
Abstract
This study evaluated quantification of fetal extracellular DNA in maternal plasma for differentiation between cases at risk of onset of placental-insufficiency-related complications and normal pregnancies. Using real-time polymerase chain reaction, fetal (sex-determining region Y [SRY] and hypermethylated RASSF1A sequence) and total (β-globin [GLO] gene) extracellular DNA was examined in 70 normal pregnancies, 18 at risk of placental-insufficiency-related pregnancy complications, 24 preeclampsia with or without (w or w/o) intrauterine growth retardation (IUGR) (median 34.0 week), and 11 IUGR (median 28.5 week). IUGR was diagnosed when estimated fetal weight was below the 10th percentile for evaluated gestational age. Although increased levels of extracellular DNA were detected in pregnancies with preeclampsia w or w/o IUGR relative to controls (RASSF1A, p < 0.001; SRY, p = 0.009; GLO, p < 0.001), quantities of fetal extracellular DNA in IUGR were not statistically significant (RASSF1A, p = 0.21; SRY, p = 0.2). RASSF1A, SRY, and GLO achieved 93.1%, 93.6%, and 92.1% accuracy for differentiation between normal pregnancy and preeclampsia w or w/o IUGR. Lower sensitivity was observed for pregnancies with onset of IUGR (RASSF1A, 60.0%; SRY, 80.0%; GLO, 72.7%), but did not influence final accuracy (RASSF1A, 91.6%; SRY, 92.5%; GLO, 89.5%). Among 18 patients at risk, 8 pregnancies involving 3 female and 5 male fetuses developed preeclampsia (n = 4), IUGR (n = 3), and chronic placentopathy causing hypoxia (n = 1). Elevation of extracellular DNA was demonstrated in 3/5 (SRY), 1/8 (hypermethylated RASSF1A), and 4/8 (GLO) patients at the earliest 26 weeks and at the latest 2 weeks before the onset of symptoms. These data indicate that fetal and total extracellular DNA concentrations can be significantly elevated in plasma of patients who later developed placental-insufficiency-related pregnancy complications. However, this is strongly individualized, and not a rule for all cases, and probably depends on the actual occurrence of excessive placental trophoblast apoptosis.
Introduction
Several groups have demonstrated the use of the multicopy DYS-14 sequence to increase the detection limit of fetal DNA in the peripheral blood of pregnant women (Sekizawa et al., 2001, 2003; Zimmermann et al., 2005, 2006; Zhong et al., 2007).
Using this approach, elevations in circulating fetal DNA concentrations have been found in a cohort of patients suffering from preeclampsia but not in pregnant women with fetal growth restriction (Sekizawa et al., 2003). The DYS-14 sequence has also been used to examine elevations of fetal DNA in the maternal circulation, before onset of symptoms, as a predictor of preeclampsia (Farina et al., 2004; Zhong et al., 2007).
Recently, Chan et al. (2006) introduced hypermethylated RASSF1A sequence as a universal fetal DNA marker and incorporated its use as a positive control, for fetal DNA detection in maternal circulation, during noninvasive prenatal fetal RHD genotyping. They demonstrated that the promotor of the RASSF1A gene was hypermethylated in the fetal part of the placenta and therefore resistant to methylation-sensitive restriction enzyme digestion, whereas maternally derived hypomethylated RASSF1A sequences were nonresistant and enzymatically digested.
Subsequently, Tsui et al. (2007) showed, in an analysis involving 10 preeclamptic and 20 normal pregnancies, a 4.3-fold higher concentration of hypermethylated RASSF1A sequences in the plasma of preeclamptic pregnancies compared with controls.
The goal of the current study was to evaluate the accuracy and potential use of hypermethylated RASSF1A, SRY, and β-globin [GLO] sequences in maternal circulation to differentiate pregnancies with the onset of placental-insufficiency-related pregnancy complications from normally progressing pregnancies in a large scale analysis.
Further, we evaluated the diagnostic potential of quantitative aberrations in hypermethylated RASSF1A, SRY, and GLO sequences in maternal circulation as risk markers for predicting late-onset placental-insufficiency-related pregnancy complications.
