
Abstract
Altered folate levels may play an important role in colon carcinogenesis. The aim of this study was to investigate the association of polymorphisms in key folate-metabolizing genes with susceptibility to sporadic colon cancer. Six common polymorphisms (two in MTHFR and one each in MTR, MTRR, RFC1, and DHFR genes) were genotyped in 300 healthy subjects and 300 colon cancer patients from Croatia. Obtained results indicate possible protective role of MTRR 66 AA in sporadic colon cancer (OR = 0.655; 95% CI = 0.441–0.973; p = 0.04). Maximum-likelihood analysis of haplotypes revealed a linkage disequilibrium (LD) between the two investigated polymorphisms of the MTHFR gene (C677T and A1298C), both in the control and patient groups (p < 0.01 for both). LD was also detected between MTRR A66G and MTHFR A1298C polymorphisms but only in a group of patients (p < 0.01). A haplotype of A66G and A1298C polymorphisms, A/A, proved to be protective (OR = 0.775; 95% CI = 0.603–0.996; p = 0.04), whereas haplotype A/G was a risk factor for colon cancer (OR = 1.270; 95% CI = 1.007–1.602; p = 0.04). Contrary to some previous studies, single-locus analyses identified no polymorphisms associated with risk for colon cancer, but demonstrated a possible protective effect of MTRR 66 AA genotype. The detected significant LD between two loci (MTHFR A1298C and MTRR A66G) located on different chromosomes indicates a strong selective force as a mechanism for the maintenance of their linkage. Specific combinations of alleles of these two polymorphisms showed a protective but also a risk effect on colon cancer susceptibility.
Introduction
A key enzyme in the folate pathway is MTHFR, which catalyzes the reduction of 5,10-methylenetetrahydrofolate (5,10-methyleneTHF) to 5-methyltetrahydrofolate (5-methylTHF). 5-MethylTHF is the major circulatory form of folate in the body and is used as a methyl donor for homocysteine remethylation into methionine, which then converts to S-adenosylmethionine (SAM), the universal methyl donor in the cell (Slattery et al., 1999).
The MTHFR gene is positioned at 1p36.3 and its most common polymorphism is C677T (1p36.22, rs1801133), which leads to substitution of alanine by valine (A222V). This polymorphism causes a thermolabile enzyme with lower activity (Frosst et al., 1995).
Another common MTHFR polymorphism is A1298C (1p36.22, rs1801131), which leads to the substitution of glutamate by alanine at position 429. It was associated with decreased enzyme activity but to a lesser extent than the C677T polymorphism (Chen et al., 2002).
The methylation of homocysteine to methionine is catalyzed by MTR, which uses 5-methylTHF as a methyl donor and cobalamin (vitamin B12) as a cofactor. During the reaction, transfer of the methyl group from methylcob(III)alamin results in the formation of the highly reactive cob(I)alamin, which may become oxidized to cob(II)alamin, occasionally resulting in MTR inactivation (Leclerc et al., 1998). MTRR catalyzes reductive methylation of cob(II)alamin by using SAM as a methyl donor, which reactivates MTR (Leclerc et al., 1998). Thus, MTRR may act as a key regulator of homocysteine conversion to methionine. The MTR gene is located on 1q43 and contains an A2756G polymorphism (1q43a, rs1805087). It causes substitution from aspartic acid to glycine at codon 919 (D919G) and may modify enzyme activity (Harmon et al., 1999; Matsuo et al., 2002). The MTRR gene is located at 5p15.2–15.3 (Leclerc et al., 1998) and A66G polymorphism (5p15.31a, rs1801394) results in the substitution of isoleucine with methionine at codon 22 (Wilson et al., 1999). MTRR 66 G allele decreases enzyme activity compared with the A allele (Matsuo et al., 2002).
