
Abstract
Patients with diabetes mellitus are characterized by an extraordinary vascular vulnerability. Traditionally, glucose-induced damage to the vascular endothelium is believed to be one of the first steps in the development of vascular damage. However, in the healthy vessel the endothelium is protected by a matrix layer of highly glycosylated proteins that form a physical barrier between the endothelium and the blood flowing past. Although its presence has been known for half a century, this so-called glycocalyx earned little attention from researchers in the past because of an underestimation of its size. In the last decade it has become clear that its full thickness actually exceeds that of the vascular endothelium. Accumulating research into the functional relevance of the endothelial glycocalyx suggests an important role for this layer in the development of cardiovascular disease in diabetes mellitus. Here we will present an overview of the biochemistry of the intact glycocalyx, current methods to assess the glycocalyx, and its possible role in the pathophysiology of cardiovascular disease in diabetes.
Historical Perspective

Schematic representation of size selectivity and charge selectivity.
Long before the publication of the Steno article an interest in the role of glycoproteins in vascular physiology had developed when in the early 1960s intravital microscopy revealed a plasma-free zone adjacent to the vascular endothelium. 2 Based on ruthenium red staining, the concept that a thin layer of negatively charged membrane-bound proteoglycans and glycoproteins covers the surface of most cells was developed. 3 Named after the polysaccharide components of the layer, it was called the “glycocalyx” or “sweet husk.” 4 The early findings indicated that the layer was only a few nanometers thick, and subsequently little attention was paid to its existence. However, mathematical models comparing the hematocrit measured in capillaries with that measured in large arteries suggested the existence of a much larger endothelial surface layer that was able to interfere with flow in the microvasculature. 5 A lack of evidence to support these estimations hampered further investigations into the function and composition of the glycocalyx. However, when in 1996 the thickness of the glycocalyx could be measured in vivo for the first time using a dye-exclusion technique, its thickness proved to be 400–500 nm in capillaries. 6 It then became clear that the glycocalyx must be a dynamic gel-like layer, dependent on the presence of plasma (flow) to maintain its full thickness. This finding established that the endothelial glycocalyx occupied a significant proportion of the microcirculation, and research into this component of the vasculature was given a new impulse.
Biochemistry of the Intact Glycocalyx
The endothelial glycocalyx can be pictured as a highly hydrated layer of endothelium- and plasma-derived macromolecules, which are either bound to the endothelial membrane or to each other (Fig. 2). Glycoproteins and proteoglycans are among its main components, serving as anchors to the endothelium. 7 Glycoproteins consist of a core protein and covalently linked saccharide side chains of two to 15 sugar residues. 8 Important examples of the glycoproteins present on the endothelial cell surface are the adhesion molecules, such as selectins, integrins, and immunoglobulins. All have different functions based on their ability to bind different plasma components and the manner in which they are connected to the endothelial cells.
