
Abstract
SUN domain-containing proteins belong to a novel protein family. To date, several members—SUN1, SUN2, SUN3, and SPAG4—have been identified as nuclear envelope (NE) proteins. In this study, we sought to characterize and define the potential function of SPAG4L, a newly identified SUN protein. Using bioinformatic analysis, we found that SPAG4L contained a conserved SUN domain in the C-terminal. Subcellular localization analysis indicated that the expression of green fluorescent protein-labeled full-length SPAG4L was localized to the NE and the endoplasmic reticulum (ER). Deletion analysis revealed that the transmembrane region and the coiled-coil domain, but not the SUN domain, were required for localization of SPAG4L to the NE and ER. Subsequently, we confirmed that the human testes expressed endogenous SPAG4L as a 43-kDa protein. Further studies revealed that mouse Spag4L colocalized with the NE marker Lamin B1 and the ER marker PDI in isolated mouse spermatocytes. In addition, the expression of Spag4L was observed in meiosis I and II stages, suggesting that Spag4L may be involved in NE reconstitution and nuclear migration occurring during the process of spermatocyte division. Together, the findings indicate that SPAG4L, a new NE protein, may play an important role in the meiotic stage of spermatogenesis.
Introduction
In the process of spermatogenesis, spermatocytes undergo division and differentiation. After a series of biochemical and morphological changes, spermatozoa are generated (Jamsai and O'Bryan, 2010). Many genes are involved in each stage of spermatogenesis (Liu et al., 2004; Mu et al., 2006; Yang et al., 2011). Recent reports have indicated that a novel gene family, encoding proteins containing the SUN domain, is associated with spermatogenesis. To date, several human SUN proteins have been identified: UNC-84A (SUN1), UNC-84B (SUN2), SUN3, SPAG4, and SPAG4L. The common features of SUN proteins is that all these proteins contain a conserved C-terminal SUN domain (Hodzic et al., 2004; Hasan et al., 2006; Wang et al., 2006; Gob et al., 2010).
Among these newly identified proteins, SUN1 has been identified as a nuclear envelope (NE) protein that is important for the anchorage of nesprin-2, an NE-associated spectrin-repeat protein (Padmakumar et al., 2005). Further studies demonstrated that SUN1 specifically associates with telomeres between the leptotene and diplotene stages of meiotic prophase I. Disruption of Sun1 in mice prevents telomere attachment to the NE, thereby resulting in infertility (Ding et al., 2007). Recent studies found that there are several transcripts of SUN1 expressed in mouse testis, among which only SUN1η is expressed in the testis and correlates with the appearance of an anterior nonnuclear LINC complex (a complex that links the nucleoskeleton and cytoskeleton) during sperm head formation (Gob et al., 2010). Human SUN2, another human SUN domain-containing protein, is reported as an inner nuclear membrane (INM) protein, which facilitates the tethering of mammalian meiotic telomeres to the NE (Hodzic et al., 2004; Schmitt et al., 2007). Moreover, Zhang et al. (2009) demonstrated that SUN1/2 and Syne/Nesprin-1/2 form complexes that connect the centrosome to the nucleus during neurogenesis and neuronal migration in mice. Sun3 is a short testis-specific member of the SUN domain protein family, whose expression correlates with sperm head formation, defining a novel LINC complex that colocalizes with the manchette (Gob et al., 2010).
SPAG4, the fourth identified human SUN domain-containing protein, was originally isolated from the testis (Tarnasky et al., 1998). Hasan et al. (2006) demonstrated that the expression of green fluorescent protein (GFP)-tagged SPAG4 localizes to the endoplasmic reticulum (ER) and the NE. Shao et al. (1999) showed that Spag4 was detectable in the transient manchette and was associated with the axoneme in elongating spermatids and epididymal sperms in the rat. Recently, Kracklauer et al. (2010) reported that Spag4 was required for the attachment between the centriole and the nucleus during spermatogenesis in Drosophila spp. They showed that in the absence of Spag4, the nuclei and centrioles or basal bodies dissociate from each other after meiosis.
