
Abstract
Caveolin-1 (CAV-1) is a key structural component of caveolae that regulates cholesterol. Employing transgenic techniques to regulate the cholesterol content of pork through CAV-1 is hindered by our lack of knowledge about its regulation. To investigate the regulatory mechanism of porcine CAV-1, a DNA segment containing the 5′-flanking region of CAV-1 was isolated from porcine genomic DNA and sequenced. The luciferase reporter assay detected five cis-acting elements for efficient expression of the CAV-1 gene at the region spanning nucleotides −213 to −20 with serially deleted 5′-flanking sequences and site-directed mutants, −123 to −114 was the core promoter. The electrophoretic mobility shift assay demonstrated potential binding of Sp1 protein to this core promoter. The purpose of this study is to systematically elucidate the transcriptional regulation mechanism of porcine CAV-1 and to contribute to the investigation of the interaction between CAV-1 and cholesterol.
Introduction
There are two sources of cellular cholesterol: de novo cholesterol synthesis and dietary cholesterol, with an estimated ratio of ∼70:30 (Ikonen, 2008). Animal products, particularly pork, contribute the majority of the saturated fat and cholesterol content of our daily diet. Decreasing the total fat content or substituting unsaturated vegetable oils for saturated fats could result in reduced total plasma and low-density lipoprotein-cholesterol concentrations (Mattson and Grundy, 1985; Khosla and Hayes, 1992; Kasim et al., 1993; Ginsberg et al., 1994). However, consumers do not always limit their consumption of animal products such as pork, and therefore, an alternative approach would be to modify the fatty acid composition of animal products (Stewart et al., 2001).
CAV-1, which was identified as the major coat protein of caveolae (Couet et al., 2001), plays an important role in intracellular cholesterol homeostasis (Murata et al., 1995; Puglielli et al., 1995; Smart et al., 1996; Uittenbogaard et al., 1998). CAV-1 can modulate signal transduction by attracting signaling molecules to caveolae and regulating their activity. Studies support an inhibitory role for CAV-1 in nitric oxide synthase enzymatic activity (Garcia-Cardena et al., 1997; Ju et al., 1997; Ghosh et al., 1998). The activation of Ras-p42/44 MAP kinase signal transduction pathway inhibits CAV-1 gene expression, whereas elevated CAV-1 protein levels strongly inhibit this pathway (Engelman et al., 1998a, 1999). A similar situation has been observed with the Neu-mediated signaling pathway (Engelman et al., 1998b).
CAV-1 is linked to virus replication and entry into cells. It supports the human influenza virus A life cycle (Llano et al., 2002; Sun et al., 2010). Phosphorylated CAV-1 is involved in respiratory syncytial virus assembly (Brown et al., 2002). The association of rotavirus and CAV-1 may contribute to rotavirus intracellular trafficking from the endoplasmic reticulum to the cell surface (Parr et al., 2006). Hepatitis B virus requires a CAV-1–mediated entry pathway to initiate productive infection in HepaRG cells (Macovei et al., 2010). CAV-1 also facilitates delivery of simian virus 40 (Anderson et al., 1996; Pelkmans et al., 2001).
Given the importance of CAV-1 in the regulation of cholesterol and other pathophysiological processes, its transcriptional control is of considerable interest. To investigate the transcriptional regulation of porcine CAV-1, the 5′-flanking sequence of the gene was cloned and Sp1 was identified as the key transcription factor.
Materials and Methods
Isolation of the CAV-1 gene 5′-flanking region
BLAST with a section of the porcine CAV-1 cDNA sequence (GenBank: NM_214438), in the high-throughput genomic sequence GenBank database, identified a contig containing the pig CAV-1 DNA sequence (GenBank: DP000017.1). Approximately 2-kb 5′-flanking sequence of the gene was amplified using primers (Table 1, C1proF/R) and confirmed via sequencing. The transcriptional start site of porcine CAV-1 was predicted by analyzing the cDNA sequence (GenBank: NM_214438) with two online tools (
Nucleotides underlined are restriction enzyme sites.