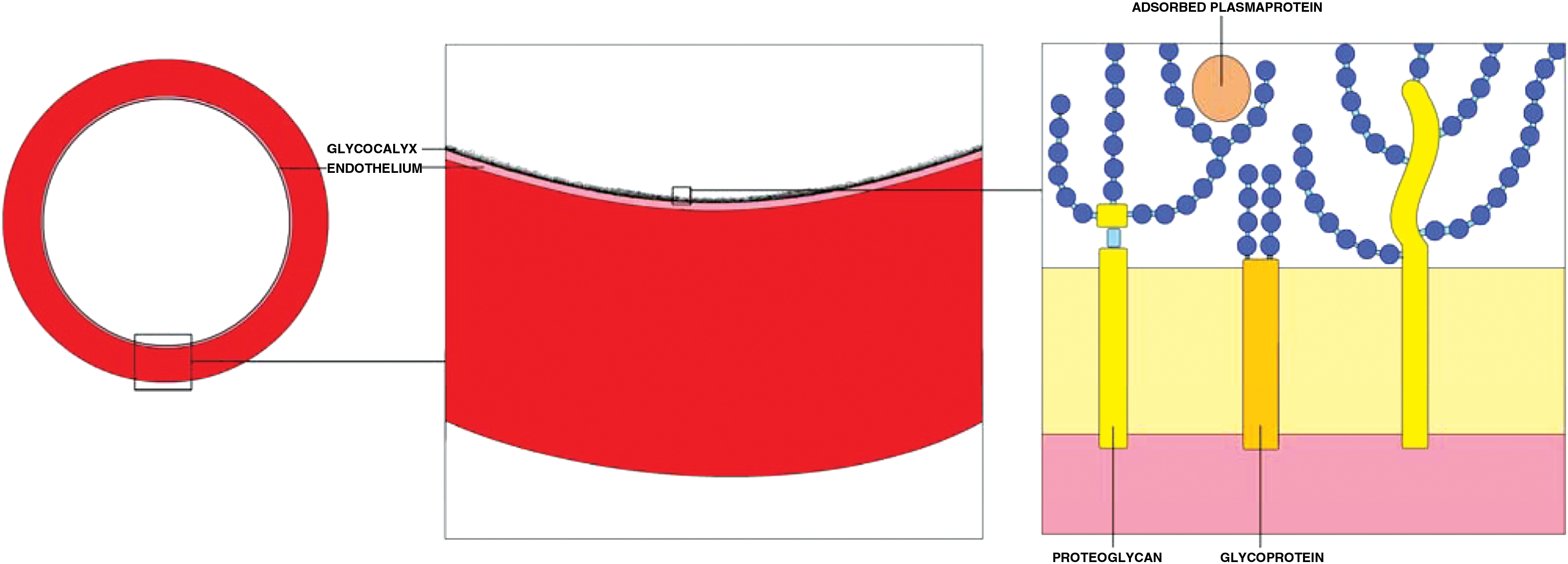
A schematic overview of the endothelial glycocalyx. The glycocalyx is visualized as a brush-like layer on the luminal side of the endothelium. The glycoproteins and proteoglycans serve as anchors to the endothelium. Sugar chains of varying length and consistence are linked to the anchor proteins and form the bulk of the layer. Adsorbed plasma proteins are harbored within the layer.
The proteoglycans are a special group of glycoproteins, which are characterized by very long, unbranched side chains of over 200 sugar residues or glycosaminoglycans (GAGs). 9 GAGs contribute greatly to the negative charge of the glycocalyx because of the occurrence of sulfate uronic acid groups. Three different core proteins with binding sites for GAGs can be found on the endothelial cell: the syndecans, glypicans, and perlecans. 10 The length and geometrical shape of the GAGs determine the extension of glycocalyx into the vascular lumen and its voluminous aspect. The GAGs most commonly found in the vasculature are heparan sulfate (50–90%), chondroitin sulfate, dermatan sulfate, and keratan sulfate. 7 Another GAG abundantly present in the glycocalyx is hyaluronic acid, also named hyaluronan. Hyaluronan is different from other GAGs because it is neither sulfated nor covalently linked to a protein during synthesis. The negative charge of the molecule is due to ionization of the carboxyl groups of the glucuronic acid constituents at physiological pH. Hyaluronan is synthesized by enzymes bound to the cell membrane (hyaluronan synthase 1, 2, and 3) and expelled to either the extracellular matrix or luminal endothelial side of the membrane. 11 There it forms a main constituent of the glycocalyx, contributing greatly to its water binding capabilities. It interacts with other extracellular matrix macromolecules, such as versican and aggrecan, with interactions believed to be essential to the structure and assembly of endothelial glycocalyx. Also, hyaluronan interacts with cell surface receptors, notably CD44, and thereby influences cell behavior. 12 Degradation of hyaluronan is predominantly done by hyaluronidase-1 and −2. 13
Assessment of the Glycocalyx
Research into this layer is traditionally complicated by its collapse when studied ex vivo, most likely due to dehydration. 14 Nevertheless, histological staining of the endothelium has unequivocally demonstrated the presence of the glycocalyx. The early studies made use of dyes such as ruthenium red 15 or Alcian blue, which bind to the negative charges on anionic glycosylated polymers, or used lectin-based stains. 16 More recent research has turned to immunostaining with monoclonal antibodies directed against particular components of the glycocalyx, such as syndecan-1 or heparan sulfates, 17 –19 to more specifically determine compositional changes in the layer.