RFC1 is responsible for active transport of 5-MeTHF from plasma to cytosol (Ulrich et al., 2005b). The RFC1 gene is located at 21q22.2–22.3 and a common polymorphism is G80A (rs1051266), which is positioned in exon 2 (Winkelmayer et al., 2003) and causes Arg27His substitution. The functional consequence of this polymorphism is not fully understood.
DHFR catalyzes reduction of ingested folic acid into fully active tetrahydrofolate, which then can be utilized for cellular reactions. The DHFR gene (5q11.2–13.2) may contain a polymorphic 19-bp deletion within intron 1, which can remove a potential Sp1 transcription binding site, consequently leading to lowered levels of DHFR and reduced folates in the cell (Johnson et al., 2004).
The aim of this study was to investigate the possible association of the polymorphisms in MTHFR, MTR, MTRR, RFC1, and DHFR genes with the susceptibility to sporadic colon cancer.
Materials and Methods
Subjects
The study included 300 unrelated healthy volunteers and a group of 300 patients with sporadic colon cancer. Subjects in the investigated groups were of comparable age and gender. All specimens were obtained from the Croatian Tumor and DNA Bank for Basic Research, Ruđer Bošković Institute, Zagreb, Croatia (Spaventi et al., 1994). The control group consisted of 152 men (average age of 60 years, ranging from 40 to 87 years) and 148 women (average age of 63 years, ranging from 40 to 89 years). The cases group consisted of 162 men (average age of 61 years, ranging from 35 to 86 years) and 138 women (average age of 64 years, ranging from 39 to 90 years) with negative family history of hereditary cancer. Genomic DNA was extracted from whole-blood samples using standard phenol/chloroform extraction. This study was approved by the Ethics Committees of Clinical Hospital Dubrava and Clinical Hospital Center Zagreb.
Genotyping
The genotyping of MTHFR C677T (rs1801133), MTHFR A1298C (rs1801131), MTR A2756G (rs1805087), and MTRR A66G (rs1801394) polymorphisms was performed on a real-time PCR ABI PRISM 7000 Sequence Detection System (Applied Biosystems) using predesigned TaqMan SNP genotyping assays, provided by Applied Biosystems.
The genotyping of RFC1 G80A was performed by PCR-RFLP method as previously described (Winkelmayer et al., 2003). Briefly, the following set of primers for PCR was used: forward primer, 5′-AGTGTCACCTTCGTCCC-3′; reverse primer, 5′-TCCCGCGTGAAGTTCTTG-3′. The product was subjected to RFLP using CfoI (New England BioLabs), and products were observed on 10% polyacrylamide gel as 125, 68, and 37 bp for wild type and 162 and 68 bp for variant type.
The genotyping of a 19-bp deletion in the DHFR gene was performed by PCR (Johnson et al., 2004). Briefly, the first forward primer was 5′-CCACGGTCGGGGTACCTGGG-3′, the second forward primer was 5′-ACGGTCGGGGTGGCCGACTC-3′, and the reverse primer was 5′-AAAAGGGGAATCCAGTCGG-3′. The product with a 19-bp deletion was observed as a 92 bp band (second forward and a reverse primer) and an amplicon with nondeletion allele as a 113 bp band (first forward and a reverse primer) on the 10% polyacrylamide gel.
