
Abstract
Prosaposin (PASP) is a sphingolipid hydrolysis protein that plays roles in both the nervous and reproduction systems. In this study, we cloned the pig PASP gene and studied its genomic organization, polymorphism, and expression pattern. Two PASP transcripts, TV1 (HQ245644) and TV2 (HQ245646), were identified in pig. TV1 was the complete transcript that encoded 527 amino acids, whereas TV2 was 9 bp shorter due to an exon 8 deletion. The pig PASP gene spanned over 34 kb in length on chromosome 14 (SSC14), and consisted of 15 exons and 14 introns. The pig PASP gene (TV1 and TV2) expressed predominantly in the cerebellum, lymphnode, pituitary, abdominal fat, hypothalamus, and cerebrum in both females and males. PASP TV1 expressed mainly in teh cerebrum, cerebellum, hypothalamus, pituitary, heart, subcutaneous fat, and foreleg muscle, while TV2 was expressed in the liver, spleen, lung, kidney, and lymphnode. In foreleg muscle, the predominant transcript was TV2 in males and TV1 in females. Some potential transcriptional elements were predicted in 5′ flanking region (∼3000 bp) of the PASP gene, and they were TATA boxes, RORE, Sp1, SRY, oct-1, Cdx A, and cap. Additionally, we identified 68 single-nucleotide polymorphisms and 9 indels in the pig PASP gene, and three single-nucleotide polymorphisms (C77932320T or L15F; C77928094T or P191L; A77917401G or K522R) were nonsynonymous substitutions. These results provide useful information for future functional investigations of the pig PASP gene.
Introduction
The male reproductive organ in PASP null (−/−) mice had reduced size and weight of the testes, epididymides, seminal vesicles, and prostates, and reduced spermiogenesis, and an involution of the prostate, seminal vesicles, and epididymis (Morales et al., 2000a). In the prostate of PASP inactivated animals, the secreted cells disappeared and only the basal epithelial cells were present; moreover, the prostatic epithelium proliferation and differentiation was reduced (Morales et al., 2000b). These findings suggested that PASP is an important gene for reproduction traits.
Located on chromosome 10q21–22, the human PASP gene consists of 15 exons and 14 introns, and encodes saposin A (exon 3–5), saposin B (exon 6–8), saposin C (exon 10–11), and saposin D (exon 12–14) (Zhao and Morales, 2000). Different PASP isoforms were identified in human (Nakano et al., 1989; Holtschmidt et al., 1991a, 1991b; Lamontagne and Potier, 1994), mice (Zhao et al., 1998; Cohen et al., 2005), rats (Hiraiwa et al., 2003), chickens, as well as zebrafish (Cohen et al., 2004). Two major isoforms of PASP differed by 9 bp (CAGGATCAG) in exon 8, which encoded three amino acid residues (Gln-Asp-Gln) of the saposin B domain (Lamontagne and Potier, 1994). Differences in expression levels were also found. Therefore, alternative splicing is an important factor to consider when studying the neurotrophic effects of PASP (Hiraiwa et al., 2003; Cohen et al., 2004). It was further reported that the longer transcript with the 9 bp insertion was preferentially secreted from cells, whereas the shorter one was mainly trafficked to the lysosome (Madar-Shapiro et al., 1999). Little is known about the pig PASP gene. In the present study, we obtained cDNA and transcript variants, and investigated the expression and polymorphisms of the pig PASP gene.
