
Abstract
Adenine and uridine (AU)–rich elements in the 3′ untranslated region (3′UTR) have been implicated in the 17β-estradiol (E2) stabilization of vertebrate estrogen receptor (ER) mRNAs. To date, fishes have the most complex arrangement of nuclear ERs with up to two isoforms of each of the two genes in some species (i.e., four different ERs). The objective of this study was to analyze the sequence variation of 3′UTRs among the four ER isoforms in the rainbow trout and determine to what degree it is responsible for the estrogen-induced increase of ER mRNAs in the liver of this fish. This was done by comparing the 3′UTR DNA sequence length and composition, and by measuring expression of ER isoform 3′UTR luciferase reporter constructs in primary cultures of trout hepatocytes treated with E2. There were large differences both in overall length and in sequence composition among the four ER isoform 3′UTRs. The ERα1 sequence was the longest and the only one of the four that contained multiple copies of the canonical AU-rich elements (AUUUA) as well as the stability sequence (GCUGAU). E2 treatment significantly increased the luciferase activity in cells transiently transfected with the ERα1 reporter construct, relative to cells transfected with reporter vectors containing the other three ER isoform 3′UTRs or the parental vector control. These results support the hypothesis that the E2-induced increase in hepatic ERα1 mRNA in rainbow trout is due in part to sequence variability among ER isoform 3′UTRs. We conclude that posttranscriptional stabilization of ER mRNA by E2 appears to be conserved among vertebrates.
Introduction
Posttranscriptional regulation of unstable mRNAs is now recognized as a critical mechanism for controlling gene expression in vertebrates from fish (Roca et al., 2007) to mammals (Annamaria et al., 2003). Messenger RNA stabilization involves the action of specific cis-acting sequences and trans-acting factors (Xu et al., 1997 and references therein). The cis-acting sequences that have received the most attention are the adenine and uridine (AU)–rich elements (AREs) found in the 3′UTRs of many unstable mRNAs. These are sequence motifs often occurring as multiple copies, which usually include the pentameric sequence AUUUA (Caput et al., 1986). The first direct evidence that this ARE functions as an instability element was demonstrated by inserting a 50 nucleotide sequence containing multiple copies of this ARE into the 3′UTR of the human β-globin mRNA, which markedly reduced the stability of this mRNA (Shaw and Kamen, 1986). Another 3′UTR sequence motif, mediating albumin mRNA stability in frog liver, is A[C/U]UGA (APyrUGA), which is recognized and degraded by the estrogen-induced polysomal endoribonuclease 1 (Margot and Williams, 1996; Chernokalskaya et al., 1997). Conversely, a recent deletion mutant analysis of sheep ER mRNA identified two minimal E2-modulated stability sequences (MEMSSs) that confer stability to a heterologous RNA in a cell-free system (Mitchell and Ing, 2003). These MEMSSs are approximately 80 nt long and each contains one copy of a 10 base sequence (UGUAUUCUUC). These sequences also contain APyrUGA-related sequences, indicating that motifs associated with mRNA destabilization can also function to stabilize mRNAs.
The complete nuclear ER family in rainbow trout consists of two subtypes (ERα and ERβ) each of which consists of two isoforms (α1/α2 and β1/β2). Others and we have shown previously that E2 treatment specifically increases the amount of ERα1 mRNA (Pakdel et al., 1989, 1991; MacKay et al., 1996) during early embryonic development (Boyce-Derricott et al., 2010) and in the sexually immature trout liver (Boyce-Derricott et al., 2009), whereas the level of the other three isoforms does not change. The objectives of this study were (1) to analyze the sequence composition of the four rainbow trout ER isoform 3′UTRs and (2) to determine if variation among those four 3′UTR sequences is responsible for the estrogen-induced increase in ERα1 mRNA observed in the liver. For the first objective, the number and location of specific sequence motifs previously reported in other animals to be involved in mRNA stability/instability were identified and compared with those in the rainbow trout. For the second objective, we tested the hypothesis that in trout hepatocytes treated with E2 the 3′UTR derived from ERα1 mRNA would confer stability to a heterologous mRNA transcribed from a luciferase reporter construct.
Materials and Methods
Chemicals
E2 and all other cell culture and bacterial selection reagents except for serum-free Leibovitz's L-15 were purchased from Sigma Chemical Co. (St. Louis, MO). Leibovitz's L-15 was purchased from Gibco BRL (Grand Island, NY). Restriction enzymes (FseI and XbaI) were purchased from New England Biolabs (Ipswich, MA). Type IV collagenase was purchased from Worthington Biochemical Co. (Lakewood, NJ).