Reverse primer.
Including 23 nt from pGL3-Basic vector.
Porcine probe designed on the basis of mutagenesis experiment result.
Standard Sp1 probe.
Mutated standard Sp1 probe.
CAV-1, caveolin-1; EMSA, electrophoretic mobility shift assay.
Construction of promoter reporter plasmids and mutagenesis
A series of plasmids containing fragments of various sizes from the 5′-flanking region of the porcine CAV-1 gene were constructed by inserting DNA fragments between the MluI and XhoI sites of the pGL3-basic vector (Promega). 5′-end–deleted DNA fragments were obtained using PCR with synthetic oligonucleotides incorporating a 5′-end MluI site and a 3′-end XhoI site (Table 1, primers for pGLB-1756∼260). The amplified DNA fragments were ligated with pGEM-T Easy vector (Promega), digested with MluI and XhoI, and inserted immediately upstream of the luciferase reporter gene. The nested deletion technique using Erase-a-base (Promega) was employed for further analysis. Targeted mutagenesis to inactivate cis-elements was performed using the QuikChange XL and Multisite-Directed Mutagenesis Kit (Stratagene) with the designated primers (Table 1). The sequences and orientations of the constructs were verified by sequencing.
Cell culture
There are no commercially available pig muscle cell lines. Thus, we selected the immortalized murine myoblast cell line C2C12 for our project. C2C12 cells were maintained in Dulbecco's modified Eagle's medium supplemented with antibiotics and 10% (v/v) fetal bovine serum (Hyclone).
Transient transfection and luciferase reporter gene assays
Transient transfection of C2C12 cells with CAV-1 promoter/firefly luciferase reporter plasmids was conducted using Lipofectamine™ 2000 transfection reagent (Invitrogen). Thirty hours after transfection, the cells were lysed and assayed for promoter activity using the Dual-luciferase reporter assay system (Promega). The luciferase activity was measured with a luminometer Centro LB 960 (Berthold). To normalize for transfection efficiency, the cells were cotransfected with 6 ng of pRL-TK vector (Promega).
Electrophoretic mobility shift assay
Nuclear extracts were prepared using a Nuclear Extract Kit (Active Motif). A probe extending from nucleotide position −130 to −105 was labeled with 32P (Table 1, PSp1). Nuclear extracts were incubated with this radioactive oligonucleotide probe containing the important putative element. Poly-dIC was added to prevent nonspecific binding of proteins to the oligonucleotide probe and reduce background noise. After binding, the samples were separated on a nondenaturing sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel and bands were detected by autoradiography. Cold competition analysis was performed using unlabeled PSp1 oligonucleotides in 50- and 100-fold molar excess (Table 1, PSp1). The human Sp1 binding sequence was employed as a standard control (Table 1, SSp1). Sp1-specific antibody was added to the binding reactions to perform super gel shift experiments.
Statistical analysis
Data are expressed as mean ± SE. The significance of observed differences between groups was determined by Student's t-test.
Results
Cloning of the 5′-flanking sequence of CAV-1
Primers were designed to amplify the full-length 5′-flanking region based on a contig (GenBank: DP000017.1) containing the complete porcine CAV-1 sequence (Fig. 1). Relative to the putative transcription start site (numbered + 1, predicted by online analysis tools), a 1790-bp (−1756/ + 34) fragment of the putative CAV-1 promoter region (Fig. 1) was cloned into the pGL3-basic firefly luciferase reporter plasmid. After transfection into C2C12 cells and examination using the dual luciferase reporter assays, this putative CAV-1 promoter fragment showed ∼16-fold higher activity than pGL3-Basic (Fig. 2A). To determine the minimal functional region, serial 5′ truncations were conducted. Although promoter activities varied, the major cis-regulatory elements that mediated promoter activity were located within the (−213/ + 34) fragment (Fig. 2A). Further deletion of this region indicated that two important regions (−213/−178 and −149/−109) were required for the activity of the CAV-1 promoter (Fig. 2B).