However, pathological changes in its dimension or permeability may be missed or underestimated when the layer is not studied under conditions of flow. Therefore, Vink and Duling 20 performed intravital microscopy to visualize hamster cremaster muscle capillaries in vivo. They observed a difference between the anatomical diameter of the capillaries and the functional diameter occupied by red blood cells or fluorescently labeled high-molecular-weight dextrans. The authors concluded that between blood and capillary wall a 0.4–0.5-μm-thick glycocalyx was present. They also found a size-dependent half-time for the invasion of the glycocalyx by anionic dextrans, showing that larger molecules took longer to penetrate the glycocalyx. 21
These permeability properties of the glycocalyx were then used to expand the research to human studies. A tracer dilution technology was used to determine the difference between the distribution volumes of a glycocalyx-permeable tracer and a glycocalyx-impermeable tracer, providing a systemic glycocalyx volume. 22 Although this method was laborious and invasive it did result in the first human studies showing the destructive effects of diabetes on systemic glycocalyx volume. 23,24 However, its time-consuming nature and underlying complicated physiology, which may be dependent on more than glycocalyx properties alone, 25 make this method unsuitable for larger-scale studies. The same principle of size-dependent exclusion was used to visualize permeation of the endothelial glycocalyx in vivo in retinal vessels by double-dye retinal angiography (Fig. 3). Two dyes conventionally used in ophthalmological practice to visualize retinal and choroidal vessels, fluorescein and indocyanine green (ICG), proved to have different intravascular distribution properties. Fluorescein is a small molecule and fills up the entire vascular compartment, whereas ICG rapidly binds to large plasma molecules on administration and is unable to fully penetrate the endothelial glycocalyx. ICG most likely does partially penetrate the glycocalyx, as it has been shown that the intravascular distribution of ICG is larger than the intravascular distribution of red blood cells, which are completely excluded from the glycocalyx. 26 Fluorescently labeled albumin has been shown to reveal capillary diameters equal to or bigger than anatomic capillaries, which may indicate incorporation of albumin in the endothelial surface layer. 20,27 Similarly, albumin bound ICG may be incorporated in the glycocalyx but most likely be restricted to the outer regions of the glycocalyx because the layer shows a size-dependent permeability. In line with this, a difference in intravascular distribution of fluorescein and ICG in the retinal vessels was found, suggestive of a zone in the glycocalyx that is impermeable for ICG but permeable to fluorescein.
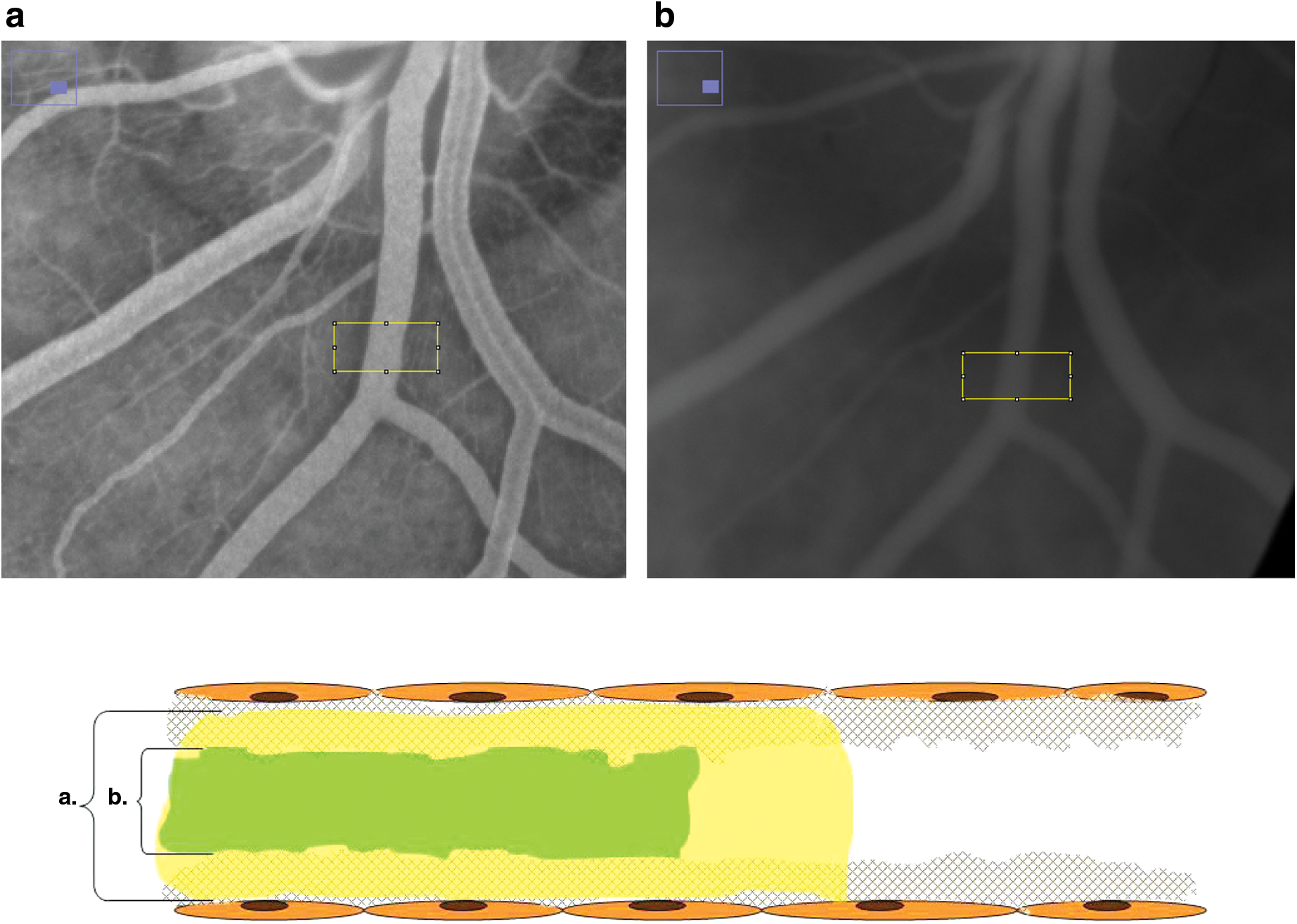
Double-dye retinal angiography: a detailed image and schematic overview of
This difference in intravascular distribution allowed the comparison of the distribution patterns in patients with diabetes and healthy volunteers, revealing a reduced ICG exclusion zone in patients with diabetes. 28