Materials and Methods
Patients
Local ethics committee approved the study, and informed consent was obtained from all study participants. The control cohort consisted of 70 uncomplicated singleton pregnancies sampled between the 12th and 13th week (median 12, n = 44), the 14th and 26th week (median 16, n = 61), and 27th and 36th week (median 36, n = 46) of gestation. The retrospective cohort involved 24 pregnancies with clinically established preeclampsia with or without (w or w/o) intrauterine growth retardation (IUGR) (median 34.0 week), 11 pregnancies with the onset of IUGR (median 28.5 week), and 4 women without risk, but who subsequently developed preeclampsia. Additionally, we prospectively sampled, on several occasions throughout gestation, women at risk of placental-insufficiency-related pregnancy complications based on the occurrence of preeclampsia in the patient's anamnesis (n = 10), diabetes mellitus type I (n = 2), and small size of the placenta and/or decreased pregnancy-associated plasma protein-A levels during the first trimester screening (n = 2). Gestational age was assessed using ultrasonography.
Pregnancies chosen as normal controls were not complicated by either pregnancy- or nonpregnancy-related medical problems, and concluded with births of healthy infants weighting >2500 g after 37 completed weeks of gestation.
Preeclampsia was defined as (1) blood pressure >140/90 mm Hg in two determinations 4 h apart or by a diastolic blood pressure >110 mm Hg, and (2) associated proteinuria >300 mg/24 h after 20 weeks of gestation (ACOG Committee on Practice Bulletins—Obstetrics, 2002).
IUGR is the failure to achieve the genetically predetermined growth potential and can be caused by maternal, fetal, placental, and external factors. IUGR was diagnosed when the estimated fetal weight, calculated using the Hadlock formula (Astraia Software, Munich, Germany), was below the 10th percentile for the evaluated gestational age (Hadlock, 1994); adjustments were made for the appropriate population standards of the Czech Republic.
Male fetal sex, which was determined using specific SRY real-time polymerase chain reaction (PCR) assay performed on maternal plasma samples, was confirmed postpartum.
Processing of blood samples, DNA extraction from plasma samples, and digestion by methylation-sensitive restriction enzyme
Ten milliliters of peripheral blood was collected into tubes containing ethylenediaminetetraacetic acid and processed within a few hours. Blood samples were initially centrifuged at 1200 g for 10 min, and then plasma samples were re-centrifuged (same conditions) and the supernatants were stored at −80°C until subsequent processing. DNA was extracted from plasma samples (1 mL) using a QIAamp DSP Virus Kit (Qiagen, Hilden, Germany) according to a modified version of the manufacturer's instructions for the vacuum protocol. DNA was eluted in Buffer AVE (60 μL) (Hromadnikova et al., 2006).
Plasma DNA (44 μL) was digested with 12.8 U of BstUI, a methylation-sensitive restriction enzyme (New England Biolabs, Beverly, MA), at 60°C for 16 h.
Quantification of fetal and total extracellular DNA in maternal plasma samples using SRY, RASSF1A, β-actin, and GLO as markers
Enzyme-digested plasma DNA (15 μL; hypermethylated RASSF1A sequence) and nonenzyme-treated plasma DNA (SRY sequence) were used to quantify fetal extracellular DNA. Untreated DNA (5 μL) was used to quantify total extracellular DNA (total RASSF1A, GLO, β-actin). The real-time PCR analysis was performed using a 7500 Real-Time PCR System (Applied Biosystem, Branchburg, NJ). TaqMan amplification reactions were set up in a reaction volume of 50 μL using TaqMan Universal PCR Master Mix (Applied Biosystems). Primer and probe sequences are shown in Table 1 (Lo et al., 1998; Hromadnikova et al., 2003; Zimmermann et al., 2006). Primers and probes were optimized to determine the minimum primer and probe concentrations to give the maximum Rn in monoplex (300 nM primers and 200 nM FAM-labeled TaqMan probes for SRY and GLO genes) and/or duplex assays (300 nM primers and 100 nM FAM-labeled MGB probe for RASSF1A sequence and 450 nM primers and 100 nM VIC-labeled MGB probe for the β-actin gene). Amplification was carried out in eight-well reaction optical tubes/stripes (Applied Biosystem). TaqMan PCR conditions were used as described in the TaqMan guidelines: 50 cycles at 95°C for 15 s and 60°C for 1 min, with 2 min preincubation at 50°C required for optimal AmpErase UNG activity and 10 min preincubation at 95°C required for activation of AmpliTaq Gold DNA polymerase. Each sample was analyzed at the three-replicate setting. A patient's specimen was considered positive if the amplification signal occurred on threshold cycle <40.