Statistical analysis
The differences in genotype and allele frequencies between case and control groups were tested by two-sided Fisher's exact test. The OR and 95% CI were calculated for each SNP according to the method of Woolf, using GraphPad InStat software (version 3.01). The level of statistical significance was set at 0.05. Haplotype frequencies were estimated from genotypic data by the maximum-likelihood method based on EM algorithm using the Arlequin program (Schneider et al., 2000). The same program was used to determine the HW equilibrium and linkage disequilibrium (LD) between all pairs of loci at all population levels. The frequencies of combinations of alleles (haplotypes) of MTRR A66G and MTHFR A1298C loci were calculated according to Hill (1974). The statistical power of our sample was evaluated using free Power and Sample Program (
Results
Genotype and allele frequencies of the investigated polymorphisms in controls (n = 300) and colon cancer patients (n = 300), as well as corresponding ORs and 95% CIs, are summarized in Table 1. Genotype frequencies of all polymorphisms in both groups were in Hardy–Weinberg equilibrium. No significant difference between male and female populations was observed. The calculation of ORs with 95% CIs according to the method of Woolf, wherein all three genotypes were investigated without considering any genotype to be a referent, demonstrated possible protective role of the MTRR 66 AA genotype (OR = 0.655; 95% CI = 0.441–0.973; p = 0.04) in colon cancer ethiopathogenesis. Genotypes MTRR AG (OR = 1.238; 95% CI = 0.898–1.706; p = 0.19) and MTRR GG (OR = 1.085; 95% CI = 0.761–1.548; p = 0.65) showed no significant effect. Single-locus analyses of all other SNPs yielded null results for association with colon cancer. The statistical power of our sample to detect differences associated with an OR of 2.0 at the level of significance of 0.05 was 0.981, 0.982, 0.958, 0.982, 0.986, and 0.986 for MTHFR C677T, A1298C, MTR A2756G, MTRR A66G, RFC1 G80A, and DHFR 19-bp del, respectively. At an OR of 1.5, the statistical power was 0.627, 0.633, 0.535, 0.660, 0.668, and 0.663.
ORs with 95% CIs were calculated according to the method of Woolf, using GraphPad Instat software. ORs for every genotype were calculated based on the comparison of the respective genotype with the combination of the other genotypes. This approach was used because of a high frequency of MTRR 66 G variant allele disabling the usage of MTRR 66 AA as a referent.
Maximum likelihood analysis of haplotype distributions between all pairs of loci demonstrated the presence of highly significant LD between MTHFR C677T and MTHFR A1298C polymorphisms in both controls and cases (D' = 0.94 and 1.00, respectively; p < 0.01 for both). Analysis of LD revealed the presence of a weak (D' = 0.24), but significant (p < 0.01), linkage between MTHFR A1298C and MTRR A66G polymorphisms in the patient group. We observed that MTHFR 1298 A/MTRRR 66 G haplotype acted as a risk factor (OR = 1.270; 95% CI = 1.007–1.602; p = 0.04) for colon cancer, whereas the MTHFR 1298 A/MTRR 66 A haplotype appeared to be protective (OR = 0.775; 95% CI = 0.603–0.996; p = 0.04). The other two haplotypes, MTHFR 1298 C/MTRR 66 A and MTHFR 1298 C/MTRR 66 G, showed no significant effect (OR = 1.064; 0.790–1.433; p = 0.68 and OR = 0.906; 95% CI = 0.655–1.252; p = 0.55, respectively) (Table 2).
As we used the method of Woolf, using GraphPad Instat software, for calculation of ORs for genotypes, ORs for every haplotype between MTHFR A1298C and MTRR A66G were also calculated based on the comparison of the respective haplotype with the combination of the other haplotypes.
The presence of association between colon cancer susceptibility and MTHFR A1298G and MTRR A66G haplotypes prompted us to analyze the association of the combinations of genotypes at these two loci with colon cancer. The observed values of combinations of MTHFR A1298G and MTRR A66G genotypes are presented in Figure 1. Genotype combinations that harbored the MTHFR 1298 A/MTRR 66 G risk haplotype (AA/GG and AC/GG) were slightly more frequent in cases than in controls. Genotype combinations that harbored the protective MTHFR 1298 A/MTRR 66 A haplotype (AA/AA and AC/AA) were slightly more frequent in controls than in cases, but no genotype combination proved to be significantly associated with colon cancer susceptibility.
