
Abstract
Proto-oncogenes are involved in cell growth, proliferation, and differentiation. In the present study, we investigated the roles and mediating pathways of proto-oncogenes c-erbB2 and c-myb in mouse oocyte maturation by RT-PCR, real-time quantitative PCR, western blot, and recombinant proto-oncogene protein microinjection. Results showed that both c-erbB2 and c-myb antisense oligodeoxynucleotides (c-erbB2 ASODN and c-myb ASODN) inhibited germinal vesicle breakdown and the first polar body extrusion in a dose-dependent manner. However, microinjection of recombinant c-erbB2 or c-myb protein into germinal vesicle stage oocytes stimulated oocyte meiotic maturation. In addition, the expression of c-erbB2 and c-myb mRNA was detected in oocytes; and c-erbB2 ASODN and c-myb ASODN inhibited c-erbB2 mRNA and c-myb mRNA expression, respectively. Maturation promoting factor (MPF) inhibitor roscovitine did not affect the expression of c-erbB2 mRNA and c-myb mRNA, but blocked the effects of recombinant c-erbB2 and c-myb protein-induced oocyte maturation. Further, cyclin B1 protein expression in oocytes was remarkably inhibited by c-erbB2 ASODN, c-myb ASODN, and roscovitine. Nonsense tat ODN had no effect on the expression of c-erbB2 , c-myb, and cyclin B1. These results suggest that c-erbB2 and c-myb may induce oocyte maturation through mediating a pathway involving the activation of MPF.
Introduction
Oocyte meiotic maturation involves several processes including disassembly of the nucleus, chromosome condensation, cytoskeletal rearrangements, and arrest in transcriptional activity, which are associated with the activation of maturation promoting factor (MPF), a key step that catalyses entry into meiosis I and meiosis II during oocyte maturation. MFP is a complex consisting of a catalytic p34cdc2 subunit and a regulatory cyclin B subunit, and it is maintained in an inactive state through phosphorylation of residues Tyrl5 and Thr14 of the p34cdc2 kinase. MPF activity is necessary for resumption of meiosis, and it requires phosphorylation of cyclin B1 (Peter et al., 2002). MPF regulators are the focus of much reproductive physiology research. Proto-onceongene c-mos is the most important regulator of MPF (Sagata et al., 1989, 1998; Colledge et al., 1994; Ohashi et al., 2003; Lazar et al., 2004); it enhances MPF activity via MAPK activiation and/or inhibiting proteolytic degradation of cyclin B, resulting in the maintenance of high MPF activity during metaphase arrest (Heikinheimo et al., 1998). Whether other proto-oncogenes besides c-mos, are involved in regulating MPF activity is an open question. In the present study, we investigated the effects of c-erbB2 and c-myb on mouse oocyte maturation to advance our understanding of proto-oncogenes on female reproduction. In addition, we explored the relationship between the two proto-oncogenes and MPF activity.
Materials and Methods
Chemicals
Antisense oligodeoxynucleotides: c-erbB2 antisense oligodeoxynucleotide (c-erbB2 ASODN) sequence is 5′-CAGCTCCATGGTGCT-3′, with an effect on the 5′ coding region in c-erbB2 mRNA; c-myb antisense oligodeoxynucleotide (c-myb ASODN) sequence (Simons and Rosenberg, 1992; Simons et al., 1992) is 5′-GTGTCGGGGTCTCCGGGC-3′, with an effect on the 4th–22nd nucleotide in mouse c-myb mRNA translation region. HIV-1 trans-activating transcriptional activator (TAT) was used as a nonsense control, and it definited the oligodeoxyribonucleotide that targeted the front 15 basyl of the second extron in the TAT sequence (sequence, 5′-CATTTCTTGCTCTCC-3′). All ASODNs were synthesized and purchased from Shanghai biological engineering center, and they were modified by phosphorothioic acid.I to increase permeability to cell membrane and to prevent nucleotidase hydrodissociation. Recombinant human c-erbB2 protein and recombinant human c-myb protein were purchased from Abnova Corporation. The other chemicals used in this experiment were purchased from Sigma Chemical Company.
Oocyte collection and culture
Kunming strain mice (4–6-week-old) were purchased from Laboratory Animal Core of Nanchang University (Jiangxi province, China). Mice were raised and handled in accordance with guidelines of animal care. Germinal vesicle (GV) stage oocytes were isolated and collected from ovaries by puncturing antral follicles using a 26-gauge needle 48 h after injection of 10 IU equine chorionic gonadotropin. Oocytes were washed three times and cultured in medium 199 supplemented with 2 mM L-glutamine, 10% fetal calf serum, 60 μg/mL penicillin, and 50 μg/mL streptomycin. All cultures were maintained at 37°C in a humidified atmosphere of 5% CO2.
Treatment and examination of oocytes
At the beginning of culture, 120 oocytes were treated with c-erbB2 ASODN and c-myb ASODN, respectively. In the second experiment, 26 oocytes were microinjected with recombinant c-erbB2 protein (recom-c-erbB2 , 5–20 μg/mL) and c-myb protein (recom-c-myb, 5–20 μg/mL), respectively; then, the oocytes were treated by MPF inhibitor roscovitine (50 μM) (Deng and Shen, 2000). After treatments, oocytes were observed for GV status and presence or absence of the first polar body (PB1) at different time points with a Nikon invert microscope. The number of oocytes in germinal vesicle breakdown (GVBD) stage and the number of oocytes with the PB1 extrusion formation were recorded. All experiments were repeated at least three times.