Structure of the porcine CAV-1 gene promoter. A 1790-bp genomic clone of porcine CAV-1 was sequenced and analyzed for consensus sequences of known mammalian transcription factors. The putative transcription initiation site, a guanine, is designated the + 1 nucleotide, and the remainder of the sequence is numbered relative to this. Overlined sequences correspond to conservative homologous matches to the noted transcription factor or promoter element. Underlined sequences indicate primers for pGLB-1756∼260 constructs.
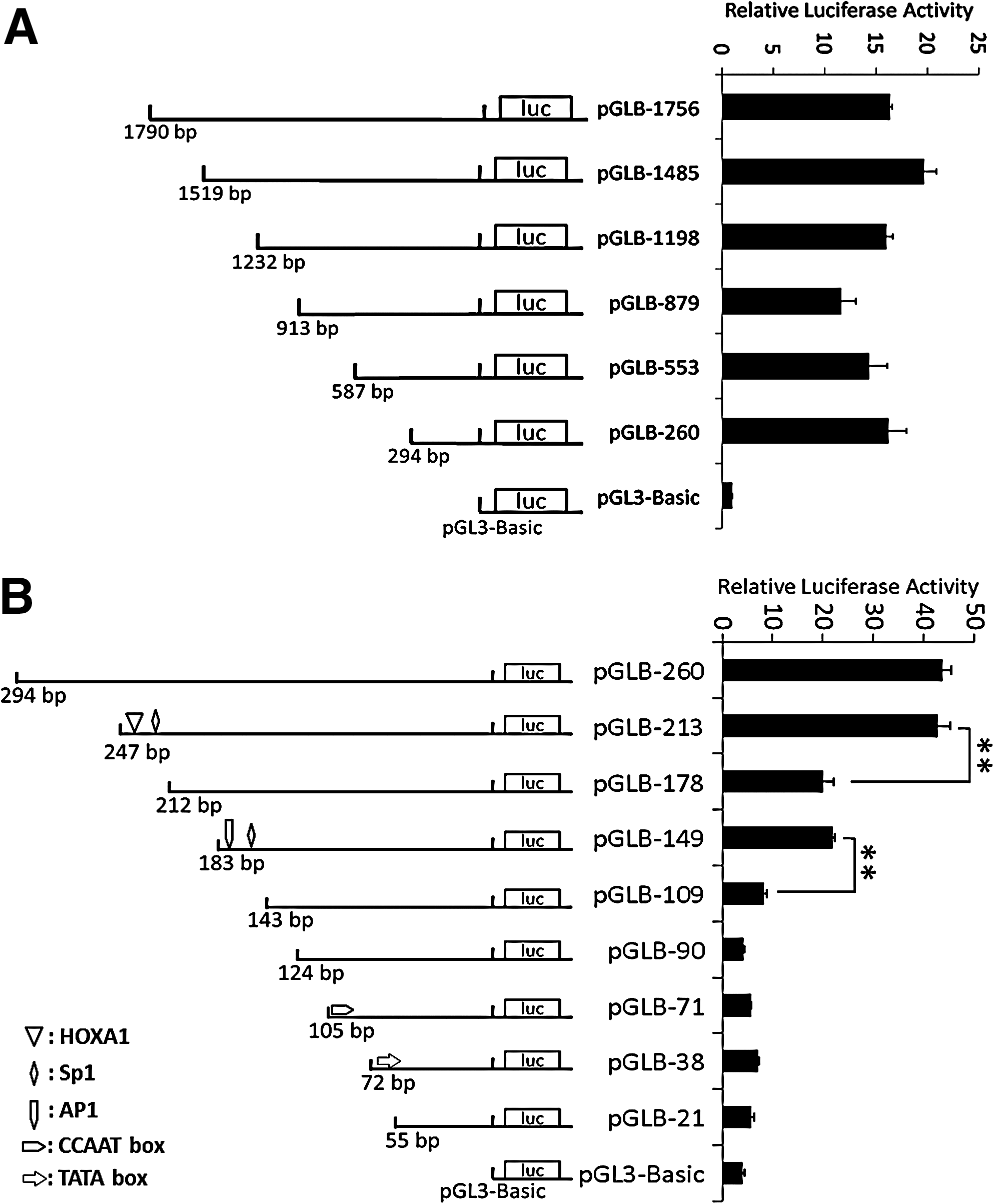
The 5′ deletion analysis of the promoter activity of porcine CAV-1. The 5′ truncated fragments of the CAV-1 promoter were inserted into luciferase reporter vectors, followed by transfection into the C2C12 cell line. The pRL-TK vector encoding Renilla luciferase was cotransfected as an internal reference, and luciferase values were normalized to Rluc activity. The x-axis indicates fold-activation to normalized luciferase activities. The results are represented as means ± SE of at least three independent experiments. **p < 0.01.
