
Abstract
Biofilm formation plays a crucial role in the development of different infections. This study was designed to examine the effects of extracellular DNA destruction by DNase I on characteristics of forming bacterial biofilms. We have found that extracellular matrix of biofilms formed in the presence of DNase I contains extracellular DNA fragments of about 30 kb. These data support the idea that cell-free DNA is constantly released to the extracellular matrix of bacterial biofilms. Our results indicate that extracellular DNA plays an important role in the properties of forming biofilms. Biofilms formed in the presence of DNase I (5.0 μg/mL) displayed reduced biofilm biomass, total bacterial biomass, decreased viability of bacteria, and decreased tolerance to antibiotics. The fact that destruction of extracellular DNA in forming biofilms by DNase I leads to the formation of an altered microbial community with decreased tolerance to environmental factors suggests the possibility to change the characteristics of forming biofilms by modifying cell-free DNA.
Introduction
In this article, we report the effect of extracellular DNA destruction starting from the beginning of biofilm formation on the characteristics of forming microbial communities.
Materials and Methods
Bacterial strains
Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 29213 were used in this study.
Medium and culture conditions
The liquid media used for bacterial growth were Luria–Bertani (Becton Dickinson, Sparks, MD) and Mueller–Hinton (bioMerieux, Paris, France). The strains were grown at 37°C, and liquid cultures were incubated without shaking. Before use in biofilm experiments, cells were harvested and washed twice with 0.15 M isotonic phosphate-buffered saline (pH 7.2), and the cell suspensions were standardized to an optical density (OD) of 0.8 at 520 nm.
Enzymes
Bovine pancreatic deoxyribonuclease I (Sigma Chemical, St. Louis, MO), with a specific activity of 2200 Kunitz units/mg was used.
Antibiotics
The antibiotics tested were ampicillin (ICN, Costa Mesa, CA) and levofloxacin (Sigma Chemical).
Planktonic antimicrobial susceptibility testing
MIC tests were performed in cation-adjusted Mueller–Hinton broth according to Clinical and Laboratory Standards Institute guidelines for broth microdilution susceptibility testing (NCCLS, 1999; CLSI, 2006).
Biofilm formation assay
An inoculum was prepared using a 24-h broth culture. The inoculum, which contained 7.53 ± 0.22 log10 CFU/mL, was added to the wells of 96-well plates (200 μL/well), 35-mm Petri dishes (2 mL), and coverslips that were placed in glass tubes (2 mL) (all from Sarstedt, Numbrecht, Germany). Then the plates, dishes, and coverslips were incubated for 24 h at 37°C.
Biofilm CFU assay
The term “the number of CFU” means the number of viable bacteria that grow on nutrient media in the current conditions. Biofilms were grown in 96-well plates for 24 h at 37°C. Liquid medium with bacteria was aspirated from the wells, which were then washed with isotonic phosphate buffer (0.15 M, pH 7.2). Biofilms were scraped thoroughly, with particular attention to well edges. The well contents were aspirated again, then placed in 1.0 mL of isotonic phosphate buffer (0.15 M, pH 7.2), and dispersed by drawing up and down through a fine-tip pipette, and the total CFU number was determined by serial dilution method and plating on appropriate media.
Spectrophotometric quantification of total bacterial biomass
The term “total bacterial biomass” is defined as the total biomass of all bacteria independent of their state of activity: viable (culturable) (Prosser et al., 1987; Oliver, 2005), viable but nonculturable (Juhna et al., 2007; Muela et al., 2008), or dead bacteria. Bacterial biomass is determined by the number of bacteria and bacterial size.
Quantification of bacteria in biofilms was performed by spectrophotometric method. Liquid medium with bacteria was aspirated from the wells, and the wells were then washed with isotonic phosphate buffer (0.15 M, pH 7.2) without disturbing the adherent film. Biofilm cells were scraped off the well walls and resuspended in 100 μL of phosphate buffer (0.15 M, pH 7.2). The well contents were then aspirated and bacteria were harvested by 10 min of centrifugation at 5000 rpm (Eppendorf 5415 C centrifuge; Eppendorf Geratgebau GmbH, Hamburg, Germany). The pelleted bacteria were diluted in 2 mL PBS, and absorption was determined at 570 nm in a LKB Biochrom Novaspec 4049 spectrophotometer (Cambridge, United Kingdom).