RT-PCR assays
RNA was isolated from 250 oocytes with Trizol Reagent (Life Technologies, Inc.), quantified by measuring absorbance at 260 nm, and stored at −80°C until assay. Total RNA (2 μg) was reverse transcribed to first-strand cDNA with 200 U SupeScript III RNase H-Reverse Transcriptase (Life Technologies, Inc.) in the presence of 0.5 mM deoxy-NTPs and 25 μg/mL oligo (deoxythymidine) in a total volume of 20 μL for 60 min at 50°C.
The c-erbB2 primers were designed as follows: (forward primers 5′ AGCTGCACTGTGGATGTCAG 3′ and reverse primers 5′ GAGCCTTCGGCACTGTCTAC 3′) according to mouse the c-erbB2 cDNA sequence (GenBank accession number: NM 001003817). The c-myb primers were designed as follows: (forward primers 5′ TTTCCTGTTTGGGAGAACACCATGCCACCCC 3′ and reverse primers 5′ GGAGTTCGAGGAGAGCTTTCGAGGATTGACC 3′) according to the mouse c-myb cDNA sequence (GenBank accession number: NM 033597). The β-actin transcript was amplified as an internal control. The primers for mouse β-actin were designed (forward primers 5′ ACACTGTGCCCATCTACGAGG 3′ and reverse primers 5′ AGGGGCCGGACTCGTCATACT 3′) according to the mouse β-actin cDNA sequence (GenBank accession number: XM 134663). Amplification was carried out under the following conditions: denaturation at 94°C for 60 s, annealing at 55°C for 55 s, and extension at 72°C for 60 s, 36 cycles. The PCR products were analyzed by electrophoresis on 1.5% agarose gels containing ethidium bromide and photographed under UV illumination. The relative integrated density of each band was digitized by multiplying the absorbance of the surface area. The mRNA levels of c-erbB2 and c-myb were normalized against β-actin expression. All experiments were repeated at least three times.
Real-time PCR
To analyze c-erbB2 and c-myb mRNA levels, total RNA was extracted from cultured mouse oocytes and used as a tem-plate for cDNA synthesis using oligo (dT) primers with the superscriptIII kit (Invitrogen). Total DNA was extracted from the mouse oocytes to assess GFP DNA levels. Real-time quantitative PCR was performed using the ABI Prism 7500 detection system (PE Applied Biosystems) with SYBR green DNA detection kit (Applera). The expression levels of β-actin were used for normalization. The PCR primers for c-erbB2 were as follows: forward primer 5′ CAGCTCCAAGTGTTCGAAACC 3′, reverse primer 5′ TGGCCATGCTGAGATGTACAG 3′. The PCR primers for c-myb were as follows: forward primer 5′ AAAGCTCGGGCTTAGATGCA 3′, reverse primer 5′ TGGTCTCGACATGGTGTCAGTT 3′. The PCR primers for β-actin were as follows: forward primer 5′ TTCAACACCCCAGCCATGT 3′, reverse primer 5′ CAGAGGCATACAGGGACAACAC 3′. All experiments were repeated at least three times.
Western blot analysis
One hundred and fifty oocytes were collected in SDS sample buffer and heated to 100°C for 4.5 min. The proteins were separated by SDS-PAGE with a 5% stacking gel and a 10% separating gel, electrically transferred to nitrocellulose membrane, and blocked for 2 h at room temperature in Tris-buffered saline Tween-20 (TBST) containing 5% nonfat dry milk. The membrane was incubated overnight at 4°C with monoclonal mouse anti-cyclin B1 antibody (Santa Cruz Biotechnology Inc.) diluted 1:100 in TBST. After washing thrice in TBST, the membrane was incubated for 1 h at 37°C with horseradish peroxidase-conjugated goat anti-mouse IgG (Santa Cruz Biotechnology Inc.) diluted 1:1000 in TBST. The membrane was washed three times in TBST and then processed using the ECL detection system (Amersham International). All experiments were repeated at least three times.
Microinjection of recombinant c-erbB2 and c-myb protein into oocytes
Recombinant c-erbB2 protein (5–20 μg/mL), recombinant c-myb protein (5–20 μg/mL), or rabbit IgG was dissolved in 50 mM Tris-acetate (pH 7.5) and was microinjected into the cytoplasm of 26 fully grown GV oocytes with an Eppendorf microinjector (Nikon). All microinjection procedures were as previously described by Dai (2000). Dibutyryl cyclic adenosine monophosphate (0.1 μM) was added to the medium to prevent GVBD. A microinjection volume of about 8 pl per oocyte was used in all the experiments. The same amount of mouse IgG diluted in PBS was injected as control. After microinjection, eggs were thoroughly washed with M199 medium and cultured in the same medium. All experiments were repeated at least three times.
Statistical analysis
All data are presented as the mean±SD (n=3). The statistical differences among the groups were determined by analysis of variance using SPSS standard version 11.0 (SPSS, Inc). p-Value of <0.05 was considered statistically significant.