Materials and Methods
Animals
Two small-ear spotted pigs (one female and one male) at 120 days of age were slaughtered to collect 22 tissues: cerebrum, cerebellum, hypothalamus, pituitary, heart, foreleg muscle, back leg muscle, back muscle, spleen, lymphnode, testicle, ovary, oviduct, uterus, lung, kidney, liver, stomach, large intestine, small intestine, subcutaneous fat, and abdominal fat. Ovary tissue was used for cloning the pig PASP gene, whereas all tissues were used for real-time quantitative polymerase chain reaction (qPCR). A total of 25 genomic samples at 120 days of age were used for identification of polymorphisms across the complete coding region of the pig PASP gene. These samples included small-ear spotted pigs (5 individuals), Landrace (5), Yorkshire (5), Lantang pigs (5), and Duroc (5). Small-ear spotted pigs and Lantang pigs are both Chinese pig breeds, whereas Landrace, Yorkshire, and Duroc are all imported commercial pig breeds. These samples were provided by either the Institute of Animal Science, Guangdong Academy of Agricultural Sciences (Guangzhou, China), or Prof. Yonggang Liu (Yunnan Agricultural University, China).The animal care and use protocol was approved by the Laboratorial Animal Care and Use Committee in Jinan University, China, in accordance with Law of the People's Republic of China on Animal Protection.
Primers
P1 was designed according to the predicted PASP cDNA sequences (GenBank accession number: XM_001928494) to clone the pig PASP gene, and P2 was used for real-time qPCR analysis of PASP gene. Primer P3 was used for qPCR analysis of pig β-actin gene (DQ452569) as the internal control. P4 was designed to specifically show the length polymorphism (9 bp indel) of two PASP transcript variants (TV1 and TV2) by reverse transcription (RT)-PCR. Based on obtained sequences and the pig genome database, P5–P13 were designed and used to identify polymorphisms across the coding regions of the pig PASP gene. All primers were designed using GeneTool Lite software (
F and R indicate forward and reverse primer. E and In refer to exon and intron.
RT, reverse transcription; PCR, polymerase chain reaction.
RNA extraction and DNA preparation
Total RNA was extracted from fresh or frozen tissues using Trizol reagent (Invitrogen, Carlsbad, CA). A mixture of 2 μg total RNA, 1 μg oligo dT primer, and 20 nmol deoxynucleotide triphosphates was used for cDNA synthesis using M-MLV Reverse Transcriptase (Promega, Madison, WI). Genomic DNA was extracted from pig ear samples with the phenol-chloroform method (Sambrook et al., 1989).
Cloning of the PASP gene cDNA by RT-PCR
RT-PCR was performed to amplify the cDNA of the PASP gene using primer P1 or P4. The 25 μL reaction mixtures contained 1 μL cDNA, 1.25 U ExTaq polymerase (Takara, Osaka, Japan), 12.5 pmol primers, and 200 μM dNTP (each). PCR was run in a Mastercycler gradient (Eppendorf Limited, Hamburg, Germany) with three steps: 94°C predenaturing for 3 min, followed by 35 cycles of 94°C denaturing for 30 s, 58°C or 60°C annealing for 45 s, and 72°C extension for 1 min, as well as a final extension of 5 min at 72°C. PCR products were checked by 1.0% agarose gel electrophoresis and purified with a QIAquick gel-extraction kit (Qiagen, Valencia, CA). The purified PCR products were then cloned into pGEM-T Easy Vector plasmid (Takara) and sequenced by a commercial service (Biosune Co. Ltd, Shanghai, China). The obtained sequences were assembled to obtain the full-length cDNA of the pig PASP gene.
Genomic structure and variation analysis of the pig PASP gene
The obtained cDNA by this study was used for BLAST search (
Quantitative real-time PCR analysis of PASP mRNA expression in pig tissues
The pig β-actin gene was used as an internal control to quantify PASP mRNA by qPCR. The 20 μL mixture contained 1 μL cDNA template, 10 μM primers, and 2× Q-PCR SYBR Green Mix (Toyobo, Japan). Real-time PCR was performed at 95°C for 3 min, followed by 40 cycles of 30 s at 95°C, 30 s at 63°C, and 40 s at 72°C in an ABI7500 thermocycler (Applied Biosystems, Foster City, CA). To analyze PCR specificity, melting curve analysis was performed. In addition, the qPCR products were verified by sequencing (Biosune Co. Ltd).
