate if the subcutaneous reaction to the sensors develops into an adaptive immune response at day 7. To translate the gene expression levels to protein levels in the tissue an immunohistochemical staining for CD3ɛ, CD4a, and CD8b was performed as these three genes were significantly up-regulated 7 days PI. The in situ protein level seems to correspond well to the gene expression levels, i.e., the protein expression of these CD markers increased over time (Fig. 1). Moreover, the finding of up-regulated CD3ɛ mRNA at 7 days PI is also supported by a previous report on the detection of CD3ɛ at the protein level by immunohistochemical detection. 29 It has been shown that TH cell effector function and production of IL4 is dependent on T-cell interaction with the co-stimulatory molecules CD80 and CD86 expressed on APCs in a model of murine infection. 47 This further supports the hypothesis of a more specific cellular reaction to implanted sensors as the genes encoding for the co-stimulatory molecules CD80 and CD86 were significantly up-regulated at 7 days PI. This is further supported by up-regulation of the cytokine IL4, a key regulator of activated B (isotype switch) and T cells (differentiation of CD4 + T cells into TH2 cells) cells. The swine histocompatibility complex II molecules SLA-DM, DQ, and DR α chains expressed on APCs are thought, like human leukocyte antigens, to play an important role in class II restricted antigen processing and presentation in swine and is known to be up-regulated by IL4. 48 IL2, IL4, IL5, and IFNγ are mainly produced by T cells and are involved in T-cell proliferation (autocrine), B-cell activation, eosinophil growth differentiation, and macrophage activation (e.g., increased expression of SLA), respectively. 39 IgM is expressed on naive B cells as their B-cell receptor. Upon stimulation, IgM is the first antibody produced in response to antigen and is normally of low affinity. IgM-positive cells have previously been identified in very few numbers at 7 days PI. 29
IL10 along with TGF-β is also produced by T-cell subpopulations termed regulatory T cells. Both cytokines are involved in the regulation of the cytokine network. IL10 is a potent suppressor of APCs, especially macrophage functions. Thus, IL10 inhibits monocyte and macrophage synthesis of different cytokines like IL1, IL6, IL8, TNF-α, and granulocyte-macrophage CSF in humans. 49,50 Furthermore, TGF-β is a multifunctional peptide that controls proliferation, differentiation, and many other functions in different cell types. Wang et al. 51 showed that healing of a skin wound in pigs coincided with a significant up-regulation of TGF-β mRNA.
The study design makes it possible to elaborate on the functional consequences our findings might have for the tissue response continuum and compare it to what is generally known from biological responses to materials. 52,53 Clearly, at 2 h PI the coagulative system is activated, and fibrin is observed at the tissue–sensor interface together with a few neutrophils. IL6 is potentially up-regulated as a consequence of endothelial cell activation as no inflammatory cells are observed histologically. During the next 22 h neutrophils and macrophages infiltrate the skin through specific interactions between complementary adhesion molecules expressed on these leukocytes and the endothelial cells. 53 The up-regulation and modulation of adhesion molecules (ICAM-1, LFA-1 [ITGB2]) on the surface of these cells is modulated by pro-inflammatory cytokines (IL1, TNF-α, IL6) and chemokines (IL8, MCP-1), respectively. 38 The observation of macrophages at this stage of inflammation is further supported by the APC-specific up-regulation of major histocompatibility complex class II molecules and the co-stimulatory molecule CD80. Although present in few numbers, the T cell-associated genes (CD3ɛ, CD4, CD8α, IL2) are up-regulated at this point together with the T cell- or mast cell-derived cytokines IL4 and IL5. Additionally, the coagulation system is still activated, resulting in hemorrhage and fibrin exudation. Essentially the same picture is seen at 72 h PI, but now the infiltration of T cells has increased. Moreover, the initial generation of granulation tissue is visible—a process that, among other growth factors, is under the influence of TGF-β, IL1, or TNF-α. 53 It could be speculated that down-regulation of the major histocompatibility complex class II molecules and CD80 at 72 h PI is caused by IL10's ability to inhibit macrophage function. 39 After 7 days T cells and macrophages dominate the cellular infiltrate, which is confirmed at the histological and the gene expression levels as essentially all genes are up-regulated, i.e., now the T cell-derived cytokines IL2 and IFNγ are also up-regulated. Furthermore, foreign body reaction is seen together with up-regulation of IL4, which is directly involved in the generation of giant cells. 54,55 Additionally, fibrin at the tissue–sensor interface and genes important for endothelial transmigration of cells are still present at 7 days PI.