In our previous study, we isolated a SPAG4-like gene, SPAG4L (synonyms: SUN5 and TSARG4; GenBank accession number: AF401350), from a human testis library (Xing et al., 2003). Subsequently, we found a homologous gene, Spag4L (synonym: SRG4; GenBank accession number: AY307077), in a mouse testis library (Xing et al., 2004). The putative amino acid sequence of SPAG4L shares 45% homology with that of SPAG4 (Xing et al., 2003). Early studies showed that Spag4 is mainly presented in round spermatids and spermatozoa (Shao et al., 1999). In contrast, in situ hybridization revealed that Spag4L mRNA is mainly expressed in the spermatocytes and round spermatids of mice (Xing et al., 2004). Guo et al. (2009) showed that Spag4L expression decreased to about 45% from pachytene spermatocytes to elongating spermatids, suggesting that Spag4L may play an important role in meiosis. Recently, Frohnert et al. (2011) reported that there is another isoform named Spag4L-2 also specifically expressed in mouse testis, which together with Spag4L is restricted to the apical NE of round spermatids facing the acrosome. Here, we report the distribution of SPAG4L expression and its potential function during spermatogenesis.
Materials and Methods
Bioinformatic analysis
The putative amino acid sequence of SPAG4L was aligned with those of SUN1, SUN2, SUN3, and SPAG4, using the Jellyfish sequence analysis software (
Protein extraction and western blot analysis
All experiments on human tissues were approved by the Ethics Committee of Third Xiang-Ya Hospital of Central South University. Human testis samples for protein extraction were collected from two prostate cancer patients after obtaining oral consent from them. These patients were scheduled for bilateral castration as part of cancer treatment at the Department of Urology, Third Xiang-Ya Hospital of Central South University. Human peripheral blood was collected from volunteers. Fat, muscle, and liver tissue specimens were obtained from patients with liver cancer surgery. Kidney tissue specimens were from the biopsy surgery.
All tissues and cultured cells were treated with RIPA lysis buffer (150 mM NaCl, 25 mM HEPES, 2 mM NaF, 0.2% SDS, and 1% NP-40, with freshly added protease inhibitor PMSF) for 30 min on ice. The lysates were clarified by centrifugation at 13,000 rpm for 15 min at 4°C. Proteins were separated by 12% SDS-PAGE and transferred onto PVDF membranes (Millipore). Membranes were blocked in blocking buffer for 2 h and incubated overnight at 4°C with SPAG4L antibody (ProteinTech Group, Inc.). The remaining steps of western blotting were performed according to the manufacturer's instructions (Auragene Bioscience).
Plasmids
To determine the sites of SPAG4L localization and its domains, the full-length open-reading frame and different deletion mutants were amplified from a pUCm-T/SPAG4L plasmid by PCR. The primer sequences are shown in Table 1. The products were inserted into a pUCm-T vector, and the insert fragments were assessed using restriction enzymes HindIII and SalI (Fermentas). Subsequently, these fragments were subcloned into pEGFP-N1 (Clontech), a transient expression vector, and the accuracy of the orientation was confirmed by restriction digestion and DNA sequencing. Prokaryotic expression plasmids of pQE30-SPAG4, pQE30-hSPAG4L, and pQE30-mSpag4L were constructed in our previous study.
Underlined sequences sites for restriction endonuclease.
Cell transfection for cellular localization
HeLa cells were maintained in DMEM medium supplemented with 10% fetal bovine serum (Gibco). We placed cover slips at the bottom of each well of six-well plates and cultured 1×105 cells/well for 24 h before transfection. The recombinant plasmids were transfected in triplicate using 4 μg of plasmids and 10 μL of Lipofectamine 2000 (Invitrogen), according to the manufacturer's protocol. Twenty-four hours after transfection, the cover slips were removed, and the cells were fixed with 4% paraformaldehyde for 30 min. Subsequently, the expression of the fusion protein was detected using inverted fluorescence microscope (IX71; Olympus) and confocal microscope (Olympus FluoView FV1000).