Generation of reporter constructs
Each of the four 3′UTRs corresponding to the four ER isoforms were amplified by PCR from rainbow trout genomic DNA. Primers used to amplify the ERα1 3′UTR (Table 1) were based on DNA sequence data initially published in Pakdel et al. (1990). Primers used to amplify the 3′UTRs from the other three isoforms (ERα2, ERβ1/β2) were based on sequences deposited in GenBank and reported in Nagler et al. (2007). Each 3′UTR was sub-cloned into either the FseI site (ERα1) or directionally inserted into the FseI and XbaI sites (ERα2 and ERβ1/β2) of the firefly luciferase reporter vector pGL-4.13 SV40/luc (Promega, Madison, WI). Each of these reporter vectors containing a different ER 3′UTR is controlled by the same constitutively active SV40 promoter.
FseI and XbaI restriction sites used to clone the ER 3′ untranslated regions into the pGL-4.13 vector are underlined.
ER isoform 3′UTR sequence analysis
Each ER isoform 3′UTR construct was sequenced using an ABI 3100 automated sequencer. All sequence data were analyzed by eye and with CLC Main Workbench (2009). Specific sequence motifs included in the search were the vitellogenin mRNA stabilizing element (ACUGAU), the albumin mRNA stabilizing elements (APyUGA), and AREs by comparison to frog (vitellogenin [VTG] and albumin mRNA) and sheep and human (ERα mRNA) sequences.
Hepatocyte isolation
A primary hepatocyte culture was established from one isogenic male rainbow trout weighing approximately 150 g maintained in a recirculating tank with a photoperiod of 12 h light:12 h dark and a temperature of 11°C, and was fed a commercial fish food. Before hepatocyte isolation the trout was deeply anesthetized with tricaine methanesulfonate (MS-222) using a two-step perfusion method as described previously (Mommsen et al., 1994). Briefly, the liver was perfused in situ with modified Hank's medium (136.9 mM NaCl, 5.4 mM KCl, 0.81 mM MgSO4, 0.44 mM KH2PO4, 0.33 mM Na2HPO4, 5.0 mM NaHCO3, and 10 mM HEPES, pH 7.63) for 10–15 min to remove blood. The liver was then perfused for 30 min with the same solution containing type IV collagenase to digest the liver tissue. The liver was then removed and cells were dispersed in ice-cold modified Hank's medium containing 1.5 mM CaCl2 and 2% BSA, filtered through 250 and 75 μm nylon mesh and collected by centrifugation at 60 g for 3–4 min. The cells were then allowed to settle on ice for 20 min in L15 medium containing 450 mM NaHCO3 and 1× penicillin–streptomycin. After this settling period the cells were resuspended in maintenance medium, which consisted of serum-free Leibovitz's L-15 containing antibiotics. Cell viability was assessed by Trypan Blue exclusion and cells were counted with a hemacytometer. The cells used in this experiment were greater than 90% viable.
Cell culture and transfection
Cells were seeded in 24-well (350,000 cells/well) plates (Becton-Dickson Labware, Lincoln Park, NJ) in antibiotic-free L-15 medium and maintained at 11°C for 24 h before transfection and treatment. Cells were transfected using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's specifications. Cells were cotransfected for 2 h in 500 μL antibiotic-free L-15 medium containing 2 μL transfection reagent and 0.5 μg of one of the four pGL-4.13SV40 luciferase-ER-3′UTR constructs and 0.05 μg of the pRL-CMV plasmid (Promega) used to determine transfection efficiency. Plasmid DNA from all constructs was obtained using a mini-prep kit (Qiagen, Inc., Alameda, CA) and amplified in transformed Escherichia coli (DH5α strain) grown in LB medium containing 50 μg/mL penicillin G.
Cell treatments
Cells were treated with vehicle control (0.001% ethanol) or 10 μM E2. Although this dose of E2 is above physiological levels, we have reported in a previously published experiment (Boyce-Derricott et al., 2009b) that 10 μM E2 induces a maximal ER transcriptional response in trout hepatocyte cell lines. After addition of treatment medium, the plates were covered with a breathable membrane (Diversified Biotech, Boston, MA) to reduce evaporative losses and any cross-well contamination. Assays to detect plasmid DNA expression were conducted 24 h after the cells were treated, and the treatment medium was not changed over the course of the experiment.