Results
Effect of c-erbB2 ASODN on oocyte maturation
To examine the effect of c-erbB2 on mouse oocyte maturation, oocytes were treated with different doses of c-erbB2 ASODN (0–20 μM). As shown in Figure 1, c-erbB2 ASODN significantly inhibited oocyte meiotic maturation in a dose-dependent manner. The percentage of GVBD and the percentage of PB1 extrusion were reduced compared with the control, and the lowest percentage (GVBD 34.6%; PB1 16.5%) was observed after culture for 16 h with 20 μM c-erbB2 ASODN. In contrast, nonsense TAT ODN (20 μM) had no inhibiting effect on oocyte maturation during the 8–16 h culture period. At the same time, both RT-PCR and Real-time PCR analysis verified that treatment with c-erbB2 ASODN (20 μM) for 8 h inhibited the expression of c-erbB2 mRNA. However, nonsense TAT ODN (20 μM) did not have the same effect (Fig. 2).
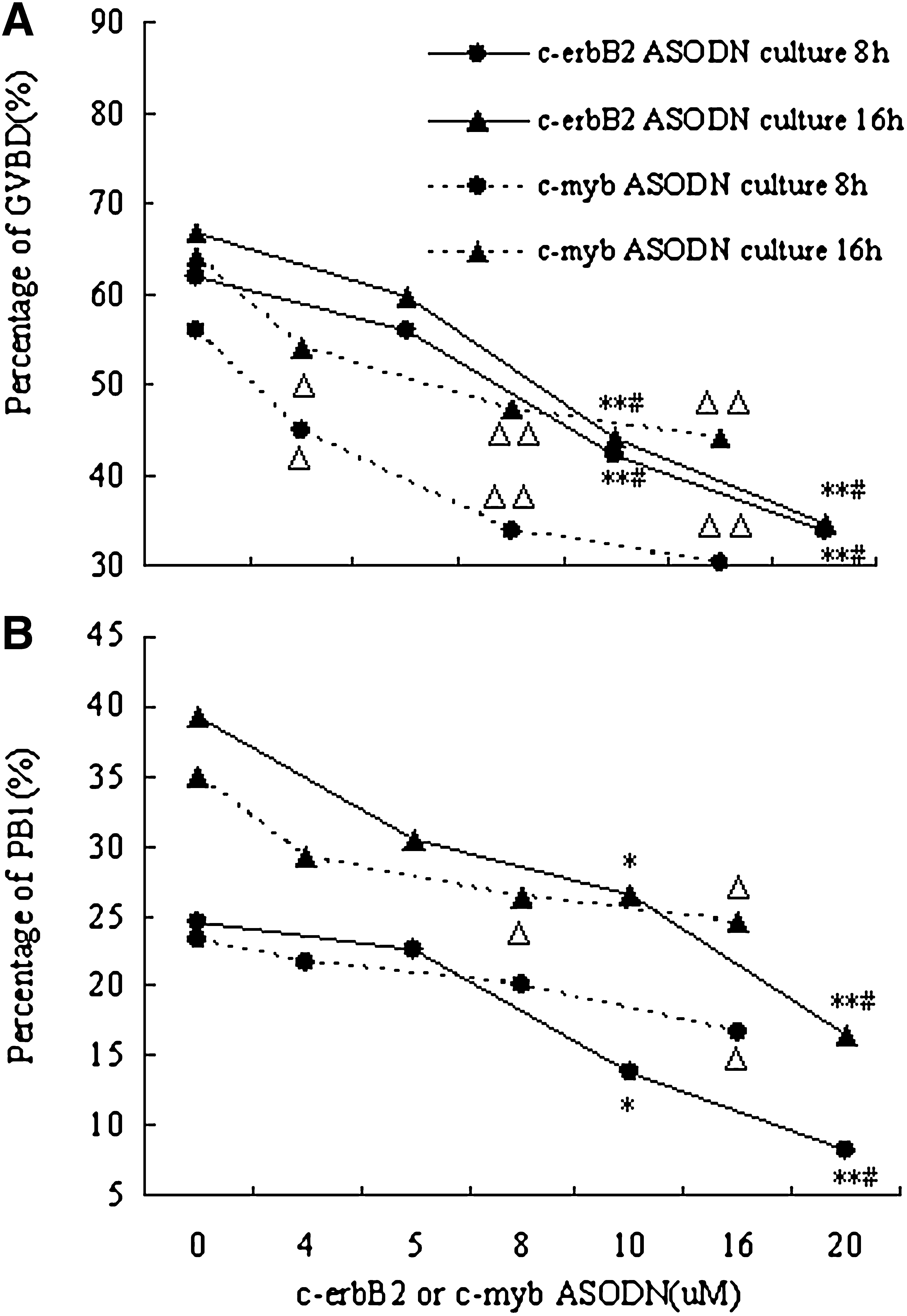
The dose-dependent effects of c-erbB2 ASODN and c-myb ASODN on oocyte GVBD and PB1 extrusion. *p<0.05, **p<0.01 compared with group 0 μM c-erbB2 ASODN, # p<0.05 compared with group 5 μM c-erbB2 ASODN. Δ p<0.05, ΔΔ p<0.01 compared with group 0 μM c-myb ASODN. ASODN, antisense oligodeoxynucleotides; GVBD; germinal vesicle breakdown; PB1, first polar body.