Animals
Balb/c mice were obtained from the experimental animal center at our university. They were maintained and housed according to approved guidelines (Guidelines for the Care and Use of Laboratory Animals).
Isolation of germ cells from mouse seminiferous tubules and immunofluorescence analysis
Testis tissues were obtained from adult Balb/c mice, and the fat and connective tissues were removed; seminiferous tubules were then minced in 0.1 M PBS on ice. Germ cells were filtered using a sieve and washed with 0.1 M PBS three times. The cells were then treated with a hypotonic solution for 20 min, fixed with 4% paraformaldehyde for another 20 min, and plated onto slides. Immunofluorescence staining was performed as previously described (Cherry and Hsu, 1984; Scherthan et al., 2000). Goat antibodies against mouse Spag4L (Santa Cruz) and rabbit antibodies against human SPAG4L and mouse Spag4L were used. Other antibodies used were mouse monoclonal antibodies against Lamin B1 (Santa Cruz) and mouse monoclonal antibodies against PDI (Santa Cruz). The cell nuclear DNA was dyed with DAPI. The specimens were then observed under a fluorescence microscope (Nikon Eclipse 80i).
Results
Homology analysis of SPAG4L
Comparison of the sequence of SPAG4L with those of the known SUN domain-containing proteins SUN1, SUN2, SUN3, and SPAG4 revealed that the former also contained a C-terminal SUN domain, suggesting that SPAG4L was a SUN protein (Fig. 1A). Phylogenetic analysis of the SUN-domain amino acid sequences further indicated that the genetic distance between SUN1 and SUN2 was 0.01190. Thus, SUN1 and SUN2 probably originated from a close common ancestor. However, the genetic distance between SPAG4 and SPAG4L was 0.05319, suggesting that SPAG4 and SPAG4L probably originated from another ancestor (Fig. 1B). Overall, these studies suggested that SPAG4L was more closely related to SPAG4 than to SUN1, SUN2, and SUN3.

Homology analysis of SPAG4L with known SUN-domain proteins.
Transfected SPAG4L expression localizes to the NE and the ER in HeLa cells
To determine the site of expression of human SPAG4L, we generated the pEGFP-N1-SPAG4L plasmid encoding the SPAG4L-GFP fusion protein. After transfection of this plasmid into HeLa cells, we found that GFP expression was predominantly localized to the NE and ER (Fig. 2A). However, the GFP was detected in both the cytosol and nuclei in the cells transfected with the negative control plasmid pEGFP-N1, which encoded only GFP and not SPAG4L (Fig. 2A). Western blot analysis revealed that a 70-kDa band (GFP-tagged SPAG4L) was detected in cells transfected with pEGFP-N1-SPAG4L using anti-SPAG4L or anti-GFP antibody, whereas a 27-kDa GFP band was only detected in cells transfected with pEGFP-N1 by anti-GFP antibody (Fig. 2B). Subsequently, ER-Tracker Red was used to recognize ER in living cells. We found that the GFP-tagged SPAG4L fusion proteins were colocalized with red fluorescence-labeled ER in HeLa cells transfected with pEGFP-N1-SPAG4L (Fig. 2C).