Luciferase assays
Measurement of firefly luciferase activity was obtained at room temperature using the Dual-Luciferase™ assay kit (Promega) and a single tube luminometer (Model LB 9507; EG&G Berthold, Bad Wilbad, Germany). At the end of the treatment period (24 h) cells were lysed with 100 μL of passive lysis buffer for 20 min at room temperature on an orbital shaker, and the resulting lysate was transferred to a microcentrifuge tube and spun down briefly to pellet any cellular debris. For each sample, 20 μL of cell lysate was added to a glass luminometer tube that contained 100 μL of the firefly luciferase assay reagent and mixed gently. This was followed by a 10-s integrated measurement of the luminescent signal. Addition of 100 μL of Stop & Glo™ reagent quenched the firefly luciferase activity, followed by a 10-s measurement of the luminescent signal from the pRL-CMV luciferase.
Statistical analysis
The statistical analysis was conducted within the SAS analysis package (SAS Institute, Cary, NC). Relative luciferase units (RLU) were calculated by dividing the firefly luciferase values by the Renilla luciferase values and were analyzed by least-squares ANOVA using the general linear model procedure and are presented as least-squares means and standard errors for treatment groups. Overall significant effects were tested with Tukey's post hoc multiple range test. Significance was set at p < 0.05.
Results
ER isoform 3′UTR sequence analysis
The trout ERα1 3′UTR (1313 nt) is longer than that of ERα2 (105 nt), ERβ1 (261 nt), or ERβ2 (405 nt). The ERα1 3′UTR contained the stability element (GCUGAU), two copies of the stability element (ACUUUU), and one copy of the stability element (ACUUCU). The ERα1 sequence also contained one copy of the instability element (GAUGAU), nine copies of the canonical ARE instability element (AUUUA), and eight copies of the frog albumin instability element APyrUGA. The ERα2 3′UTR contained no sequence elements reported to be involved in mRNA stability/instability. The ERβ1 3′UTR sequence contained one copy of the stability element (ACUUCU) and one copy of the instability element (AUUUA). The ERβ2 3′UTR sequence contained one copy each of the stability elements (ACUUCU) and (ACUUUU) and three copies of the frog albumin instability element (APyrUGA).