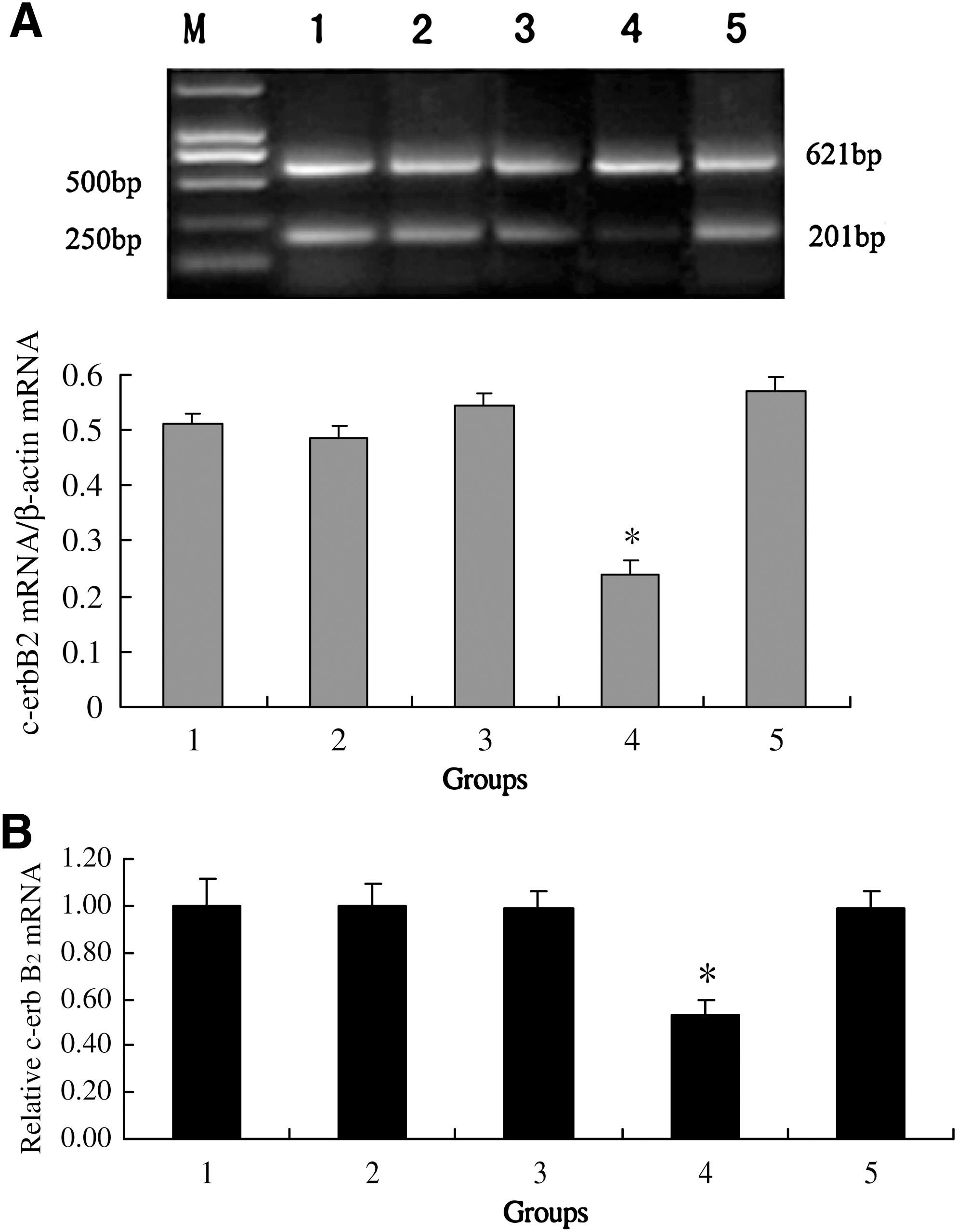
Effects of c-erbB2
ASODN, c-myb ASODN, and roscovitine on c-erbB2
mRNA expression in mouse oocytes after 8 h culture.
Effect of c-myb ASODN of on oocyte maturation
As shown in Figure 1, after culture for 8–16 h, oocyte meiotic maturation was significantly inhibited by treatment with c-myb ASODN (0–16 μM) compared with the control, and the inhibition was enhanced by increasing c-myb ASODN concentration. The maximal effect (GVBD 44.12%; PB1 24.51%) was observed at the highest concentration evaluated (16 μM). Further, both RT-PCR and Real-time PCR analysis revealed that inhibition of oocyte maturation induced by c-myb ASODN (16 μM) was accompanied by reduced c-myb mRNA levels. However, nonsense TAT ODN (20 μM) had no inhibiting effect on c-myb mRNA expression (Fig. 3).