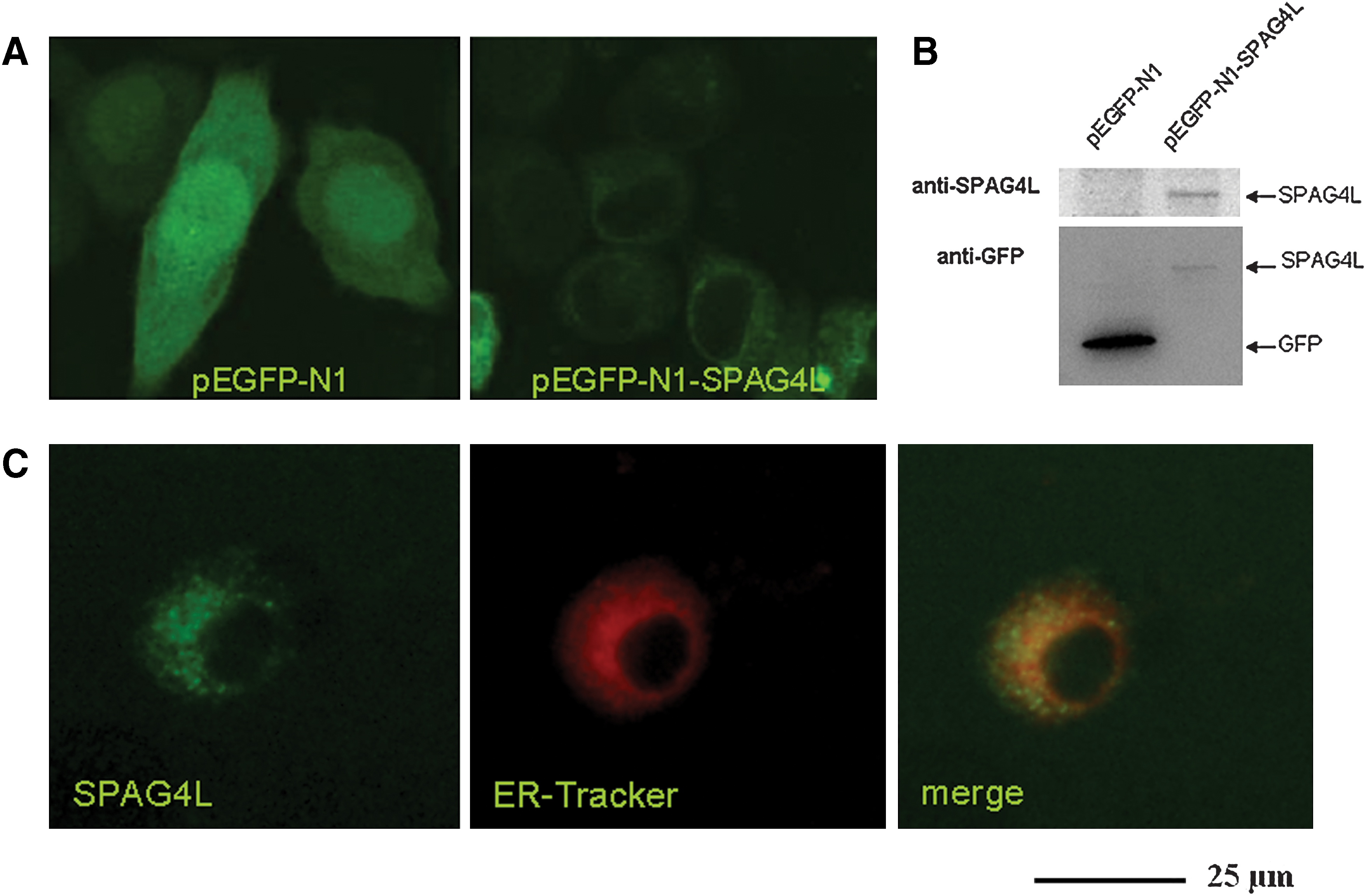
Subcellular localization of SPAG4L protein.
Spag4L was localized to the ER and NE of mouse spermatocytes
Because it was difficult to obtain human testis from young individuals, we used mouse models to analyze the expression profile of Spag4L during spermatogenesis. We isolated germ cells from mice and performed immunofluorescence analysis to assess the localization and expression pattern during this process. In particular, NE and ER markers (Lamin B1 and PDI, respectively) were used. The results of the analysis showed that Spag4L was colocalized with the nuclear membrane protein marker Lamin B1 (Fig. 3A) and the ER marker PDI (Fig. 3B) in spermatocytes. Together, these results provide evidence that Spag4L is expressed in the ER and associated with NE.

Spag4L colocalized with the ER marker PDI and the nuclear membrane protein Lamin B1 in mouse spermatocytes.