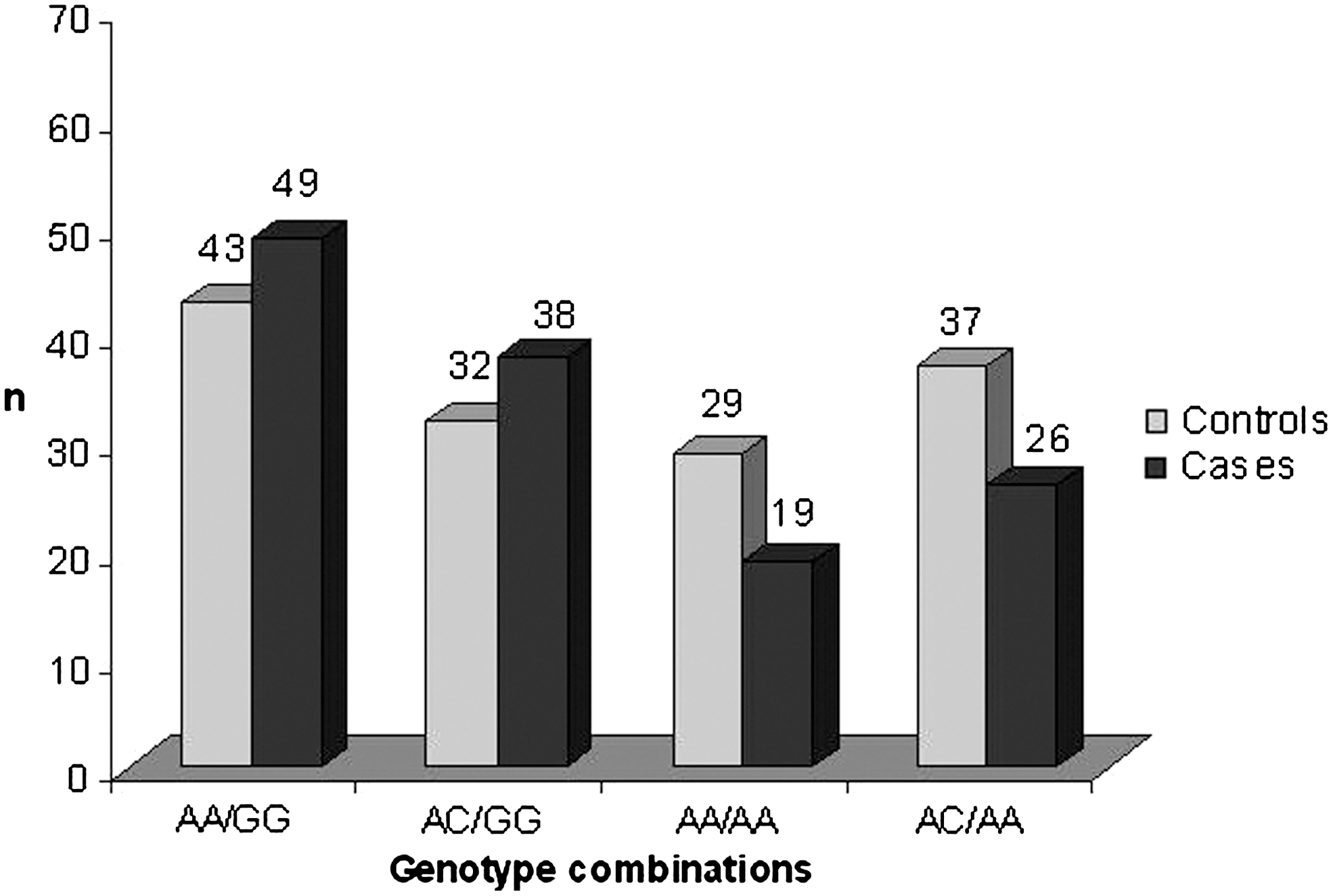
Combinations of MTHFR A1298C/MTRR A66G genotypes. The number of individuals carrying combinations of MTHFR A1298C and MTRR A66G polymorphisms is shown. Combinations of genotypes AA/GG and AC/AG, which harbor risk haplotype A/G but not protective A/A haplotype, are more common in cases than in controls. Combinations of genotypes AA/AA and AC/AA, which harbor protective A/A haplotype but not risk A/G haplotype, are more common in controls than in cases. Because of a relatively small number of individuals carrying these genotype combinations, no genotype combination proved to be significantly associated with higher or lower risk of colon cancer.