In this study, each data point was repeated three times. Quantitative values were obtained from the threshold PCR cycle number (Ct) at which the increase in signal associated with an exponential growth for PCR product starts to be detected. The relative mRNA levels in each sample were normalized by β-actin. The relative expression levels of the pig PASP gene was indicated by 2−ΔCT, for which ΔCT = Cttarget gene - Ctß-actin.
Semiquantitative RT-PCR assay of TV1 and TV2 mRNA expression in adult pig tissues
P4 was used to amplify parts of TV1 and TV2 by RT-PCR, giving rise to fragments of 106 bp (TV1) and 97 bp (TV2) each. The mixture and PCR reaction program was described above. Then, the amplified fragments were separated by nondenatured polyacrylamide gel (8%, w/v) in 90 mM Tris buffer pH 8.0, 90 mM borate, and 2 mM EDTA. DNA was stained with silver nitrate. The relative expression of these two transcripts was investigated in different tissues: cerebrum, cerebellum, hypothalamus, pituitary, heart, liver, spleen, lung, kidney, abdominal fat, subcutaneous fat, foreleg muscle, back leg muscle, back muscle, small intestine, large intestine, stomach, lymphnode, testicle, ovary, oviduct, and uterus.
Bioinformatic analysis and prediction of transcription factors
The protein and CD data sets of PASP in various species were downloaded from NCBI database, and they were human (protein ID of NP_035309, CDs ID of NM_011179), mouse (NP_001035930; NM_001042465), rat (NP_001177165; NM_001190236), horse (XP_001503814; XM_00153764), dog (XP_861590; XM_856497), chicken (AAF05899; AF108656), frog (NP_571968; NM_131883), and zebrafish (NP_571968; NM_131883). Identity percentages (%) were calculated with DNASTAR software (
Results
The full-length cDNA of the pig PASP gene
Two transcripts of the pig PASP gene, the longer transcript (PASP TV1) and the shorter one (PASP TV2), were obtained in this study. They are available in the NCBI database with accession numbers of HQ245644 and HQ245645, respectively. The obtained PASP TV1 was 1660 bp in length containing an open reading frame of 1584 bp and encoded a protein of 527 amino acids (AA) (Fig. 1). The PASP TV2 was 9 bp shorter than TV1 due to a 9 bp deletion, and therefore it encoded a shorter protein (deletion of 3 AA) (Fig. 1). The pig PASP encoded by TV1 showed high homology to human (85.0%), mouse (62.9%), rat (62.5%), and chicken (58.0%) (Fig. 2).

cDNA and deduced amino acid sequences of the pig PASP gene. Capital and italic letters show amino acids for each codon upside, whereas “*” refers to a stop codon. The boxed 9 nucleotides (CAGGATCAG, which encodes 3 amino acids, QDQ) indicate the region in exon 8 of the pig PASP gene which is present in TV1 but absent in TV2. PASP, prosaposin.

Alignment of the pig PASP with those of human, mouse, rat, and chicken. The four saposin domains are indicated by A–D.
Genomic organization of the pig PASP gene
BLAT search revealed that the pig PASP gene spanned over 34 kb at chromosome 14 (reverse strand) and consisted of 15 exons (9 to 201 bp) and 14 introns (101 to 19 368 bp) (Table 2). Exon 8 was the smallest exon, and the largest intron (19 368 bp) was intron 1. All introns identified in this study followed the GT-AG rule (Table 2). Further analysis of pig PASP showed that Saposin A, B, C, and D was encoded by exons 3–5, 6–9, 10–11, and 12–14, respectively (Fig. 3). Each saposin molecule did not start or end exactly at the beginning or end of a certain exon, due to the partial proteolytic processing of these proteins. The translation start codon (ATG) was located in exon 1 and the translation stop codon was in exon 15. Comparison of TV1 and TV2 showed that the presence or absent of exon 8 (CAGGATCAG) led to the formation of two PASP transcripts (Fig. 3).