In conclusion, the course of events is comparable to what is generally known from implantation from medical devices. 52,53 However, the presence of fibrin around the sensor seems to play a role throughout the implantation period and is probably a result of increased vascular permeability caused by inflammation but potentially also the sensor causing direct physical damage to vessels during movements. It has been reported that proteins adsorbed to the biomaterial may undergo changes in protein conformation exposing previously hidden epitopes. 56 –58 An example is the D domain of fibrinogen, which binds tightly to biomaterial surfaces resulting in a conformational change of the protein exposing a P1 epitope. The P1 epitope reacts with the macrophage integrin Mac-1 (LFA-1). Subsequently, the interaction of Mac-1 and the P1 epitope triggers phagocyte adherence and activation. 58 Therefore, the observed fibrin/fibrinogen around the sensor seems to be an important initiator and driver in the inflammatory response to subcutaneously implanted glucose sensors.
The present study demonstrated that the technology for targeted multiple gene expression by real-time RT-PCR is useful in the evaluation of the immune response to a subcutaneously implanted glucose sensor. Furthermore, the temporal nature of this study over a time course from 2 h to 7 days after sensor implantation allows for a reliable description of the gene expression. Findings of up-regulated mRNA from this study also support previously published histochemical and immunohistochemical results. 20,29,30 In these it was argued that the functionality of glucose sensors is dependent on modulation of the immune response. 20 However, a targeted modulation of the immune response to implanted sensors also requires detailed knowledge of how it develops. Therefore, we evaluated the inflammatory response to glucose sensors by real-time RT-PCR. The porcine model is useful in terms of size (its own implantation control), pathophysiological and anatomical similarities to humans, 59 –61 and it allows a systematic evaluation of the immune response to sensors.
In order to increase number of samples (n) per time point we implanted intact sensors without any applied potential as this would have restricted the number of total implanted sensors to six per pig. However, previously we have shown that the potential increase in local hydrogen peroxide concentration, normally electrochemically oxidized under working conditions to 2H + and O2, does not influence the extent of inflammation around the sensors. 30 Therefore, the inflammatory reaction we observe around sensors in this study is representative of what happens when sensors are implanted under working conditions in the subcutis.
This is the first report describing the use of real-time RT-PCR in conjunction with histology to describe the biocompatibility of implantable glucose sensors. Henninger et al. 62 previously evaluated the gene expression profile in tissue surrounding glucose sensors implanted in rat skin. However, they focused on other genes and used another technique, i.e., the Affymetrix® (Santa Clara, CA) GeneChip®. In general, real time RT-PCR is considered to be the most sensitive and reliable method for analysis of gene expression. In contrast, it is also considered to be time consuming and tedious when several genes are to be evaluated.
Conclusions
In conclusion, over time the implantation of glucose sensors in the subcutis results in a multifaceted response. The response includes increasing levels of genes encoding adhesion molecules, chemokines, cytokines, CD antigens, and SLA. Clearly, this study provides insight into the immune and inflammatory mechanisms of glucose sensor implantation in the skin of pigs. Specifically, this includes early innate immunological responses including fibrin exudation. Furthermore, it demonstrates that real-time RT-PCR is a useful tool to evaluate the biocompatibility of implantable glucose sensors, and the data correlate well with the histological picture.
Footnotes
Acknowledgments
We thank Hanne H. Møller for skilful assistance during the in vivo experiment and Ethiopia Beshah and Eudora Jones for their competent and skillful assistance during the work in the laboratory.
Author Disclosure Statement
P.H.K. is an employee of Novo Nordisk A/S.