Transmembrane region and coiled-coil domain are required for proper localization
To identify the region that is critical for localization of SPAG4L to the nuclear envelop, a series of GFP-SPAG4L deletion mutants were generated and introduced into HeLa cells (Fig. 4A). The results showed that two deletion mutants (aa 1–200 and aa 52–230), which contained the transmembrane region and the coiled-coil domain, localized to the NE and ER. However, the C-terminal region of SPAG4L (aa 221–379), which mainly included the SUN domain, was found distributed throughout the cells. Deletion of the N-terminal region of SPAG4L (aa 1–220), which included the transmembrane region and the coiled-coil domain, resulted in complete absence of the NE and ER localization of SPAG4L. Together, these data indicated that the transmembrane region and the coiled-coil domain, but not the SUN domain, were required for the localization of SPAG4L to the NE (Fig. 4B).

Localization analysis of deletion mutants of SPAG4L.
Detection of endogenous SPAG4L protein by western blotting
We first determined the specificity of anti-SPAG4L antibody. Western blot of prokaryotic pQE30 expression of hSPAG4, hSPAG4L, and mSpag4L with anti-His-tag showed 36-, 31-, and 23-kDa products, respectively, but using anti-human SPAG4L antibody showed specificity to human SPAG4L, cross-reaction to mouse Spag4L, and no reaction to human SPAG4 (Fig. 5A). Western blot of different tissues showed that SPAG4L was expressed specifically in testis (a single 43-kDa band), but not in other tissues (Fig. 5B). Thus, these results provide evidence that the human testis contains endogenous SPAG4L.

Specificity of anti-SPAG4L antibody and detection of endogenous SPAG4L protein in human testis.
Detection of endogenous Spag4L in different stages of meiosis
Our early studies showed that mouse Spag4L was abundantly expressed starting from the age of 3 weeks and mainly expressed in spermatocytes and round spermatids (Xing et al., 2004). To determine the potential involvement of Spag4L during spermatogenesis, we used immunostaining to examine the intracellular distribution of endogenous Spag4L at different stages of meiosis. For immunostaining, we used the anti-Spag4L antibody (Santa Cruz), an affinity-purified goat polyclonal antibody against the N-terminus of mouse Spag4L. We found that at the onset of the first round of division (prophase I), Spag4L proteins were distributed along the rim of the nuclei in leptotene spermatocytes and pachytene spermatocytes. By metaphase I, as the NE was torn apart, NE transmembrane proteins detached from chromatin and redistributed into the ER (Kutay and Hetzer, 2008). We found that Spag4L protein formed a loop around chromosomes. During the period between anaphase I and telophase I, the chromosomes migrated to the opposite poles. Spag4L proteins separated the condensed chromatin from the sides and reconstituted the NE. At the end of meiosis, the NE was reconstituted in the daughter cells, and the secondary spermatocytes were generated (Fig. 6).