SRY, sex-determining region Y; GLO, β-globin.
A calibration curve (logarithmic scale), plotting the threshold cycle against known concentrations of a serially diluted DNA reference sample, was run in parallel with each analysis. Enzyme-untreated DNA extracted from the fetal part of the male placenta was used as a reference sample for quantification of SRY, RASSF1A, and β-actin sequences in maternal plasma. Without the use of methylation-sensitive restriction enzyme, both RASSF1A and β-actin sequences were detectable at all times; however, with enzyme treatment, unmethylated β-actin was completely digested and hypermethylated fetal-derived RASSF1A sequences remained intact.
The coefficient of variation for RASSF1A used as a marker for fetal extracellular DNA quantification in maternal plasma (GE/mL) did not exceed an average of 7.5% between runs.
Concentrations of RASSF1A, SRY, GLO, and β-actin were expressed as copies per milliliter of maternal plasma. The mean plus two standard deviation of fetal and total extracellular DNA levels, in age-matched normal control pregnancies, was used as the cutoff to differentiate between placental insufficiency and normal pregnancies. This cutoff showed the maximum feasible specificity (the percentage of normal pregnancies identified as not having the condition) and sensitivity (the percentage of pregnancies identified with placental-insufficiency-related complications) for SRY, RASSF1A, and GLO assays. The percentage of correct identification (accuracy for differentiation between normal and placental-insufficiency-complicated pregnancies) was calculated for each assay based on the total correct tests divided by the total tests.
Statistical analysis
Levels of extracellular fetal and total DNA in maternal plasma samples were compared using the two-tailed Student's t-test and the Mann–Whitney U-test within cohorts. Significance level was established at a p-value of <0.05. Both tests showed the same level of significance.
Results
Detection of hypermethylated RASSF1A sequences in the fetal part of the placenta and maternal plasma samples
We optimized the digestion of hypermethylated DNA in the fetal part of the placenta derived from the chorionic sac that encloses the embryo, consisting of the chorionic plate and villi, maternal plasma, and buffy coat samples.
The efficiency of enzyme digestion was monitored using duplex real-time PCR assay for RASSF1A and β-actin sequences. Without enzyme digestion, RASSF1A and β-actin sequences were always detectable, whereas after complete unmethylated β-actin digestion, only hypermethylated RASSF1A sequences were detectable.
The best efficiency of enzyme digestion was observed when 1 ng of DNA derived from the fetal part of the placenta was treated with 10 U of BstUI enzyme and 1 ng of maternal buffy coat was treated with 1 U of BstUI enzyme. In both cases, we achieved optimal results when we used 5 to 10 times higher concentrations of BstUI enzyme contrary to Chan et al. (2006).
In maternal plasma samples, optimal results (RASSF1A amplification below threshold cycle 40 and no amplification of β-actin in any replicates) were found when 10 μL of sample was digested using 2.9 U of BstUI enzyme, which corresponds to a 10 times lower concentration of BstUI enzyme than that used by Chan et al. (2006). Complete enzyme digestion was confirmed by the absence of β-actin amplification; thus, only fetal-specific hypermethylated RASSF1A sequences were detected in the fetal part of the placenta and maternal plasma samples.
Quantification of fetal and total extracellular DNA in maternal plasma samples using hypermethylated RASSF1A, SRY, and GLO sequences as markers in normal pregnancies
We quantified the extracellular DNA in maternal circulation in uncomplicated singleton pregnancies sampled between the 12th to 13th week (n = 44), the 14th to 26th week (median 16, n = 61), and the 27th to 36th week (median 36, n = 46) of gestation.
The levels of extracellular fetal DNA determined using hypermethylated RASSF1A sequences significantly increased during the progression of normal pregnancies from a median of 6.8 copies/mL at the 12th week of gestation to 12.1 copies/mL at the 16th week of gestation (p = 0.004), and 114.4 copies/mL at the 36th week of gestation (p < 0.001). In the same cohort of controls, quantities of male fetal extracellular DNA, determined using the SRY gene as the marker, showed no changes between the 12th and 16th weeks of gestation (median 42.0 vs. 41.9 copies/mL, p = 0.68); however, a significant increase during the third trimester of gestation (median 41.9 vs. 193.5 copies/mL, p < 0.001) was observed.