ER 3′UTR luciferase assays
Least-squares ANOVA detected a significant effect of treatment on RLU. E2 treatment significantly increased the RLU in hepatocytes transiently transfected with the ERα1-3′UTR construct over those treated with vehicle control. The mean RLU in all other groups were not different from the vector control group (Fig. 1).
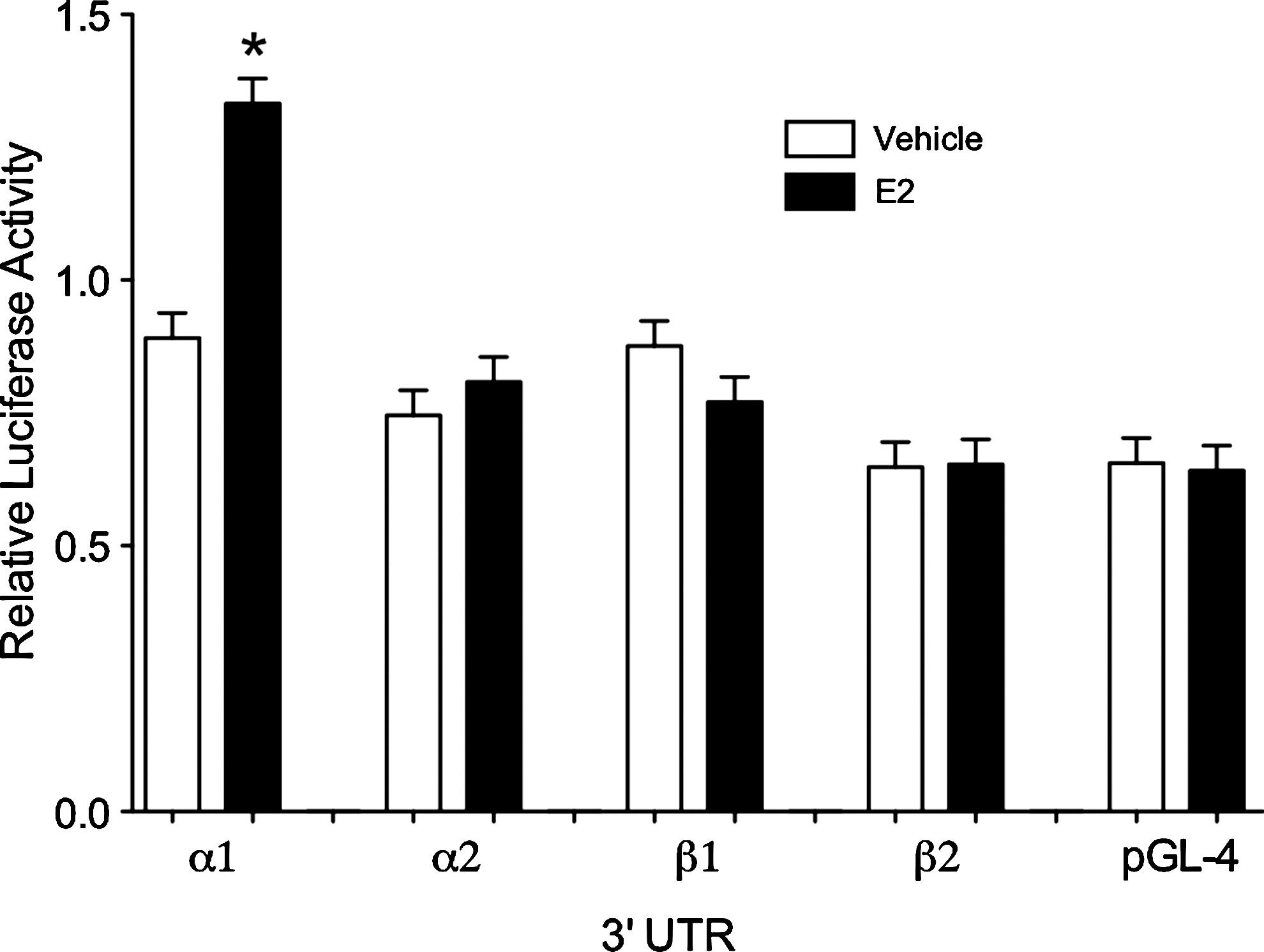
Relative luciferase activity in primary cultures of rainbow trout hepatocytes transfected with one of the four estrogen receptor (ER) isoform 3′ untranslated region constructs in the presence (closed bars) or absence (open bars) of 10 μM 17β-estradiol. The asterisk represents a statistical difference according to Tukey's test (p < 0.05).
Discussion
This study has generated data that supports the hypothesis that variation in length and/or sequence composition among rainbow trout ER isoform 3′UTRs mediates the observed E2-induced increase in hepatic ERα1 mRNA levels. The four ER isoform 3′UTRs were extremely variable both in overall length and sequence element composition (Fig. 2). There were numerous, highly conserved sequence elements common to frog, sheep, and human 3′UTRs (Dodson and Shapiro, 1994; Robertson et al., 2002; Mitchell and Ing, 2003), the majority of which were found within the ERα1 3′UTR. There were also numerous insertions and deletions in the ER isoform 3′UTR sequences we obtained by PCR as compared to those initially reported for ERα1 in Pakdel et al. (1990) and for the other three isoforms reported in Nagler et al. (2007); however, none of these polymorphisms were within the sequence motifs of interest. Interestingly, the presence of some of these motifs in the two ERβ isoforms does not appear to affect the stability of these mRNAs, suggesting that sequence-based stability may arise not only from the type of sequence motif present but also from the copy number and location of those sequence motifs (Chen and Shyu, 1995; Meisner et al., 2004; Zhao et al., 2008b).