Transcriptional regulation of CAV-1
To identify conserved putative binding sites for transcription factors, the genomic sequences of the porcine CAV-1 proximal promoter were analyzed using the TESS and TFSEARCH online tools. The results suggested that nucleotides −213/−178 harbored a HOXA1 binding site and an Sp1 binding site (named Sp1.1), and an AP1 binding site and another Sp1 binding site (named Sp1.2) were identified in the −149/−109 region. A classical TATA box motif (−30/−20) and a CCAAT box (−65/−55) were also present.
To determine the functional importance of the predicted regulatory motifs, the core consensus sequences of the predicted sites were altered by mutagenesis of the CAV-1 promoter constructs pGLB-213 (−213/ + 34) or pGLB-149 (−149/ + 34). C2C12 cells were transfected with the plasmids and promoter activities determined. Using pGLB-213 as template, mutations of the HOXA1 binding site, Sp1.1, or AP1 binding sites resulted in a significant 56%, 55%, or 32% reduction in promoter activity, respectively (p < 0.01), and promoter activity was significantly reduced by about 90% after mutation of Sp1.2 (p < 0.01). Using pGLB-149 as template, a significant 95% decrease of the promoter activity was observed after mutation of Sp1.2 (p < 0.01). These results suggest that all these four sites are functional and Sp1.2 is crucial for CAV-1 activity (Fig. 3A, B). Similarly, mutation of the CCAAT box sequence resulted in a significant (78%) loss of promoter activity (p < 0.01). However, we did not observe a significant change in promoter activity when the putative TATA box was mutated (Fig. 3A). Double mutants with mutated AP1 binding site and Sp1.1, or the CCAAT box and TATA box, demonstrated significant lower promoter activity than single-site–mutated mutants (p < 0.01 or p < 0.05; Fig. 3A). These results confirmed the importance of these cis-elements in the promoter activity of CAV-1.