Quantitative determination of biofilm formation
Quantitative determination of biofilm formation was performed by a spectrophotometric method that measures the total biofilm biomass, including bacterial cells and EPS. The term “biofilm biomass” means the total biofilm biomass, including bacterial cells and EPS, determined by the spectrophotometric method.
Liquid medium with bacteria was aspirated from the wells, and the wells were then washed with isotonic phosphate buffer (0.15 M, pH 7.2) without disturbing the adherent film. Biofilms were stained with 200 μL of 0.1% crystal violet, incubated at room temperature for 30 min, and excess stain was removed by three gentle washes with sterile distilled water. After being dried, the crystal violet was solubilized by adding 200 μL of ethanol–acetone (80:20, wt/wt) and the extent of biofilm was determined by measuring the absorbance of the stained adherent film with a microplate reader (Stat-Fax-2100; Awareness Technology, Palm City, FL) at 570 nm.
Isolation of extracellular DNA
Biofilm formation was carried out in Petri dishes as described in biofilm formation assay for 24 h at 37°C. The supernatant fluid was aspirated and biofilms were gently washed three times with isotonic phosphate buffer (0.15 M, pH 7.2) without disturbing the adherent film. Biofilms were scraped from the Petri dishes in the presence of 0.5 mL of isotonic phosphate buffer (0.15 M, pH 7.2) and biofilm matrix was separated from bacteria by centrifugation at 5000 g for 10 min (Eppendorf 5415 C centrifuge, Eppendorf Geratgebau GmbH). Extracellular DNA was extracted from the supernatant solution containing biofilm matrix with an AquaPure genomic DNA isolation kit (Bio-Rad Laboratories, Hemel Hempstead, United Kingdom), following the manufacturer's instructions.
Electrophoresis
The size of extracted DNA was ascertained by electrophoresis on a 1.0% agarose gel (ICN) in the presence of 0.5 μg/mL of ethidium bromide that was used for DNA staining and visualized under ultraviolet light (UV Transilluminator 2000; Bio-Rad Laboratories).
Effect of DNase I on biofilm formation
Biofilm formation was carried out in 96-well plates as described for the biofilm formation assay. Nutrient medium was supplemented with DNase I 5.0 μg/mL. After 24 h of incubation at 37°C, solutions were discarded and wells were filled (200 μL) with fresh media and the number of CFU was counted or quantitative determination of biofilm formation or total bacterial biomass was carried out.
Effect of antibiotics on 24-h-old biofilms formed without DNase I
Biofilm formation was carried out as described for the biofilm formation assay. After 24 h of incubation at 37°C, solutions were discarded and wells were filled (200 μL) with fresh media supplemented with correspondent antibiotic at 50 times the MIC. After 24 h of additional incubation at 37°C, biofilm characteristics were investigated.
Effect of antibiotics on 24-h-old biofilms formed in the presence of DNase I 5.0 μg/mL
Biofilm formation in the presence of DNase I 5.0 μg/mL was carried out as described for the biofilm formation assay All media were supplemented with DNase I 5.0 μg/mL. After 24 h of incubation at 37°C, solutions were discarded and wells were filled (200 μL) with fresh media supplemented with correspondent antibiotic at 50 times the MIC. After 24 h of additional incubation at 37°C, the number of CFU was counted or quantitative determination of biofilm biomass or total bacterial biomass was carried out.
Exonuclease activity assay of DNase I solutions
Exonuclease activity was determined by incubating 1.0 μg plasmid (pVAX1; Invitrogen, Carlsbad, CA) with 19.0 μL of cell-free extracellular biofilm matrix, followed by incubation for 30 min at 37°C with subsequent analysis by 1.0% agarose gel electrophoresis.