Similarly, a significant difference in total extracellular DNA was observed between the second and the third trimesters of gestation (median 4852.7 vs. 7126.0 copies/mL, p = 0.004), whereas quantities of total extracellular DNA during the first and second trimester of gestation stayed relatively unchanged (median 3917.2 vs. 4852.7 copies/mL, p = 0.67).
Quantification of fetal and total extracellular DNA in maternal plasma samples using hypermethylated RASSF1A, SRY, and GLO sequences as markers for the onset of placental-insufficiency-related complications
The concentrations of extracellular fetal DNA determined using either hypermethylated RASSF1A sequences (mean 693.3, median 563.5 vs. mean 107.8, median 114.4 copies/mL; p < 0.001) or the SRY gene (mean 1206.0, median 1014.6 vs. mean 233.6, median 193.5 copies/mL; p = 0.009) were significantly higher in the 24 pregnancies with clinically established preeclampsia w or w/o IUGR compared with 46 normal gestational-age-matched pregnancies (Fig. 1a, b). Similarly, a statistically significant difference in total extracellular DNA levels was observed between the studied cohorts (mean 49017.9, median 32916.3 vs. mean 7391.5, median 7126.0 copies/mL; p < 0.001) (Fig. 1c).
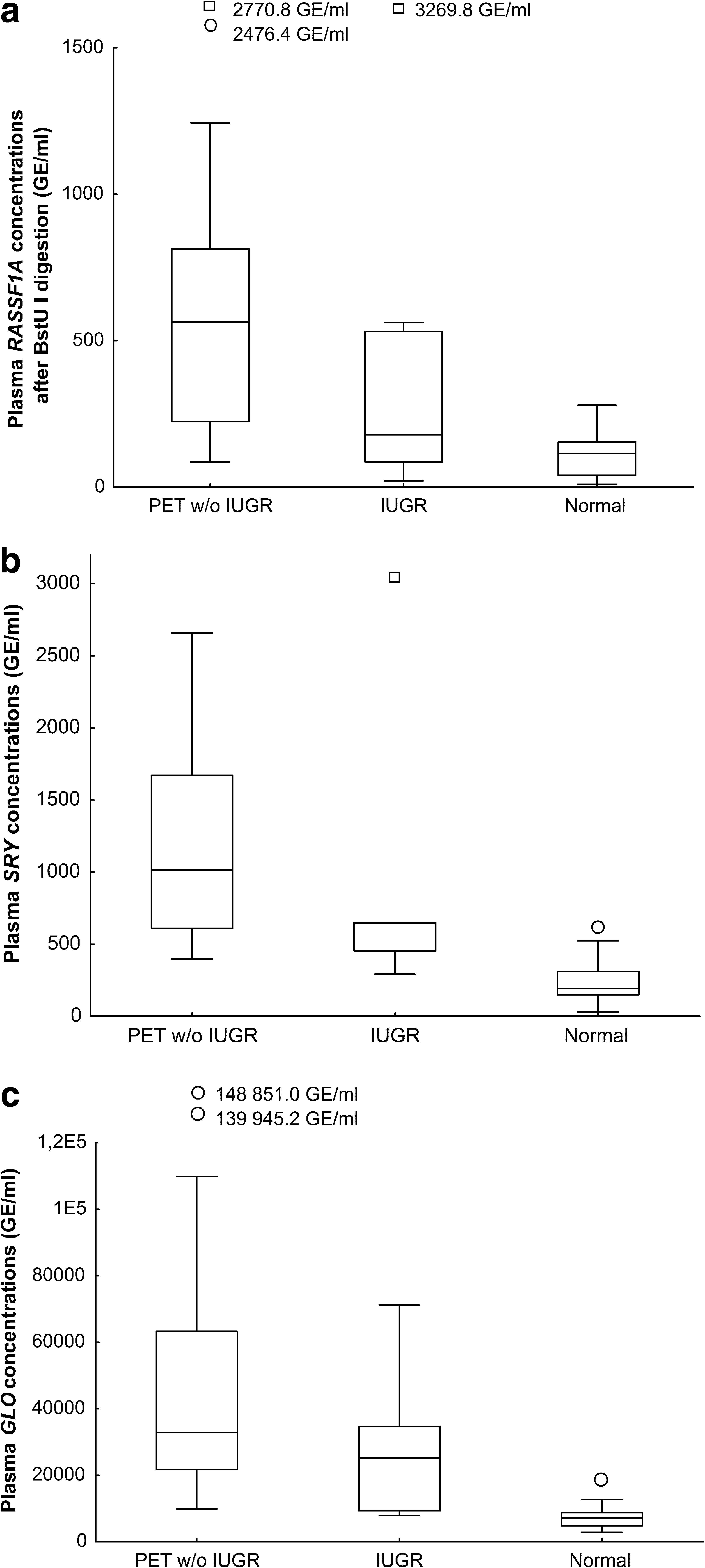
Box plots of extracellular DNA concentrations using (
The concentrations of total extracellular DNA in maternal plasma (mean 27098.5, median 25100.7 vs. mean 7391.5, median 7126.0 copies/mL; p = 0.006) were significantly increased in pregnancies during the onset of IUGR (n = 11) compared with gestational-age-matched controls (n = 46), whereas quantities of fetal extracellular DNA in maternal plasma, determined using either hypermethylated RASSF1A sequences (mean 527.6, median 179.1 vs. mean 107.84, median 114.37 copies/mL; p = 0.21) or the SRY gene (mean 1016.9, median 645.0 vs. mean 233.6, median 193.5 copies/mL; p = 0.2), did not show a statistical difference (Fig. 1).
The mean plus two standard deviation of fetal and total extracellular DNA levels in maternal plasma from normal gestational-age-matched pregnancies, which was used as a cutoff, showed 94.5%, 93.1%, and 91.3% specificity for hypermethylated RASSF1A, SRY, and GLO assays regarding differentiation between pregnancies showing the onset of placental-insufficiency-related complications and normal pregnancies. Achieved sensitivity was 86.4%, 100%, and 95.7% for hypermethylated