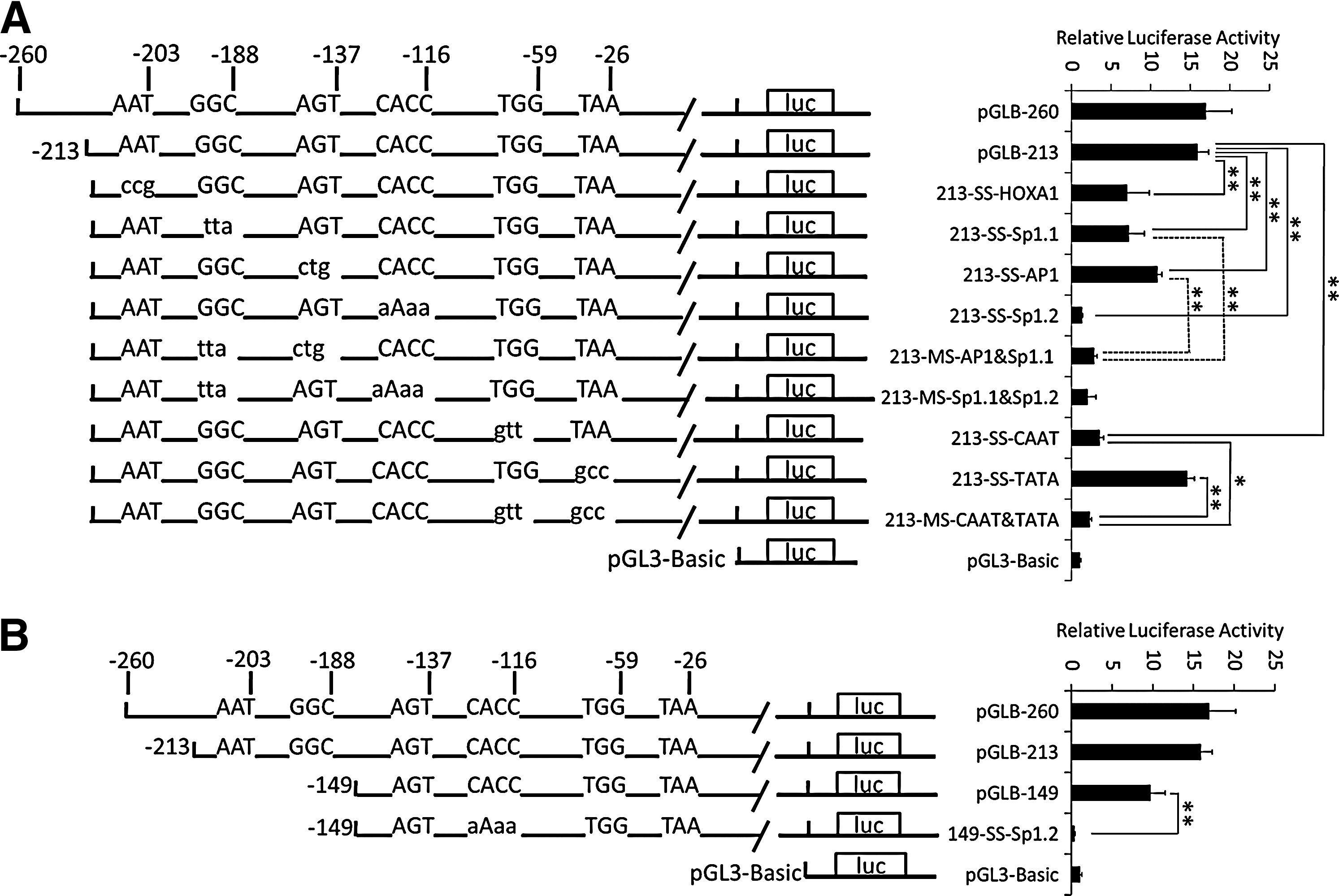
Impact of important potential binding motifs within the core promoter on porcine CAV-1 transcription. Site-directed mutation was carried out to generate several mutated plasmids, followed by transfection into the C2C12 cell line. The pRL-TK vector encoding Renilla luciferase was cotransfected as an internal reference, and luciferase values were normalized to Rluc activity. The x-axis indicates fold-activation to normalized luciferase activities. Mutated elements and changed nucleotides are depicted. Uppercase letters represent the wild-type sequence and lowercase letters represent mutation. The results are represented as means ± SE of at least three independent experiments. *p < 0.05; **p < 0.01.
Sp1 protein binds to the −123/−114 region
Sp1 is involved in the regulation of CAV-1 expression (Bist et al., 1997, 2000; Cao et al., 2005; Dasari et al., 2006). Our study demonstrated that the −123/−114 region was crucial for promoter activity, and Sp1 protein was predicted to bind to this site. Thus, electrophoretic mobility shift assays (EMSA) were performed using the corresponding sequence from the CAV-1 proximal promoter as an EMSA probe (Fig. 4A, B). A probe containing the Sp1 consensus sequence was used as a positive control. Nuclear extracts from C2C12 cells interacted with the oligomer probes to form complexes. The specificities of these complexes for the sequence were demonstrated in a competition experiment in which the complexes were competed with a 100-fold excess of unlabeled wild-type probe. Addition of an anti-Sp1 antibody caused the band to become less intense.