Statistical analysis
All statistic analyses were performed using the statistics package Statistica for Windows (version 5.0). A p-value of <0.05 was considered statistically significant.
Results
Confirmation of the constant release of extracellular DNA by bacteria and verification of permanent existence of extracellular DNA in matrix of biofilms formed in the presence of DNase I
Extracellular DNA fragments with molecular sizes of about 30 kb were found in the extracellular matrix of all bacterial biofilms tested at time 2, 4, 8, 12, and 24 h of biofilm growth, which were formed in the presence or absence of DNase I, by using DNA isolation kit (Bio-Rad Laboratories). We have previously confirmed that extracellular polymeric molecules are DNA fragments by using DNase, RNase, and proteinase treatments with subsequent analysis by gel electrophoresis (Tetz et al., 2009). The results indicate that the extracellular matrix of 2, 4, 8, 12, and 24-h-old bacterial biofilms formed by unrelated gram-positive and gram-negative bacteria in the presence or absence of DNase I contains detectable DNA fragments of about 30 kb.
Confirmation of the extracellular DNA sensitivity to DNase I
To confirm that DNase I at 5.0 μg/mL effectively cleaves cell-free DNA in biofilms, we investigated its effect on the extracellular matrix of E. coli and S. aureus biofilms. Cell-free extracellular matrix of E. coli and S. aureus biofilms was obtained as described earlier. Control samples were left untreated, and experimental samples of the extracellular matrix were treated with DNase I 5.0 μg/mL and then incubated for 30 min at 37°C. Extracellular DNA was extracted using DNA extraction kit with subsequent analysis by 1.0% agarose gel electrophoresis. The results revealed complete degradation of 30 kb band using DNase 5.0 μg/mL, compared with untreated control. Thus, addition of DNase I at 5.0 μg/mL to the cell-free biofilm matrix of gram-positive and gram-negative bacteria effectively cleaves extracellular DNA.
Confirmation of existence of exonuclease activity of biofilm matrix
To confirm that matrix of biofilms formed in the presence of DNase I at 5.0 μg/mL retains exonuclease activity after 2, 4, 8, 12, and 24 h of biofilm cultivation, we studied its activity as described in exonuclease activity assay. Matrix of 2, 4, 8, 12, and 24-h-old biofilms displayed potent exonuclease activity.
Effect of extracellular DNA destruction on biofilm characteristics
The effect of extracellular DNA destruction by different concentrations of DNase I on characteristics of forming biofilms was studied using light microscopy, spectrophotometric measurements of total bacterial biomass and biofilm biomass, and determination of the CFU number. On examination by light microscopy, the number of microcolonies and their sizes in biofilms formed in the presence of DNase I at 1.0 and 5.0 μg/mL was reduced when compared with untreated control. The architecture of biofilms treated with DNase I differs from that of untreated control. Bacterial cells within DNase-treated biofilms form mesh-like structures with increased area of cell-free zones.
Various concentrations of DNase I enzyme were simultaneously inoculated together with 24-h bacterial broth culture in a 96-well plate as described in the biofilm formation assay. Plates were incubated for 24 h at 37°C with subsequent spectrophotometric determination of biomass and comparison between biomass of treated and untreated biofilms.
The results of five independent experiments revealed a DNase concentration-dependent reduction in biofilm biomass, indicating that DNase I has a consistent effect on biofilm formation of various bacteria (Table 1).
The results are representative of those from three to four separate experiments.
OD, optical density.
Addition of DNase I at 5.0 μg/mL to developing biofilms at time zero decreased the biomasses of the E. coli and S. aureus biofilms by 54% and 51% (Fig. 1). The numbers of CFU determined for strains tested were found to be two- to threefold reduced (Table 2).