RASSF1A, SRY, and GLO markers, which correspond to 93.1%, 93.6%, and 92.1% accuracy for differentiation between preeclampsia w or w/o IUGR and normal pregnancies, respectively. Lower sensitivity was observed in pregnancies with the onset of IUGR compared with those suffering from preeclampsia w or w/o IUGR for all examined markers (RASSF1A, 60.0%; SRY, 80.0%; GLO, 72.7%); however, this had little effect on final accuracy (RASSF1A, 91.6%; SRY, 92.5%; GLO, 89.5%).
Quantitative aberrations of extracellular DNA in maternal circulation as risk markers for predicting late-onset of placental-insufficiency-related pregnancy complications
Eight pregnancies involving three female and five male fetuses tested at various gestational stages developed placental-insufficiency-related pregnancy complications such as preeclampsia (n = 4), IUGR (n = 3), and chronic placentopathy causing hypoxia (n = 1) (Table 2).
↔, extracellular DNA concentration within the range of controls;
Fetal extracellular DNA concentrations were elevated before clinically manifested disease in three out of five (SRY sequence used as a marker) and one out of eight pregnancies (hypermethylated RASSF1A sequences used as marker). Elevation of fetal extracellular DNA (SRY sequence used as a marker) was demonstrated at 26, 23, 11, 6, 4, and 2 weeks before the onset of symptoms (unique patient number [UPN]: 1857, 2327, and 1687).
Total circulatory DNA levels were higher in four out of eight patients who later developed placental-insufficiency-related pregnancy complications. Similarly, an increase was observed at 23, 13, 11, 6, 4, and 2 weeks prior the onset of clinical symptoms (UPN: 2327, 2335, 2313, and 1687). Interestingly, fetal-derived (SRY and hypermethylated RASSF1A sequences) and total extracellular DNA (GLO) levels were persistently elevated in the plasma of a pregnant woman bearing a male fetus that was postnatally diagnosed with an undefined genetic syndrome associated with trochocephaly, hypacusia, transient anemia, and placental insufficiency leading to IUGR (UPN: 2327).
Discussion
In accordance with other authors, we confirmed significantly increased levels of fetal and total extracellular DNA in pregnancies developing preeclampsia w or w/o IUGR (Lo et al., 1999; Smid et al., 2001; Zhong et al., 2001; Sekizawa et al., 2004), suggesting an increased release of placental-trophoblast-derived apoptotic bodies into the maternal circulation.