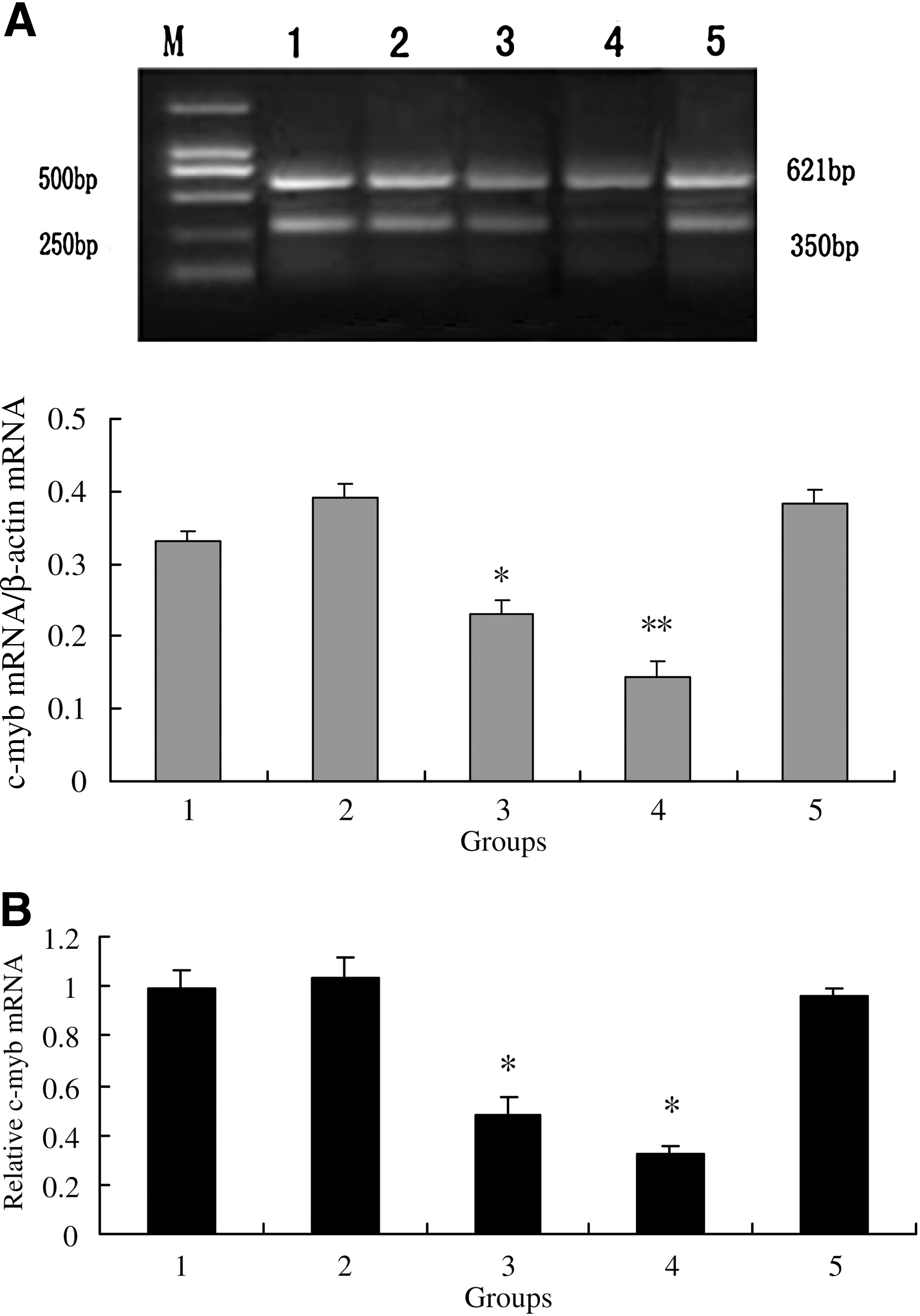
Effects of c-erbB2
ASODN, c-myb ASODN, and roscovitine on c-myb mRNA expression in mouse oocytes after 8 h culture.
Effects of c-erbB2 and c-myb protein microinjection on oocyte maturation
Compared with the control (GVBD 40.9%; PB1 22.7%), microinjection of 5, 10, 20 μg/mL recombinant human c-erbB2 protein into resting oocytes significantly promoted GVBD in a dose-dependent manner after 6 h culture (GVBD 46.5%, 64.0%, 67.0% respectively), and it also stimulated PB1 extrusion in a dose-dependent manner after 12 h culture (PB1 24.3%, 40.0%, 44.2% respectively). Likewise, microinjection of 5, 10, 20 μg/mL recombinant human c-myb protein promoted GVBD in a dose-dependent manner after 6 h culture (GVBD 53.4%, 73.1%, 76.2% respectively), and also stimulated PB1 extrusion in a dose-dependent manner after 12 h culture (PB1 27.3%, 46.2%, 49.5% respectively) (Fig. 4).