At the same time, a method was developed to determine microvascular glycocalyx dimensions by orthogonal polarization spectroscopy or sidestream darkfield imaging of the sublingual microcirculation. 22 Orthogonal polarization spectroscopy allows the visualization of the smallest sublingual capillaries by visualizing the hemoglobin in the red blood cells. 29 Measurement of the diameter of the red blood cell column in these small vessels provides information on the intravascular space available to erythrocytes. Because leukocytes are much larger than erythrocytes and have been observed to compress the endothelial glycocalyx transiently while passing through the smallest of capillaries, 20 the widening of the erythrocyte column after leukocyte passing became a first measure for in vivo microvascular glycocalyx thickness. 22 Further development of this noninvasive method to evaluate the microcirculation led to an automated assessment of the dynamic range of the erythrocyte column width. This method is based on the principle that erythrocytes are able to transiently occupy the endothelial glycocalyx space, thereby contributing to the dynamic range of the erythrocyte column. Reduced dynamic range of the erythrocyte column width, determined by the distribution pattern of multiple measurements of erythrocyte column width, could therefore be interpreted a reduction of glycocalyx dimension. 30
Several studies have also used plasma measurement of degradation products of the endothelial glycocalyx to establish shedding from the layer. Increased plasma levels of hyaluronan, hyaluronidase, heparan sulfates, and syndecan-1 have all been implicated as markers of damage to the endothelial glycocalyx. 17,19,23,24,31 Further biochemical studies are essential to determine the biochemical composition of the layer and establish shedding patterns.
Effects of Diabetes on the Glycocalyx
Several pro-atherogenic stimuli affect the endothelial glycocalyx. Smoking, dyslipidemia, ischemia/reperfusion, and inflammatory cytokines all cause dimensional changes and shedding of endothelial glycocalyx components. 17,19,31,32 Already in 1983 Ceriello et al. 33 described disturbances in the metabolism of GAGs in serum of patients with diabetes, which correlated highly with longlasting hyperglycemia (glycosylated hemoglobin [A1c]). In 2005 an animal study in C57BL/6 mice showed that acute hyperglycemia increased endothelial glycocalyx permeability, most likely by affecting the hyaluronan component of the glycocalyx. 34 This was followed by a study in healthy volunteers showing that acute hyperglycemia (15 mmol/L) reduced total systemic glycocalyx volume by 50% after 6 h. 23 Similar reductions in systemic glycocalyx volume were found when a state of chronic hyperglycemia was studied in a group of patients with type 1 diabetes: systemic glycocalyx volumes were reduced by almost 50% when compared with healthy volunteers. In patients with type 1 diabetes and microalbuminuria this loss of glycocalyx volume was even more profound. 24 Likewise, patients with type 2 diabetes were found to have significantly reduced sublingual and retinal glycocalyx dimensions when compared with age-matched controls. 28 The exact mechanism by which these glucose levels lead to structural changes in the glycocalyx is still unknown. Several effects of hyperglycemia may contribute to damage to the glycocalyx.