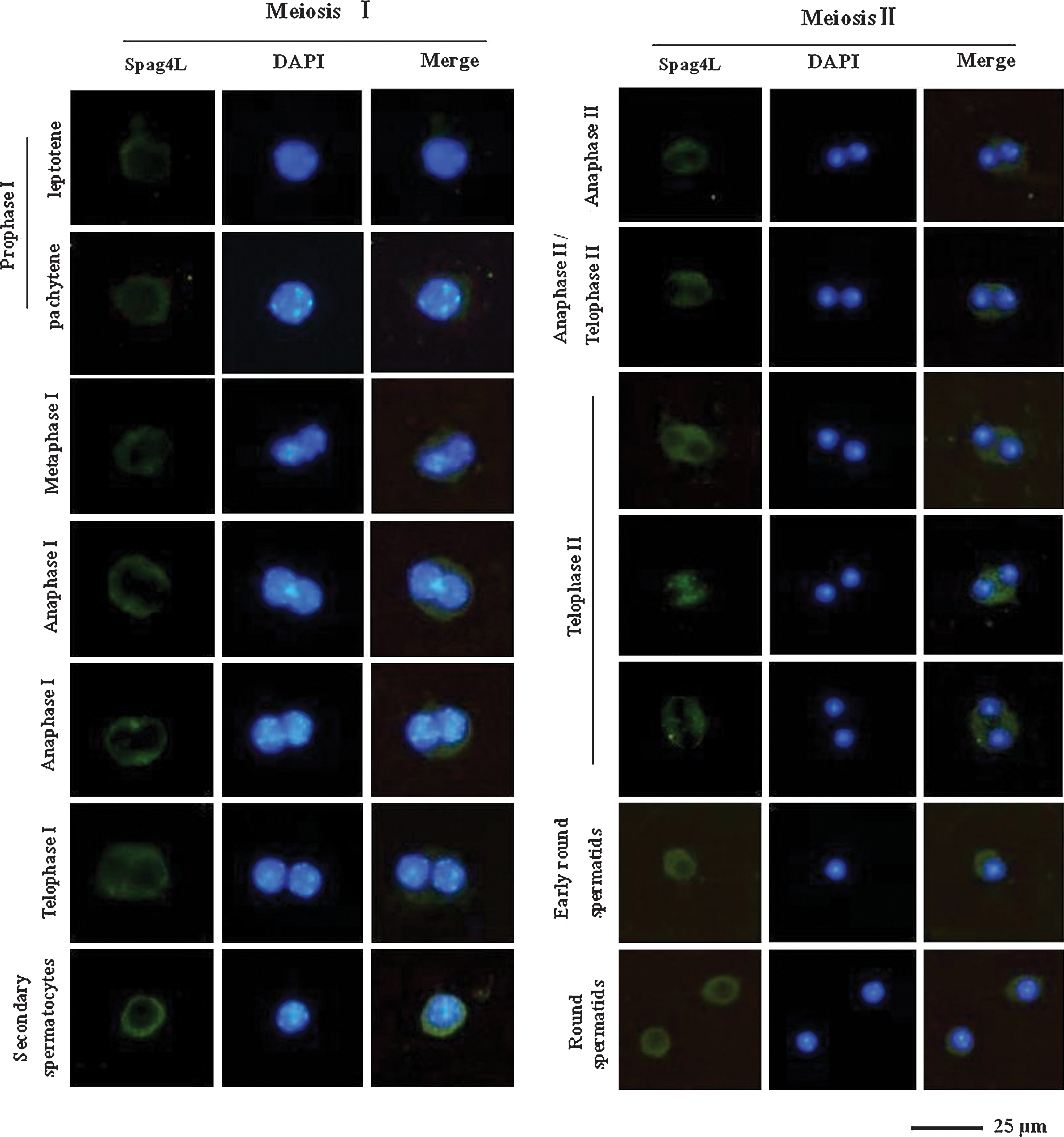
The expression of Spag4L in mouse spermatocyte division. Spermatocytes and round spermatids were labeled with goat antibodies against Spag4L, and donkey anti-goat IgG (H+L) conjugated with FITC (Southern Biotech) was used for immunofluorescence staining. DNA was counterstained with DAPI. Color images available online at
In phase II of meiosis, the secondary spermatocytes undergo a second round of division. Between anaphase II and telophase II, when the nucleus divided into two parts and the parts migrated to the opposite poles, Spag4L proteins separated and surrounded the condensed chromatin from both the sides. Subsequently, Spag4L proteins appeared in a large amount between two nuclei. During telophase II, while the condensed chromatin migrated toward the two poles, Spag4L proteins likely participated in tethering the two nuclei to both sides. When the nuclei reached the two poles, Spag4L was mainly distributed between the nuclei and possibly contributed to the NE reconstitution. In fact, Spag4L was observed to encircle the nuclei in the stage of round spermatids (Fig. 6). Together, these findings provide considerable data of the temporal and spatial expression patterns and the expression levels of Spag4L at different time points during spermatocyte division; these data suggest that Spag4L was probably involved in NE reconstitution and nuclear migration in the meiotic stage of spermatogenesis.