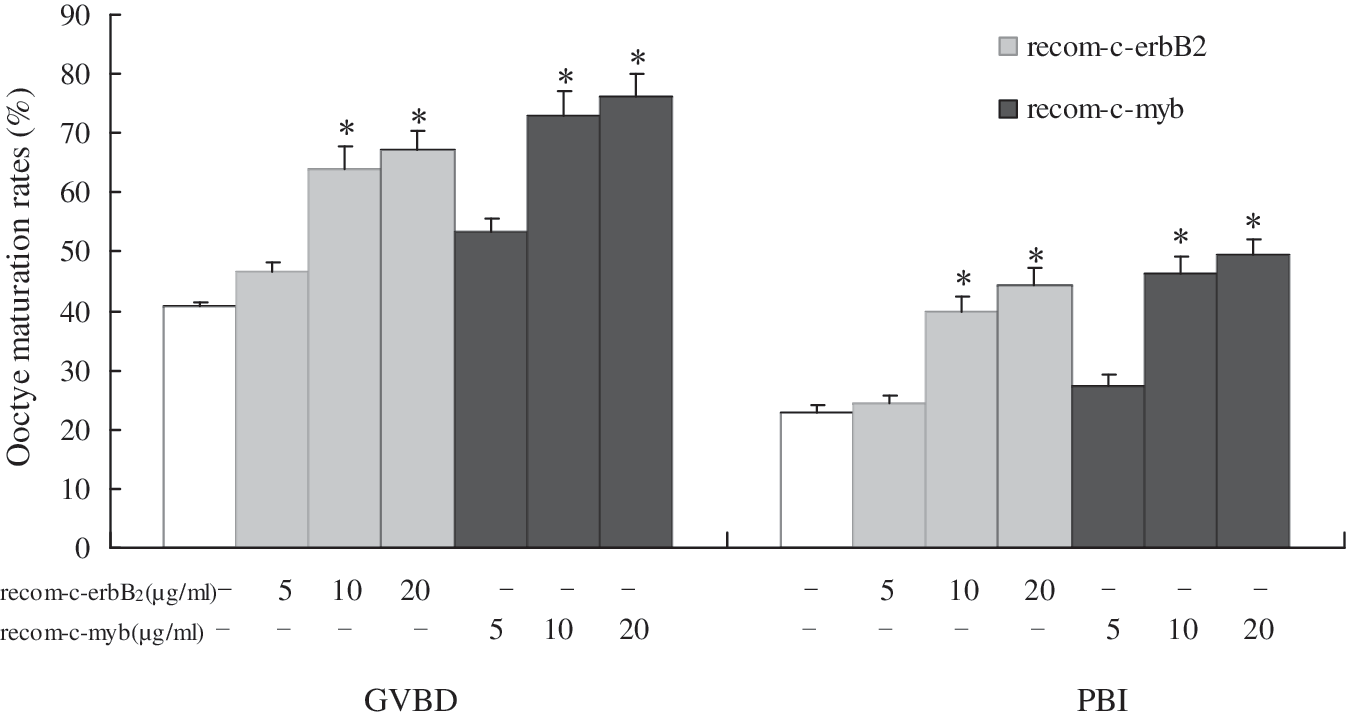
Effects of recombinant human c-erbB2 and c-myb protein on mouse oocyte maturation after microinjection into oocyte cytoplasm. rabbit IgG (10 μg/mL); c-erbB2 recombinant protein (5, 10, 20 μg/mL); c-myb recombinant protein (5, 10, 20 μg/mL). *p<0.05 compared with rabbit IgG group.
Effect of roscovitine on c-erbB2 and c-myb protein-induced oocyte maturation
To investigate the relationship between c-erbB2 , c-myb and MPF activity, oocytes were challenged by 50 μM roscovitine after microinjection of recombinant human c-erbB2 (10 μg/mL) or c-myb (10 μg/mL) protein to observe GVBD and PB1 extrusion. As shown in Figure 5, roscovitine blocked the promoting action of both c-erbB2 and c-myb recombinant proteins on oocyte maturation, and it reduced the rates of GVBD and PB1 extrusion to the control levels (recom-c-erbB2 : GVBD 37.5% vs 69.6%, PB1 29.2% vs 47.8%; recom-c-myb: GVBD 33.3% vs 78.9%, PB1 29.2% vs 57.9%, p<0.05 respectively). However, roscovitine had no effect on the expression of c-erbB2 mRNA and c-myb mRNA (Figs. 2 and 3).
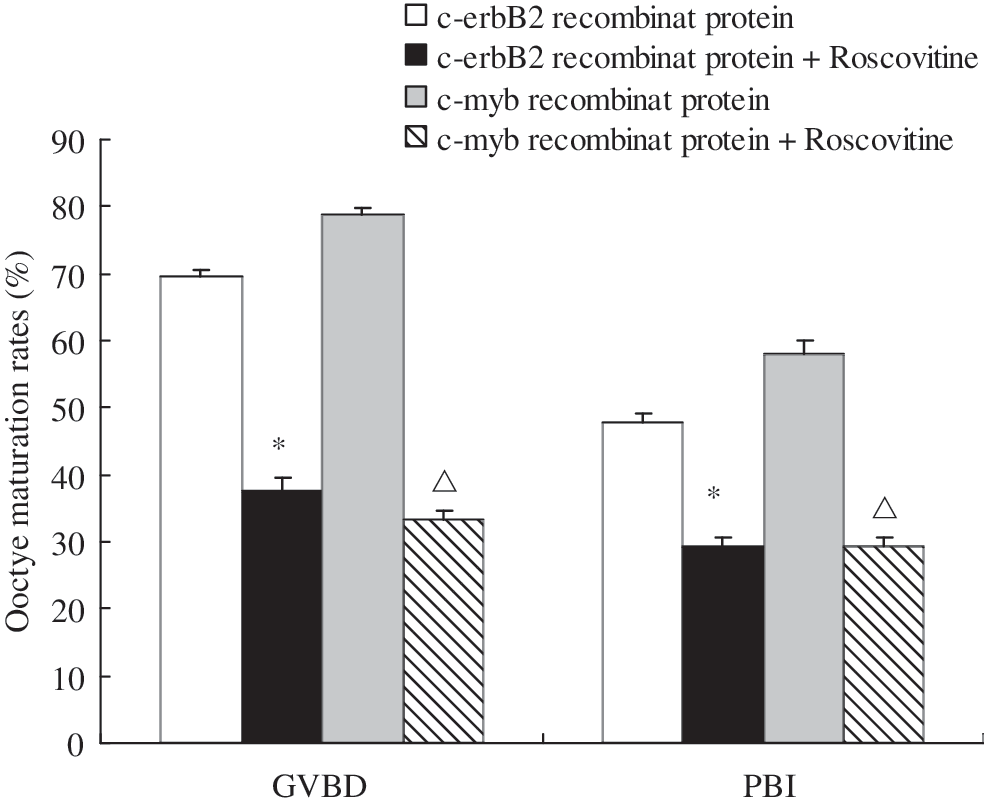
Roscovitine blocked recombinant human c-erbB2 protein and c-myb protein-induced mouse oocyte maturation. c-erbB2 recombinant protein (10 μg/mL); c-erbB2 recombinant protein (10 μg/mL)+roscovitine (50 μM); c-myb recombinant protein (10 μg/mL); c-myb recombinant protein (10 μg/mL)+roscovitine (50 μM). *p<0.05 compared with c-erbB2 recombinant protein group. Δ p<0.05 compared with c-myb recombinant protein group.