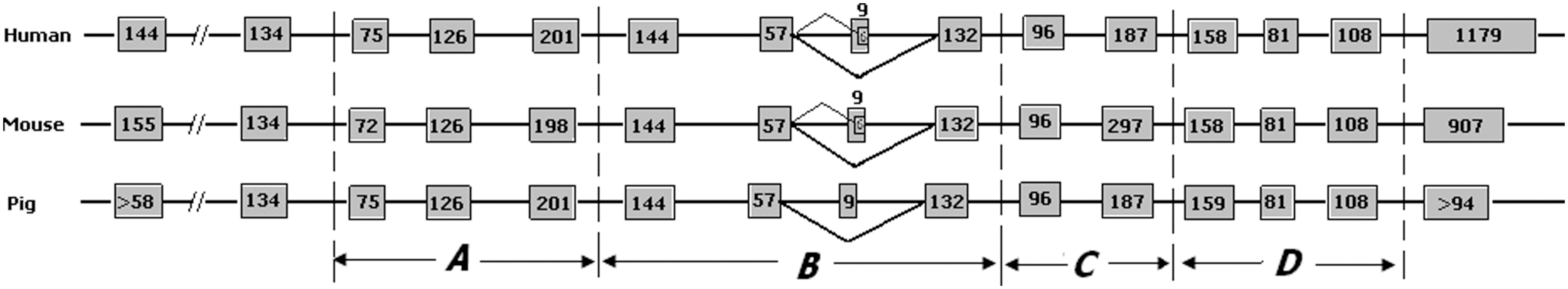
The genomic organization of the PASP gene among human, mouse, and pig. The four saposin domains are indicated by A–D. Exons are indicated by boxes with nucleotide size indicated inside except for exon 8, which varies between the TV1 and TV2 isoforms. “-//-” indicates a genomic gap existed there according to BLAT search (
The capital and bold letters indicate the boundary nucleotides of exons, and lowercase letters indicate the boundary nucleotides of introns.
PASP mRNA expression in different tissues
qPCR showed that the total PASP transcripts (TV1 plus TV2) were found in most tissues in both females and males (Fig. 4). In females, it expressed predominantly in subcutaneous fat, and then in cerebellum, hypothalamus, cerebrum, lymphnode, pituitary, liver, abdominal fat and spleen (Fig. 4). In males, it expressed predominantly in lymphnode and cerebellum, and then in back muscle, pituitary, and abdominal fat (Fig. 4).

Total PASP mRNA (TV1 plus TV2) in different pig tissues. Cer, cerebrum; Ceb, cerebellum; Hyp, hypothalamus; Pit, pituitary; Hea, Heart; Liv, liver; Spl, spleen; Lun, lung; Kid, kidney; Abf, abdominal fat; Suf, subcutaneous fat; Flm, foreleg muscle; Blm, back leg muscle; Bam, back muscle; Smi, small intestine; Lai, large intestine; Sto, stomach; Lym, lymphnode; Tes, testicle; Ova, ovary; Ovi, oviduct; Ute, uterus. NS indicates nondetectable.
Differential expression of PASP TV1 and TV2 transcripts in pig tissues
Semiquantitative RT-PCR analysis showed that although TV1 and TV2 expressed in most pig tissues in both males and females, differential expression of the two transcripts was noted (Fig. 5). PASP TV1 was abundantly expressed in cerebrum, cerebellum, hypothalamus, pituitary, heart, subcutaneous fat, and foreleg muscle, but higher PASP TV2 expression was detected in liver, spleen, lung, kidney, and lymphnode. In males and females, we also found expression differences, such as in foreleg muscle: the predominant transcript in male was TV2, but it was TV1 in female. It was interesting that only TV2 was found in small intestine of females and foreleg muscle of males (Fig. 5).