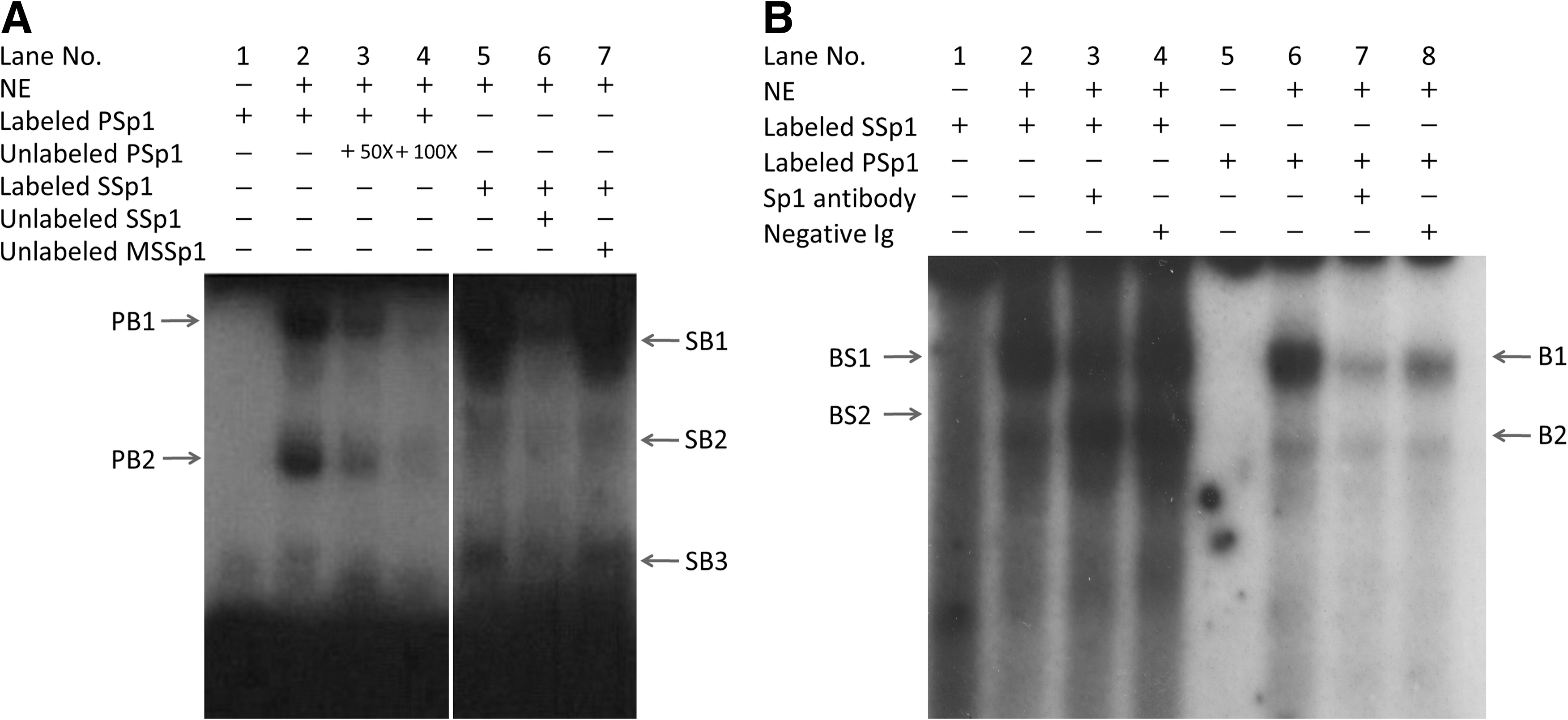
Sp1 from C2C12 nuclear protein extracts binds to the wild-type porcine CAV-1 promoter −123/−114 region. An antisense oligonucleotide was utilized; EMSA was performed with radioactive labeled oligonucleotides including PSp1, which corresponds to region −123/−114 of the porcine CAV-1 promoter, a standard probe SSp1, which contains the Sp1 consensus sequence, and a mutated standard probe, MSSp1.