Involvement of cyclin B1 in c-erbB2 and c-myb-induced oocyte maturation
Western blot analysis revealed that the expression of cyclin B1 protein was inhibited by c-erbB2 ASODN, c-myb ASODN, and roscovitine, compared with the control (Fig. 6).
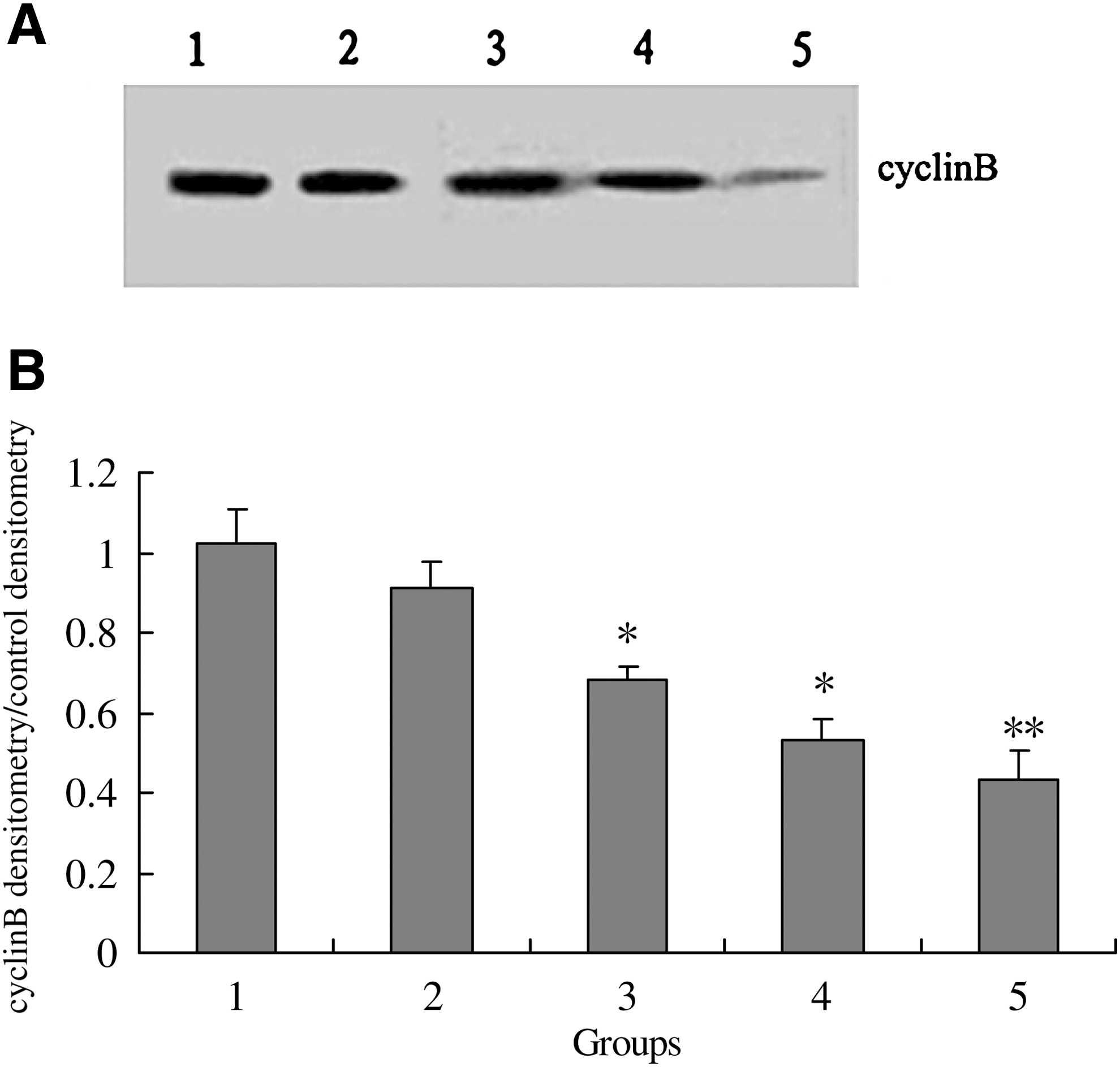
Effects of c-erbB2
ASODN, c-myb ASODN, and roscovitine on cyclin B1 expression of mouse oocytes after 8 h culture.