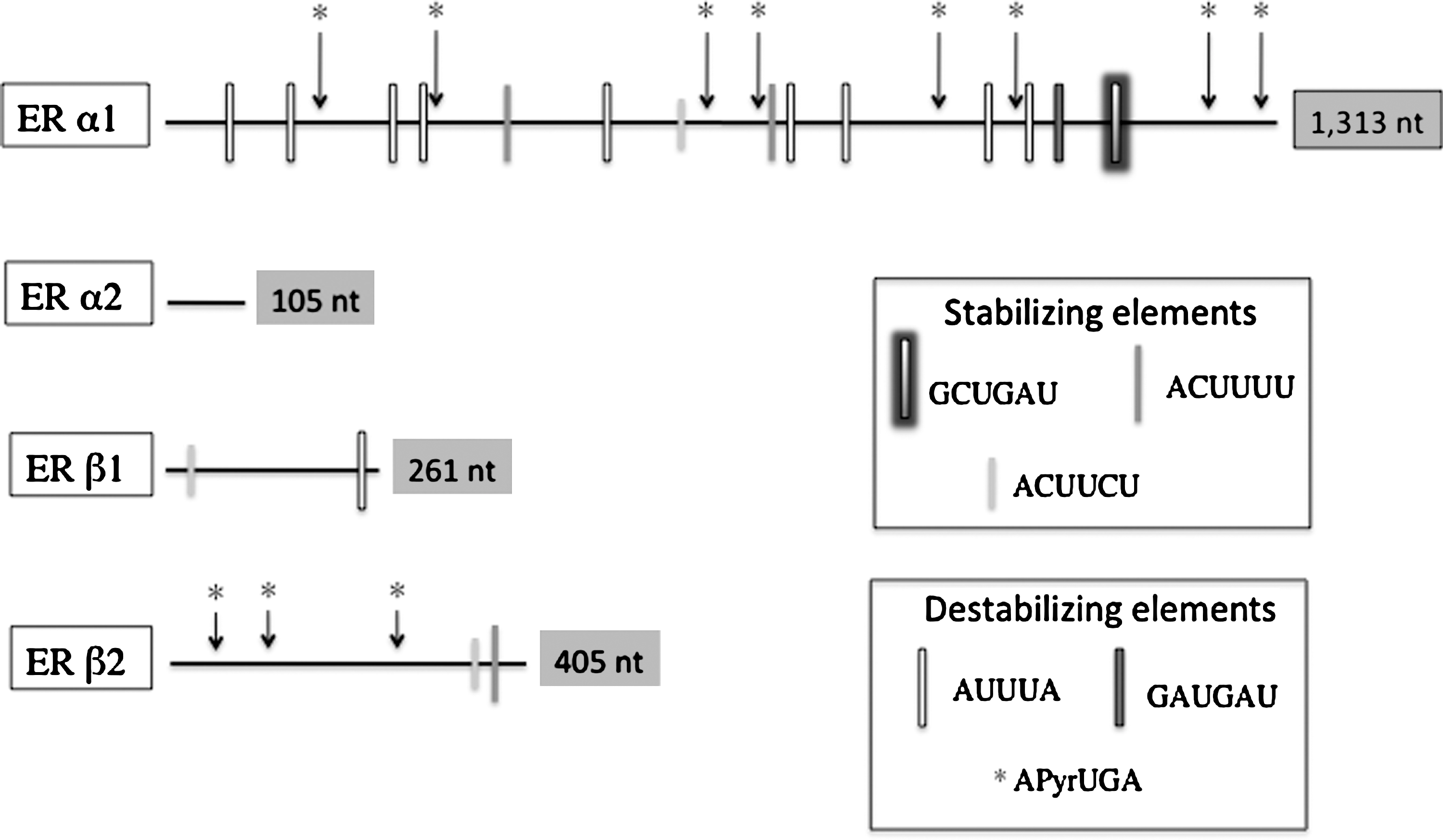
Schematic representation of the four (ERα1, ERα2, ERβ1, and ERβ2) rainbow trout ER isoform 3′ untranslated regions showing the variation in length and distribution of adenine and uridine (AU)–rich elements and other putative cis-elements.
Through extensive analyses of deletion mutants and stable structure predictions using RNA folding algorithms, Mitchell and Ing (2003) identified four MEMSSs in the sheep ERα mRNA 3′UTR that appear to be important in the observed E2-induced increase in endometrial ER mRNA stability. Further, it was observed that, of the four MEMSSs, two could confer E2-associated stability to a heterologous RNA. These two sequences were approximately 80 nt long and shared a 10-base uridine-rich segment (UGUAUUCUUC). The predicted structures of these MEMSSs include single-stranded regions, which contain the 10-base U-rich sequence and double-stranded regions, which include multiple copies of the APyUGA motif. This 10 base U-rich sequence was not found in the any of the four trout 3′UTR sequences, which suggests that the other sequence motifs may be involved in the E2-enhanced stability reported here.