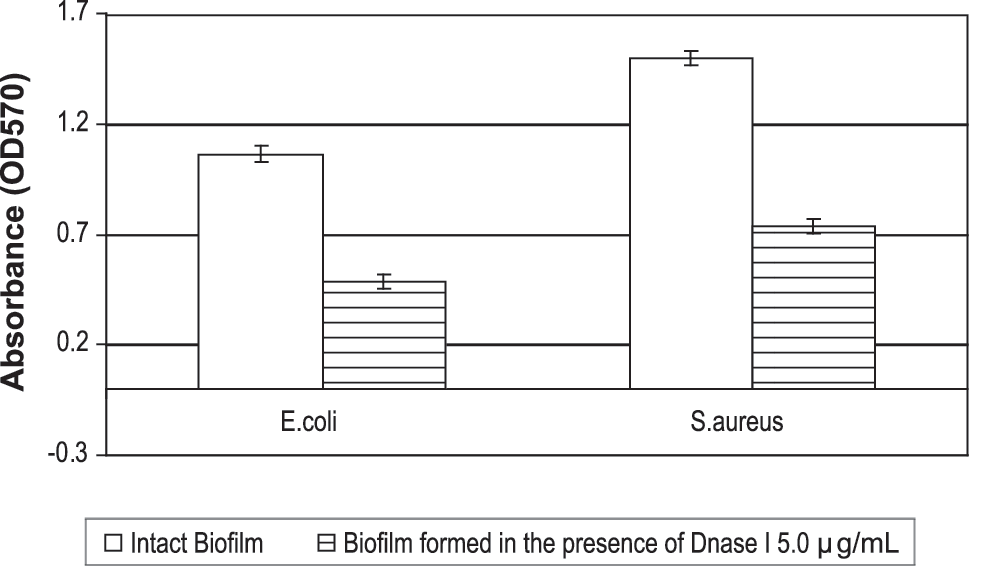
Quantitative biofilm analysis of the effect of the extracellular DNA destruction by DNase I 5.0 μg/mL. Biofilms were formed in the presence or absence of DNase I 5.0 μg/mL for 24 h at 37°C. The bacteria adhering to microtiter plates were stained and the absorbance was measured as described in Materials and Methods section. Each bar represents the average of three or four experiments, with error bars representing the standard errors of the means.
Viable cells (log10 CFU/mL) and total cell biomass were measured after 24 h of growth at 37°C in intact biofilms or in intact biofilms supplemented with antibiotics 50 times their MICs as described in Materials and Methods section. The results are the means ± standard errors of the means from three independent experiments with viable counts done in triplicate.
Significantly different from the corresponding value for the control (p < 0.05); all other values are not significant.
Total bacterial biomass of E. coli and S. aureus biofilms was measured as described in Materials and Methods section. When E. coli or S. aureus biofilms were formed in the presence of DNase I at 5.0 μg/mL, total bacterial biomass of biofilm was reduced by 57–68% (Table 2).
Planktonic antimicrobial susceptibility testing
We studied the in vitro susceptibilities of bacterial strains in planktonic suspension to antimicrobial agents, including representatives of various classes of antibiotics with different mechanisms of action. For instance, ampicillin inhibits bacterial cell wall synthesis and levofloxacin blocks DNA replication. The MIC of ampicillin for E. coli was 16.0 μg/mL and for S. aureus 0.025 μg/mL. The MIC of levofloxacin for E. coli was 0.035 μg/mL and for S. aureus 0.3 μg/mL.
We have also studied the persistence of in vitro susceptibilities of bacteria to antibiotics after 24 h of cultivation in nutrient media supplemented or not supplemented with DNase I 5.0 μg/mL (Table 3). Antibiotic susceptibility patterns of E. coli and S. aureus isolated from biofilms formed in the presence of DNase I were the same as susceptibility of bacteria isolated from intact biofilms.
The results are representative of those from three to four separate experiments.
Effects of antibiotics on intact 24-h-old biofilms and 24-h-old biofilms formed in the presence of DNase I 5.0 μg/mL
Antibiotics at concentration 50 times their MICs were added to intact 24-h-old biofilms or 24-h-old biofilms formed in the presence of DNase I and the mixtures were incubated for 24 h at 37°C.
Ampicillin decreased the biomasses of intact biofilms of both gram-negative and gram-positive bacteria by 11–15%. At the same time, ampicillin added to biofilms formed in the presence of DNase I 5.0 μg/mL reduced the biomass of E. coli biofilms by 18% and biomass of S. aureus biofilms by 19% (Table 2).