Discussion
Much effort is currently focused on investigating the functions of a large number of genes involved in female reproduction. In the present study, we used antisense oligodeoxyribonucleotides and the microinjection of recombinant proto-oncogene protein to study the functions of c-erbB2 and c-myb in the mouse oocyte maturation.
The protein product of c-erbB2 proto-oncogene belongs to the erythroblastosis oncogene B (ErbB) receptor family, which is also called the epidermal growth factor (EGF) receptor family. Binding of EGF to the extracellular region of EGF receptor induces receptor dimerization and activates the cytoplasmic kinase, which leads to autophosphorylation and initiation of downstream signaling pathways. Several reports indicated that EGF enhanced oocyte maturation in a dose- and time-dependent manner (Moor et al., 1998; Pang and Ge, 2002). Therefore, we hypothesized that c-erbB2 may be involved in oocyte maturation. The c-myb proto-oncogene encodes a sequence-specific DNA-binding protein (Myb) that induces transcriptional activation or repression. Some proteins that had long-term effects on cellular structure and function were generated, and cellular function was influenced by Myb (Klempnauer et al., 1982; Calabretta and Nicolaides, 1992; Sano and Ishii, 2001; Hess et al., 2006; Malaterre et al., 2007). In the current experiment, when expression of c-erbB2 or c-myb mRNA in oocytes was arrested by antisense oligodeoxyribonucleotides, the oocyte meiotic maturation was significantly inhibited. On the other hand, oocyte meiotic maturation was promoted when the ErbB2 and Myb proteins were increased by microinjection of recombinant human c-erbB2 protein and c-myb protein. These results indicated that the two proto-oncogenes play an essential role in oocyte maturation.
MPF plays an important role during oocyte meiotic maturation. MPF is a ubiquitously expressed serine/threonine protein kinase. Cdc2 has been identified as the catalytic subunit of MPF, whereas cyclin B acts as the regulatory subunit. The binding of these two subunits is critical to the transition into M-phase of the mammalian cells. MPF regulates the maturation of oocytes, and activated MPF phosphorylates cellular machinery components involved in nuclear envelope breakdown, chromosome condensation, spindle assembly, and cyclin degradation through activating many proteins such as histone H1, RNA ploymerase II, and pp60c-src kinase (Hampl and Eppig, 1995; Heikinheimo et al., 1998).The regulators of the catalytic subunit cdc2 have been well studied, and CPEB-Mos-MAPK-90-kD ribosomal protein S6 Kinase (p90rsk)-cytostatic factor C is one important pathway to regulate cdc2 activity (Palmer et al., 1998; Lu et al., 2002; Fan and Sun, 2004; Liang et al., 2005; Madgwics et al., 2006; Racki and Richter, 2006). However, the upstream activator of the regulatory subunit, cyclin B of MPF, is more complex. Unlike mitotic cells, GV stage mouse oocytes have only a limited pool of cyclin Bl, and successful completion of the first meiotic cycle requires intensive cyclin Bl synthesis throughout M-phase (Polanski et al., 1998). The amount of cyclin Bl during meiotic maturation is regulated by synthesis and degradation of this protein (Ledan et al., 2001; Hyslop et al., 2004).The reports in starfish and mouse suggested the presence of an unknown GV-associated factor(s) acting via 3′UTR-dependent control of cyclin Bl synthesis (Lapasset et al., 2005; Hoffmann et al., 2006).Two early mitotic inhibitors 1 (Emil) and 2 (Emi2) were also reported to control cyclin B content (Madgwics et al., 2006; Marangos et al., 2007).
To further explore the molecular mechanism of c-erbB2 and c-myb action during oocyte meiotic maturation, we attempted to investigate MPF as a possible mediator with roscovitine, an active MPF inhibitor. Oocyte meiotic maturation induced by recombinant human c-erbB2 protein or c-myb protein was blocked by roscovitine, but not by rabbit IgG, an inactive control, indicating that MPF is involved in c-erbB2 and c-myb induced oocyte maturation. In addition, western blot analysis demonstrated that the expression of cyclin B1 was inhibited by c-erbB2 ASODN and c-myb ASODN, but the expression of c-erbB2 mRNA and c-myb mRNA in oocytes was not influenced by treatment with roscovitine. These results suggested that proto-oncogenes c-erbB2 and c-myb might the upstream activators of MPF in regulating oocyte maturation through increasing the synthesis of cyclin Bl.
In conclusion, both c-erbB2 ASODN and c-myb ASODN inhibited oocyte meiotic maturation. Microinjection of recombinant c-erbB2 or c-myb protein promoted oocyte maturation, whereas the promoting effect was blocked by a MPF inhibitor. In addition, the expression of cyclin B1 was inhibited by c-erbB2 ASODN and c-myb ASODN. Therefore, we presume that c-erbB2 and c-myb may be upstream regulators of MPF and induce oocyte maturation via activation of MPF.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 30260035) and Jiangxi province major sciences and technologies supporting program. The authors thank Zhi-Sheng Zhong of the Temple University of Fels Institute for his kind suggestions. The authors declare that there is no conflict of interest that would prejudice the impartiality of this scientific work.
Disclosure Statement
No competing financial interests exist.