We confirmed the data of Lo et al. (1999), who found fivefold higher median concentrations of male fetal DNA using the single copy SRY gene in plasma of preeclamptic pregnancies compared with controls; our results showed a 5.2-fold elevation of male fetal DNA in our cohort of pregnancies with established preeclampsia w or w/o IUGR compared with gestational-age-matched controls. Similarly, in the same studied cohorts we observed a 4.9-fold elevation of hypermethylated RASSF1A levels in complicated pregnancies. These data are consistent with the study of Tsui et al. (2007), who found a 4.3-fold elevation of hypermethylated RASSF1A concentrations in preeclamptic subjects compared with controls.
Although we showed statistically significant accuracy for differentiation between normal and preeclampsia w or w/o IUGR-complicated pregnancies in our cohort, results in pregnancies at the onset of fetal growth restriction were not nearly as accurate. This might be explained by the fact that an increase in fetal extracellular DNA is limited only to those pregnancies associated with placental dysfunction, and not due to other causes (Alberry and Soothill, 2008).
Further, we evaluated the accuracy and potential for using hypermethylated RASSF1A, SRY, and GLO sequences for extracellular fetal and total DNA quantification in maternal circulation with the aim of differentiating between uncomplicated pregnancies and those with the late-onset of placental-insufficiency-related pregnancy complications.
The results suggested that both fetal and total extracellular DNA concentrations might be significantly elevated in the plasma of pregnancies that later develop placental-insufficiency-related pregnancy complications. However, this is very individualized, and not a rule for all cases, and probably depends on the actual occurrence of excessive placental trophoblast apoptosis. Our finding is consistent with those reported by other groups, which observed elevations of fetal and/or total extracellular DNA in maternal plasma several weeks before the onset of symptoms (Leung et al., 2001; Zhong et al., 2002; Farina et al., 2004; Levine et al., 2004). Similarly, we detected the elevation of male fetal DNA in maternal plasma using SRY as a marker in three subjects as early as 26 weeks before clinical manifestation. Total extracellular DNA was increased in four cases as early as 23 weeks prior the onset of symptoms.
In one case (UPN: 2327) we were able to detect increased concentrations of fetal extracellular DNA in four maternal plasma samples examined on the 15th, 27th, 30th, and 34th weeks of gestation. Thus, we were able to predict the onset of placental-insufficiency-related complications starting early in the pregnancy and continuously up to shortly before the onset of severe IUGR (38th week of gestation), at which time the fetus had to be delivered by a cesarean section. Later, the newborn was found to have an as yet unclassified genetic syndrome associated with trochocephaly, hypacusia, transient anemia, and placental insufficiency resulting from severe IUGR. On the other hand, at the 32nd week of gestation, the level of fetal extracellular DNA was not increased in maternal plasma. These observations correspond with the findings of Levine et al. (2004), who demonstrated two-stage elevation of fetal extracellular DNA in patients with preeclampsia, initially between 17 and 28 weeks of gestation, thought to be due to placental necrosis and apoptosis, and again at 3 weeks before clinical symptoms, which may reflect impaired DNA elimination.
On the basis of our results we can say that differentiation between uncomplicated pregnancies and those that later develop placental-insufficiency-related pregnancy complications requires a combination of several markers to achieve the desired sensitivity with regard to detecting aberrant quantities of extracellular fetal and total DNA in maternal circulation.
Conclusion
Levels of fetal and total extracellular DNA are significantly increased in pregnancies developing preeclampsia w or w/o IUGR. Moreover, concentrations of fetal and total extracellular DNA might also be elevated in the plasma of patients who later develop placental-insufficiency-related complications. However, this is strongly individualized, and not a rule for all cases, and probably depends on the actual occurrence of excessive placental trophoblast apoptosis.
Footnotes
Acknowledgments
We thank the staff of the Clinic of Obstetrics and Gynecology and the Institute for the Care of Mother and Child for their participation in the collection of data. We would also like to thank Prof. Y.M. Dennis Lo from the Chinese University of Hong Kong, Prince of Wales Hospital, Shatin, Hong Kong, for his valuable suggestions concerning digestion of extracellular DNA by methylation-sensitive restriction enzyme. This work was supported by the grant project MSM 0021620806.
Disclosure Statement
No competing financial interests exist.