Discussion
As altered folate status may influence colon carcinogenesis, we hypothesized that polymorphisms in the key folate-metabolizing genes, which may influence the enzyme's activity, might be associated with susceptibility to colon cancer.
MTHFR C677T has been greatly investigated for susceptibility to colon cancer but with contradictory results. A study by Ma et al. (1997) observed that men with adequate folate levels and 677 TT genotype had decreased risk of colon cancer. Slattery et al. (1999) also observed that high levels of folate intake and a 677 TT genotype were associated with reduced risk of colon cancer. A meta-analysis of 25 populations by Hubner and Houlston (2007) indicated that individuals carrying the 677 TT genotype were at moderately reduced risk of colorectal cancer risk. Also, a meta-analysis by Huang et al. indicated that, overall, the 677T allele showed a small but significant protective effect against colorectal cancer risk (Huang et al., 2007). On the other hand, Shannon et al. (2002) showed that 677 TT was associated with an increased risk of colorectal cancer in older populations, and a study by Haghighi et al. (2009) observed a decreased risk of colon cancer in individuals with 677 CC genotype and this association was stronger at higher levels of folate intake. Also, there are studies that found no association of 677 TT with colon cancer risk (Keku et al., 2002; Sachse et al., 2002; Komlósi et al., 2010), which is in accordance with our results, although we were not able to evaluate the impact of folate intake.
Several studies have investigated the role of MTHFR A1298C in colorectal cancer. A study by Chen et al. (2002) reported slightly reduced risk in subjects with a CC compared with those with an AA genotype, but results did not reach statistical significance. A study by Curtin et al. (2004) showed that 1298 CC genotype as well as MTHFR 677 CC/1298 CC genotypes were significantly associated with lower risk in women. Also, it was reported that 1298 CC genotype was inversely associated with colon cancer in whites (Keku et al., 2002). A meta-analysis by Huang et al. (2007) found a significantly decreased risk of colorectal cancer for the 1298C polymorphism. Our study revealed no association of MTHFR A1298C alone or in combination with C677T with the risk of colon cancer.
The association of MTR A2756G with the disease was investigated in few studies (Chen et al., 1998; Ma et al., 1999; Ulrich et al., 2005b; Eussen et al., 2010), which observed no association with colon cancer susceptibility as confirmed by our study. However, a meta-analysis of Yu et al. (2010) showed that 2756 GG genotype was associated with a significantly reduced risk in European populations, whereas a significant association between the MTR 2756 GG genotype and cancer risk was observed in Asian populations.
It was shown that the RFC1 G80A polymorphism was not associated with colon cancer risk (Ulrich et al., 2005b), but the frequency of the RFC1 80 G allele was found to be increased in children with neural tube defects (De Marco et al., 2001). The DHFR 19 bp polymorphism was associated with risk of autism (Adams et al., 2007), but we found no association of these polymorphisms with risk of colon cancer.
The homozygous variant of the MTRR A66G polymorphism (GG) was associated with premature coronary artery disease (Brown et al., 2000) and, in combination with low B12, with neural tube defects (Wilson et al., 1999). Certain combinations of MTRR A66G and MTHFR A1298C genotypes were associated with risk for hepatocellular carcinoma (Kwak et al., 2008). Several studies found no association of the MTRR A66G polymorphism with colon cancer risk (Steck et al., 2008; Eussen et al., 2010), but a study by Matsuo et al. (2002) reported that MTRR 66 GG genotype acted as a risk factor for colorectal cancer in Japanese. Our results, demonstrating a possible protective role of the MTRR AA genotype (OR = 0.655; 95% CI = 0.441–0.973; p = 0.04), might point in the same direction, although we could not demonstrate a statistically significant risk effect of either AG or GG genotype in our population. Our results indicate that wild-type MTRR 66 AA genotype might have a role in preventing colon carcinogenesis, possibly through proper regulation of MTR activity, which might consequently influence SAM levels and thus DNA methylation reactions.