Under the same conditions, levofloxacin caused 32% decrease in the biomasses of intact biofilms of gram-negative bacteria. Gram-positive bacteria were less susceptible to levofloxacin, which reduced the biomass of intact S. aureus biofilm by 11%. When bacteria studied were grown in the presence of DNase I 5.0 μg/mL, levofloxacin inhibited the biofilm biomasses of E. coli by 44% and S. aureus biofilms by 57% (Table 2).
Bacteria from intact biofilms and biofilms formed in the presence of DNase I 5.0 μg/mL had quantitatively different levels of viability in the presence of antibiotics (Table 2). Ampicillin did not statistically significantly reduce the numbers of CFU of intact biofilms formed by gram-positive and gram-negative bacteria, although ampicillin caused an approximate four-fold decrease in viable bacterial counts of the E. coli and S. aureus biofilms formed in the presence of DNase I 5.0 μg/mL. Levofloxacin added to intact biofilms caused 14- to 20-fold decreases in viable bacterial counts of the gram-positive and gram-negative bacteria, although levofloxacin added to biofilms formed in the presence of DNase 5.0 μg/mL reduced the numbers of CFU by up to 80-fold.
The effects of antibiotics at 50 times their MICs on total bacterial biomasses of 24-h-old E. coli and S. aureus biofilms were studied (Table 2). Ampicillin reduced total cell biomasses of intact 24-h-old biofilms formed by gram-positive and gram-negative bacteria by 16–24%. When ampicillin was used against biofilms formed in the presence of DNase I 5.0 μg/mL, total cell biomasses in microbial communities formed by E. coli and S. aureus were reduced by 66–67%.
Levofloxacin at concentration 50 times the MIC caused 40% and 58% decrease of total cell biomasses of intact S. aureus and E. coli biofilms. Under the same conditions, when levofloxacin was added to biofilms formed in the presence of DNase I at 5.0 μg/mL, it caused reductions only in total cell biomasses of gram-negative bacteria, reducing total cell biomass by 74%.
When added to the biofilm formed in the presence of DNase I 5.0 μg/mL, antibiotics slightly reduced the total biofilm biomasses and total cell biomasses and significantly decreased the numbers of CFU compared with that when added to intact biofilms.
Discussion
In this study, we investigated the effects of extracellular DNA destruction by DNase I on characteristics of biofilm formation by different unrelated gram-positive and gram-negative bacteria. It is known that bovine pancreatic DNase I with molecular weight of 33 kDa does not penetrate bacteria and is able to cleave only extracellular DNA when is added to bacterial suspension (Liao and Salnikow, 1973). Moreover, DNase I has been shown to retain enzymatic activity for more than 24 h after inoculation to the nutrient media. Our results clearly indicate that DNase I at 5.0 μg/mL effectively cleaves cell-free DNA in biofilm matrix. We have observed that DNase I added to developing biofilms at time zero does not lead to total destruction of extracellular DNA, which can be extracted from the biofilm matrix after biofilm formation. The results suggest that biofilms formed in the presence of DNase I contain extracellular DNA fragments of about 30 kb, which showed the same electrophoretical mobility as DNA fragments isolated from the untreated biofilms. We have also detected total degradation of extracellular DNA by DNase I in bacteria-free extracellular matrix and retaining of DNase I enzymatic activity for more than 24 h in nutrient solution. We consider that these results support the idea that cell-free DNA is constantly released into matrix (Allesen-Holm et al., 2006). We have also detected alteration of biofilm formation in the presence of DNase I. Two different methods have been used to estimate biofilm biomass and total bacterial biomass in our experiments. Biofilm biomass is expressed as amount of crystal violet taken up by the biofilm sample and quantified and expressed as OD absorbance units. Total bacterial biomass was estimated by determining the OD at 570 nm of bacterial cells isolated from biofilms (cells were not stained for the determination of total bacterial biomass). Thus OD values at 570 nm of biofilm biomass should not be compared directly with OD at 570 nm of total bacterial biomass.