First, the Steno hypothesis suggested that hyperglycemia influenced the sulfation of heparan sulfates in the Golgi apparatus. The subsequent studies on sulfation patterns in diabetes have mainly focused on the glomerular endothelial matrix, as this was also subject of the Steno hypothesis, and results have been conflicting. Some have reported an altered sulfation pattern of heparan sulfates in experimental diabetes, whereas others failed to confirm these findings. 35 –37 It has also been suggested that GAG size and sulfation pattern remain unchanged under hyperglycemic conditions, but the number of heparan sulfate GAG chains present on the core protein is decreased. 38
Second, the evidence of the contribution of reactive oxygen species formation to endothelial damage, including damage to the endothelial surface layer, is accumulating. It has been shown that hyperglycemia-induced damage to the endothelial glycocalyx is accompanied by the formation of reactive oxygen species and can also be partially prevented by addition of potent antioxidants.
23
Hyperglycemia leads to the formation of reactive oxygen species by mitochondrial overproduction of superoxide
Finally, it has been suggested that hyperglycemia affects the binding of hyaluronan to the glycocalyx. As described above, the main binding site for hyaluronan is CD 44, and its hyaluronan binding capacity is affected by glycosylation. It is therefore tempting to speculate that glucose levels of 10 mmol/L or higher cause increased glycosylation of CD 44 and reduce its hyaluronan binding capacity.
Although it is clear that hyperglycemia, acute or chronic, has a damaging effect on the endothelial glycocalyx, less is known about the reversibility of this damage. Antioxidant treatment has been shown to attenuate the effects of hyperglycemia, but the high doses used in these experimental settings are not suitable for clinical use. However, treatment with oral sulodexide treatment, which consists of mammalian-derived heparan and dermatan sulfates and thus offers precursor N-acetylglucosamine substrate for GAG synthesis, did result in beneficial changes in endothelial glycocalyx dimensions and metabolism of glycocalyx constituents. 28 Several other interventions have also shown beneficial effects in restoration or preservation of the glycocalyx in other disease states, such as tumor necrosis factor-α inhibition in acute inflammation, 31 statin treatment in familial hypercholesterolemia, 32 and hydrocortisone 43 and antithrombin 44 in ischemia/reperfusion damage. These interventions help clarify the pathophysiology behind glycocalyx damage in disease but are not all suitable for routine use in patients at risk, such as the patient with diabetes.
Clinical Consequences of Glycocalyx Damage
Increased vascular permeability
As the early studies into the presence of an endothelial glycocalyx have shown, the layer shows a clear size-dependent impermeability. 21 This quality contributes significantly to the barrier function of the vascular endothelium. 45 In vitro studies have shown that enzymatic degradation of the endothelial glycocalyx increases the permeability of vessels to solutes of different size. 46 This is exemplified by the role of the endothelial glycocalyx in preventing urinary albumin loss by maintaining the glomerular barrier 47,48 and thus. Endothelial glycocalyx degradation by chronic hyaluronidase infusion in apolipoprotein E-deficient mice led to proteinuria without visible changes in glomerular morphology or tubulointerstitial inflammation. 49 This supports a substantial role for the glycocalyx in upholding the glomerular barrier, even when the filtration barrier formed by the underlying tissue is intact. In diabetes, where urinary albumin loss is considered to be one of the first signals of the development of complications and a potent predictor of cardiovascular disease, it was shown that loss of systemic endothelial glycocalyx volume was most profound in patients with microalbuminuria. 24
What is more, it has been shown that removing the glycocalyx from the coronary capillary endothelial surface led to myocardial edema, which is known to contribute to cardiac dysfunction. 50 Moreover, the “leakage” of large (lipo-) proteins may also contribute to the development of atherosclerosis in patients with diabetes. In support, it was shown that a reduction in glycocalyx dimension induced by an atherogenic diet leads to intimal accumulation of low-density lipoprotein in the carotid artery 51 and transcapillary chylomicron leakage. 52
Impaired signal transduction
The transmembrane domain of the proteoglycans lining the endothelial cells makes them especially suitable for signal transduction from the intravascular lumen to the interior of the endothelial cell. In this respect, there is a specific role for the glycocalyx in shear stress sensing by the endothelial cell. This is illustrated by an impairment in the release of the vasoactive nitric oxide in response to shear stress when specific components of the glycocalyx, such as heparan sulfates 53 or hyaluronan, 54 are degraded by enzyme treatment. Also, it has been demonstrated that glycocalyx dimensions increase in areas of high flow, most likely via incorporation of hyaluronan into the glycocalyx. 55 In contrast, at sites of disturbed vascular flow, such as near arterial bifurcations, the glycocalyx dimension is significantly less. 56 These regions are characterized by a high atherogenic risk, and this supports the hypothesis that glycocalyx damage contributes to vascular vulnerability.