Discussion
SUN proteins are a family of NE proteins involved in spermatogenesis (Shao et al., 1999; Ding et al., 2007; Schmitt et al., 2007). Here, we characterized SPAG4L, a novel SUN protein, which is expressed in the human testis. Deletion analysis revealed that the transmembrane region and coiled-coil domain, but not the SUN domain, are required for the localization of SPAG4L in the NE and ER. Further, the findings of human studies were consistent with those of our mouse studies. Finally, we observed the expression of Spag4L in spermatocyte division process and the result suggested that Spag4L may be involved in the process of nuclear migration and NE reconstitution during spermatogenesis.
The NE separates the cytoplasm from the nucleus and plays an important role in nuclear positioning during cell division (Kutay and Hetzer, 2008, Graumann et al. 2010). Previous studies have shown that SUN1 and SUN2 were ubiquitously expressed except SUN1η (Gob et al., 2010). SUN1 and SUN2 interacts with KASH domain proteins and formed the LINC complexes that connect the centrosome to the nucleus (Hodzic et al., 2004; Padmakumar et al., 2005; Hasan et al., 2006; Wang et al., 2006; Zhang et al., 2009). In contrast, SPAG4 is expressed in a limited number of normal tissues, notably the pancreatic and testicular tissues, and the expression of SPAG4 was localized to the ER and the NE (Shao et al., 1999; Hasan et al., 2006; Kracklauer et al., 2010). In this study, we demonstrated that the SPAG4L expression is localized to the NE and ER. Similar to human studies, mouse studies showed that the homologous protein, Spag4L, is localized with either the NE (Lamin B1) or ER (PDI) markers in mouse spermatocytes. According to the diffusion-retention model, proteins must interact with the nuclear lamina or chromatin to be retained at the inner NE (Worman and Courvalin, 2000). We speculate that SPAG4L may interact with Lamin B1 or other lamins to anchor onto the NE. In addition, the transmembrane domain and the coiled-coil domain, but not the SUN domain, were found to be essential for the proper localization of SPAG4L. Recently, Kracklauer et al. (2010) demonstrated that Spag4 mutant male Drosophila were sterile. Male fertility was restored to Spag4 mutants by introducing a single copy of a 4.9-kb genomic spag4 transgene. A transgene generating Spag4 protein lacking the SUN domain did not restore fertility to spag4-null males, indicating that Spag4 requires this domain for its role in spermatogenesis. Although the SUN domain did not contribute to SPAG4L's localization, SPAG4L may play an important role in male fertility.
Recently, Frohnert et al. (2011) reported that there is another isoform named Spag4L-2 also specifically expressed in mouse testis. We do not exclude the existence of SPAG4L isoforms in human testis.
In our early studies, we found that Spag4L was abundantly expressed from the age of 3 weeks, mainly in spermatocytes and round spermatids; this suggests that Spag4L may play an important role in the meiosis stage of spermatogenesis (Xing et al., 2004). In this study, we observed the dynamic pattern of Spag4L in spermatocyte divisions and found that Spag4L was involved in the spermatocyte division process, especially in meiosis II stage. We speculated that Spag4L was probably involved in NE reconstitution and nuclear migration in the meiotic stage. Zhang et al. (2009) demonstrated that SUN1/2 and Syne/Nesprin-1/2 complexes connected the centrosome to the nucleus during neurogenesis and neuronal migration in mice. However, the role played by Spag4L in the process of nuclear migration remains to be elucidated. Our findings provide a strong basis for further studies in this regard.
In summary, we have identified SPAG4L, a novel SUN protein localized to the NE and ER and probably involved in NE reconstitution and nuclear migration occurring in meiotic stages of spermatogenesis. In our future studies, we will attempt to determine whether Spag4L interacts with nesprin proteins or nuclear lamina proteins, such as Lamin B1, to regulate nuclear migration.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 30600681) and China Postdoctoral Science Foundation (No. 20090461024).
Disclosure Statement
No competing financial interests exist.