The results indicate that DNase I at concentrations varying from 0.1 to 100 μg/mL added to the forming biofilms at time zero results in 9–69% reductions in the community biomasses. We have demonstrated that the addition of DNase I to forming biofilms altered the total bacterial biomasses of biofilms. The results of these experiments are compared using microscopy and with the number of CFU. Destruction of extracellular DNA by DNase I at a concentration of 5.0 μg/mL added to developing biofilms at time zero reduced total bacterial biomasses of E. coli and S. aureus biofilms by 47–54%, and numbers of CFU by twofold (Table 2). These data were confirmed by light microscopy, which revealed changes in morphology of bacterial communities, such as reduced number and sizes of microcolonies and increased area of cell-free zones. Our results thus suggest that extracellular DNA plays an important role in biofilm formation and microbial communities formed in the presence of DNase contain reduced number of viable cells. We have demonstrated that reduction of total bacterial biomasses of biofilms formed by both gram-positive and gram-negative bacteria for the most part was accompanied by a reduction of the CFU number.
The addition of antibiotics at 50 times their MICs to biofilms formed in the presence of DNase I at 5.0 μg/mL decreased the biomasses by 1.25-fold (statistically not relevant) to 5.2-fold, total bacterial biomasses by 1.2–1.4-fold (statistically not relevant), and the numbers of CFU by 1.8–5.3-fold more compared with reductions caused by addition of antibiotics at 50 times their MICs to intact 24-h-old biofilms.
Thus, bacteria in the biofilms formed in the presence of extracellular DNA destruction by DNase I were less tolerant to antibiotics, resulting in moderate reduction of E. coli biofilm biomass and total bacterial biomass, significant reduction of S. aureus biofilm biomass, and moderate reduction of total bacterial biomass. At the same time, we have detected a statistically relevant reduction of the numbers of CFU of both gram-positive and gram-negative bacteria.
The fact that sensitivity to antibiotics of bacteria isolated from the untreated biofilms and biofilms formed in the presence of DNase I was not changed (Table 3) allows us to assume that extracellular DNA plays an important role in the development of biofilm tolerance (Tetz, 2005).
Numerous studies have demonstrated that bacteria in biofilms are embedded within an extracellular matrix that protects biofilm bacteria against influence of environmental factors (Branda et al., 2005; Steinberger and Holden, 2005; Böckelmann et al., 2006; Flemming et al., 2007). These structures reduce the penetration of different molecules including antibiotics into biofilms, making biofilm bacteria more tolerant to antimicrobial agents compared with their planktonic counterparts (Stewart and Costerton, 2001; Larsen, 2002; Stewart, 2002; Fux et al., 2005; Tetz et al., 2004). Previously, antibiotics at 50 times their MICs have been demonstrated to have slight effect on characteristics of formed biofilms (Garcia-Castillo et al., 2007).
Reduction of the biofilm biomass affecting extracellular DNA by DNase I supports an idea that cell-free DNA as a part of EPS plays an important role in the defense of biofilms from environmental factors and antibiotics in particular (Whitchurch et al., 2002; Sponza, 2003; Chambless et al., 2006; Hall-Stoodley et al., 2008; Mulcahy et al., 2008).
We revealed that extracellular matrix of bacterial biofilms formed in the presence of DNase I retains exonuclease activity after 2, 4, 8, 12, and 24 h of biofilm cultivation and contains detectable DNA fragments, which indicates a constant release of cell-free DNA in biofilm matrix. Moreover, the destruction of extracellular DNA during biofilm formation decreases defense forces of biofilms toward environmental factors including different unrelated antibiotics with different mechanisms of action.
As soon as extracellular DNA is found to be a component of many bacterial biofilms, it can serve as a universal target perhaps allowing for broad and stereotypic changes of biofilms properties including interactions with antibacterial agents.
Footnotes
Acknowledgment
The authors thank Michael Cynamon for helpful comments during preparation of the manuscript and for editing the manuscript.
Disclosure Statement
No competing financial interests exist.