In this study, we observed no significant difference in genotype frequencies among all investigated polymorphisms between males and females. This might be due to a relatively small sample number when the investigated individuals were divided by sex. The other limitation of our study is its relatively small power, which was observed when we set an OR of 1.5 for the polymorphisms being studied. On the other hand, we investigated a population that is ethnically homogenous, and to our knowledge, not many similar studies on Slavic populations were performed, which we consider to be a strength of this study.
Maximum likelihood analysis of haplotype distributions between MTHFR A1298C and MTRR A66G, located on chromosomes 1 and 5, respectively, revealed LD only in the patient group. This is an interesting finding because, in most cases, LD is detected between linked loci. The decay of disequilibrium depends on the recombination rate between loci. If loci are tightly linked (located on the same chromosome), LD decays very slowly. If they are not linked (located on different chromosomes), as is the case here, LD decays much faster (it is halved in each generation). Therefore, the presence of significant LD between MTHFR A1298C and MTRR A66G suggests the existence of a relatively strong selection that maintains it. However, it is not possible to distinguish exactly what kind of natural selection is involved in the maintenance of LD between MTHFR A1298C and MTRR A66G. One possibility is that LD is maintained due to natural selection at the level of gametes. Such mechanism is, however, unlikely to be the directly linked with colon cancer susceptibility, because colon cancer appears at later age. Another possibility is that selection could act on viability at the genotypic level (from zygote to adult).
Analysis of estimated frequencies of haplotypes of these two loci showed that the frequency of MTHFR 1298 A/MTRR 66 G haplotype was higher in cases than in controls, resulting with an OR of 1.270 (95% CI = 1.007–1.602; p = 0.04). On the other hand, MTHFR 1298 A/MTRR 66 A haplotype was shown to be protective (OR = 0.775; 95% CI = 0.603–0.996; p = 0.04), whereas MTHFR 1298 C/MTRR 66 A and MTHFR 1298 C/MTRR 66 G haplotypes were both found to be neutral (their frequencies did not differ statistically between the groups). Further, we have observed that combinations of genotypes harboring MTHFR 1298 A/MTRR 66 G risk haplotype and no MTHFR 1298 A/MTRR 66 A protective haplotype were more frequent in cases (Fig. 1). On the other hand, genotypes harboring MTHFR 1298 A/MTRR 66 A protective haplotype, but not MTHFR 1298 A/MTRR 66 G risk haplotype, were more frequent in controls (Fig. 1). However, no specific genotype combination could be statistically proven to act as a risk or protective factor for colon cancer, which might be related to insufficient statistical power of our sample for such analysis.
Regarding the functional consequence of these two SNPs, MTHFR Glu429Ala, which results from A1298C, is located near the binding site for SAM, the allosteric inhibitor of MTHFR (Curtin et al., 2004). As MTRR influences homocysteine conversion to methionine, which in turn converts into SAM, A66G may influence SAM production, which could change MTHFR feedback inhibition.
In conclusion, the obtained results indicate the role of MTHFR A1298C and MTRR A66G polymorphisms in colon carcinogenesis. Additional studies including folate intake, diet, and lifestyle should be conducted to fully understand the associations between polymorphisms in folate-metabolizing enzymes and colon carcinogenesis.
Conclusion
Our results have confirmed, on a new and ethnically homogenous sample, the role of MTRR A66G polymorphism in the etiology of colon cancer. Moreover, we showed that polymorphisms MTRR A66G and MTHFR A1298C combined influence colon cancer risk, but further studies are needed to fully elucidate the mechanism by which they are linked.
Footnotes
Acknowledgment
This study was supported by a grant (098-0982464-2508) from the Ministry of Science, Education, and Sports of the Republic of Croatia.
Disclosure Statement
The authors confirm that no conflict of interest exists in this study.