Discussion
The function of CAV-1 has been studied extensively since its identification as the marker protein of caveolae in the early 1990s (Couet et al., 2001). However, the transcriptional regulation of this gene, particularly in pigs, remains unknown. Regulatory analyses of the porcine CAV-1 gene promoter are reported in this study and reveal a structure characterized by the absence of an active TATA-box sequence but having several potential regulatory elements (Fig. 1). These include several functional elements including putative AP1 and HOXA1 binding sites; one CCAAT-box sequence, which is an important element for transactivation of CAV-1; and one of two functional Sp1 binding elements (−123 to −114, named Sp1.2) that is crucial for promoter activity.
The promoter structure of the porcine CAV-1 gene is similar to that of bovine CAV-1. They share nearly 80% identity in the 900-bp sequence upstream to transcription start site. They are both characterized by absence of active TATA box and the presence of various Sp1 sites. Although the human CAV-1 promoter is substantially different from that of the pig (∼45% identity), there are several Sp1 response sites in the promoter region (Bist et al., 1997). The similarity within this proximal promoter region highlights its importance in regulating the transcription of CAV-1 homologs and suggests that their transcriptional regulation is likely achieved through similar mechanisms.
Sp1 family proteins are ubiquitously expressed in many tissues and they can promote the assembly of transcription complexes by interacting with transcription factors (Ellwood et al., 1999). Sp1 functions synergistically with sterol-responsive element (SRE)-binding proteins (SREBP) to activate transcription of target genes in response to cholesterol depletion (Sudhof et al., 1987; Brown and Goldstein, 1997; Naar et al., 1998). More recently, complementary and alternative coregulatory proteins have been identified, which influence SREBP/Sp1 co-operation, suggesting additional complexities to this transcriptional paradigm (Cao et al., 2005). The putative response element for SREBP is required for human CAV-1 promoter activity (Bist et al., 1997; Cao et al., 2005). However, no putative SRE elements have been identified in the porcine CAV-1 promoter region −1756/ + 34.
In some studies investigating the regulation of CAV-1 expression, as transcriptional regulation was considered important, whereas several studies have reported that FOXO and c-myc could regulate the transcription of CAV-1 (Park et al., 2001; van den Heuvel et al., 2005; Roy et al., 2008). It was proposed that ERM and PEA3 bind the CAV-1 promoter in the mouse E10 cell line (Kathuria et al., 2007), and p53 and E2F/DP-1 have been implicated in the transcriptional regulation of CAV-1 (Engelman et al., 1999; Fielding et al., 1999; Bist et al., 2000). Thus, we conclude that expression of pig CAV-1 is synergistically activated by Sp1 and other factors and that Sp1 protein could play a crucial role in promoter activity.
Conclusions
In summary, this study demonstrated that the region spanning nucleotides −213 to +34 of porcine CAV-1 was sufficient for basal transcription. The region −123/−114 was crucial for transactivation of the promoter, and four other functional cis-elements were identified. The transcription factor Sp1 interacts with nucleotides −123/−114, indicating that Sp1 could play a key role in the transcriptional regulation of pig CAV-1. Further research is required to investigate this regulatory mechanism, and a comprehensive understanding will aid in further elucidation of physiopathological processes related to CAV-1 including the transport and metabolism of cholesterol.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (30700568), the National Natural Science Foundation of China-Guangdong Key Project (U0731003), and the National High Science and Technology Foundation of China (2006CB102101).
Disclosure Statement
No competing financial interests exist.