Long 3′UTR sequences increase the number of potential sites for RNA binding proteins, including endonucleases (Mitchell and Ing, 2003). The extensive 3′UTRs of mRNAs of frog vitellogenin and a number of mammalian nuclear hormone receptors appear designed to be unstable, carrying large numbers of ARE instability elements (Ing, 2005). In contrast, stable mRNAs such as β-globin and glyceraldehyde 3-phosphate dehydrogenase are shorter and contain fewer elements (Mitchell and Ing, 2003). The trout ERα1 3′UTR is much longer than that of ERα2, ERβ1, or ERβ2 and contains many more ARE sequence motifs. The ERα1 sequence also appears to cross the boundaries recently described for an ARE classification system (Xu et al., 1997). This system groups sequences according to ARE copy number, suggesting that class I sequences contain 1–3 copies of the AUUUA pentamer and are typical of nuclear transcription factors such as ER; however, the trout ERα1 sequence has nine copies of this motif.
The regulation of trout ER isoform expression could involve a combination of processes acting at the transcriptional, posttranscriptional, and posttranslational levels as has been described in mammals (Nawaz et al., 1999; Reid et al., 2002; Ing et al., 2008). Recent evidence indicates a role for noncoding RNAs in the regulation of steroid hormone receptor expression (Hayes et al., 2006; Varghese and Cohen, 2007), including the human ERα (Adams et al., 2007; Zhao et al., 2008a; Liu et al., 2009). Noncoding RNAs such as small interfering RNAs (siRNA) and microRNAs (miRNA) regulate gene expression by directing the degradation and/or translational repression of many classes of mammalian mRNAs involved in growth (Elia et al., 2009), differentiation (Lize et al., 2009), and reproduction (Nagaraja et al., 2008). One of the critical components of this genetic regulatory system is the ribonuclease III enzyme, Dicer, which is critical for processing of miRNAs and synthesis of siRNAs from long double-stranded RNA (reviewed in Shyu et al., 2008; Wu and Belasco, 2008). Recent reports have shown that miRNA biogenesis is affected by estrogens (reviewed in Klinge, 2009) and that the miRNA, mir-22, negatively regulates human ERα expression by binding to its 3′UTR (Pandey and Picard, 2009). These authors also showed that the human ERβ isoform is not affected by this miRNA because the recognition sequence necessary for binding is not present in its 3′UTR. In the trout liver we have observed that E2 treatment induces a large increase in the level of Dicer mRNA (unpublished data), thus possibly expanding the potential mechanisms of posttranscriptional regulation of nuclear hormone receptors in fish.
The mechanism of E2-related posttranscriptional mRNA regulation identified for amphibian vitellogenin (Dodson and Shapiro, 1994, 1997) apolipoprotein II (apoII) expression in the chicken liver (Ratnasabapathy, 1995), and ERα expression in the mammalian uterus (Ing et al., 1996, 2008; Robertson et al., 2002; Mitchell and Ing, 2003) appears conserved, because in all of these an RNA-binding protein has been implicated as the trans-acting factor mediating the E2-related stabilization process. Amphibian vitellogenin mRNA is protected from cleavage by the protein vigilin, and avian apoII mRNA is stabilized by an estrogen-regulated mRNA stabilizing factor (E-RmRNASF). Sheep endometrial extracts contain an E2-induced protein, which requires Mg2+ and reducing conditions to exert its stabilizing effect zon ER mRNA (Mitchell and Ing, 2003) and appears to be an AU-rich RNA-binding factor 1 isoform (Ing et al., 2008). Currently, there is no information concerning a possible trans-acting factor(s) responsible for stabilization of rainbow trout ER isoform mRNAs; however, work in the experimental systems described above provide a framework for elucidating the identity of such factors in the future.
Conclusions
The extensive sequence variation, both composition and length, observed among the four ER isoform 3′UTRs appears to mediate the observed effect of E2 on the stability of hepatic ERα1 mRNA in rainbow trout. More research is needed to elucidate the molecular mechanisms responsible for this E2-enhanced stability—in particular, the cis- and trans-acting factors, presumably specific sequence motifs and one or more RNA binding proteins, involved in the posttranscriptional regulation of this family of nuclear ERs. These experiments could include measuring endogenous ER mRNA stability in cultured hepatocytes treated with E2 and a transcriptional inhibitor such as actinomycin D and mutational analyses to test for the effect of sequence length and composition on mRNA stability. Finally, a bioinformatic analysis of ER 3′UTRs to identify miRNA recognition sequences could provide insight into the possibility that ER expression is regulated by noncoding RNAs in the rainbow trout.
Footnotes
Acknowledgment
This work was partially supported by National Institute of Environmental Health Sciences Grant ESO12446-5.
Disclosure Statement
No competing financial interests exist.