This is further supported by a recent study in which hyaluronan synthesis was inhibited over a period of 25 weeks in apolipoprotein E-deficient mice. This caused severe damage to the endothelial glycocalyx and markedly increased aortic plaque burden, disturbed vascular function, and had a prothrombotic as well as pro-inflammatory effect. 57 In parallel, in patients with diabetes, plasma hyaluronan concentrations were found to be a marker for angiopathy, 58 and a linear relationship between carotid intima thickness and increased level of plasma GAGs was found. 59
Inflammation and coagulation activation
The glycocalyx not only functions as a barrier to prevent the transport of solutes and proteins across the endothelium, it also shields receptor molecules such as selectins and integrins from the passing blood and prevents the adhesion of white blood cells to the endothelium. When the glycocalyx is removed or damaged, leukocyte adhesion to the endothelium quickly follows. 60 –62 In fact, the white blood cell–endothelial cell interaction in an inflammatory response occurs too swiftly to be part of up-regulation of the integrins on the endothelial surface. Rather, it will result from the uncovering of already present intercellular adhesion molecules on the endothelial cell surface. 62 This is supported by the finding of increased leukocyte–endothelium interactions in the ocular vasculature in mice lacking the heparan sulfate proteoglycan syndecan-1. 63
Similarly, the presence of the glycocalyx prevents the adhesion of activated platelets to the vascular endothelium, as a decrease in glycocalyx dimension has been shown to lead to platelet adhesion to the endothelium. 64,65
Because increased vascular permeability and a pro-atherogenic, pro-inflammatory, and pro-thrombotic state are all hallmarks of the vasculature in diabetes, it seems the endothelial glycocalyx holds special relevance for the development of cardiovascular complications in diabetes.
Future Perspective
Despite recent advances in research into the role of the endothelial glycocalyx in the development of cardiovascular disease in diabetes, several pivotal questions remain to be answered. First, the exact mechanism by which hyperglycemia damages the endothelial glycocalyx layer and its composition is still speculative. Further studies into oxidative stress-induced pathways of glycocalyx damage, altered sulfation patterns of GAGs, epigenetic changes in genes involved in turnover of glycocalyx constituents, and changed binding capacity of anchor proteins during hyperglycemia need to be evaluated to gain insight in this pathophysiological process.
Second, as of yet it is unclear whether the current measures of the endothelial glycocalyx accurately reflect the state of the glycocalyx in the cardiovascular system. Because the endothelium can adapt to the demands of the underlying tissue, 66 it is plausible that endothelial glycocalyx properties also differ depending on the tissue, and thus the sublingual or retinal vascular bed may not be representative of the cardiovascular bed.
Finally, the contribution of a damaged endothelial glycocalyx to the development of overt cardiovascular disease in diabetes needs to be clarified. Is a loss of glycocalyx merely an indicator of a diseased endothelium, a predecessor of endothelial damage, or perhaps both? And is its relevance in protecting the inner lining of the vasculature limited to the early stages of the disease, or can it play a role in delaying further damage to an already damaged cardiovascular bed in advanced diabetes?
In summary, research from the last two decades suggests an important role for the endothelial glycocalyx in upholding vascular integrity. Evidence suggests that the glycocalyx is especially vulnerable in diabetes, and thus preserving this innate protective barrier could become a new strategy in the prevention of cardiovascular complications of this increasingly prevalent disease. The focus of future research is on improving noninvasive measurement tools to reliably assess the dimension, permeability, and composition of the glycocalyx as well as large prospective trials studying the relationship between cardiovascular disease and endothelial glycocalyx perturbation in humans. Only then can the extent of its contribution to cardiovascular disease in patients with diabetes be truly known and treatment strategies to repair or preserve the layer can be critically evaluated.
Footnotes
Author Disclosure Statement
No competing financial interests exist.