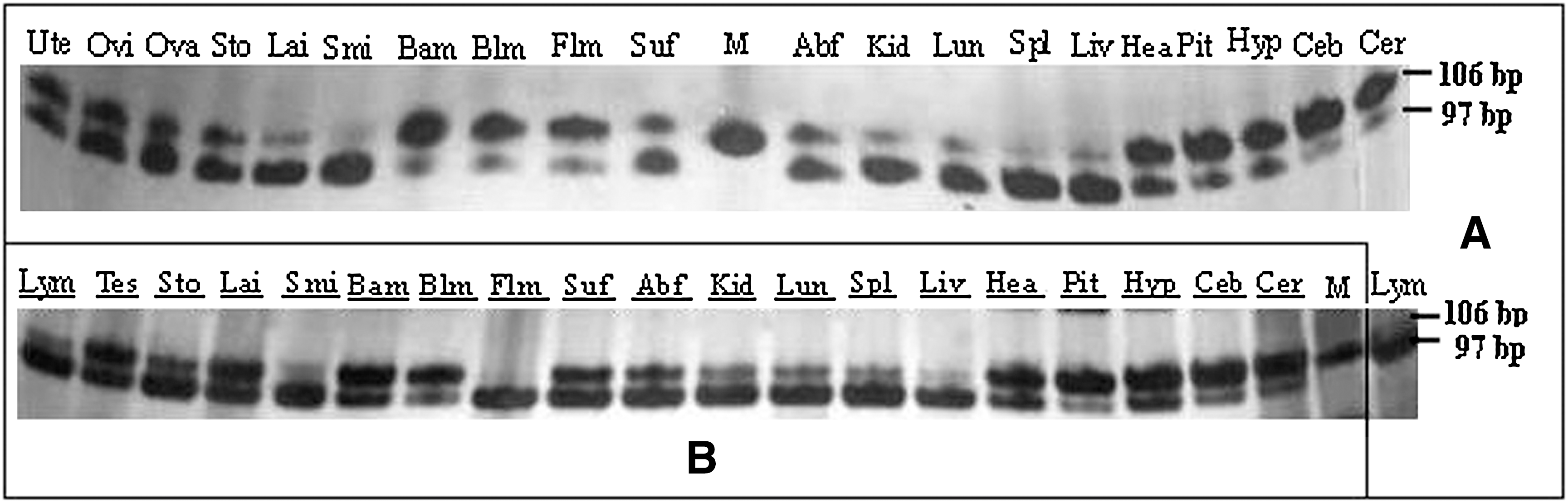
Detection of the differential expression of PASP TV1 and TV2 transcripts in male
Online prediction on binding sites
Online motif prediction showed that the 3000 bp fragment (−3000 to +1, nt) 5′ of the pig PASP gene contains binding elements for transcription factors (Fig. 6). The TATA box was located at −322 bp from the translation start site. We also found putative binding elements for RORE [retinoic acid-receptor-related orphan receptor a subunit (RORa)-binding element], Sp1 (stimulating protein 1), SRY (sex-determining region Y gene product), and other positive and negative regulatory elements (oct-1, Cdx A, and cap) (Fig. 6).

Putative binding sites for transcription factors. RORE, retinoic acid-receptor-related orphan receptor a subunit (RORa)-binding element; Sp1, stimulating protein 1; SRY, sex-determining region Y gene product; Ap-1, activator protein 1; Oct-1, octamer factor 1; Cap, cap signal for transcription initiation; CdxA, CdxA.
Single-nucleotide polymorphisms of the pig PASP gene
A total of 68 single-nucleotide polymorphisms (SNPs) and 9 indels were identified in exons and introns of the pig PASP gene (Table 3). Among 17 SNPs located at the coding region, three led to substitution of amino acids, and they were C77932320T (CTC→TTC, L15F), C77928094T (CCG→CTG, P191L), and A77917401G (AAG→AGG, K522R). One SNP was located at the 3′ UTR region of the pig PASP gene. Moreover, 62 variations were located at recognize sites for restriction enzymes, and therefore could be genotyped by PCR-RFLP.
The site of each SNP refers to the nucleotides in chromosome 14, according to Nov. 2009 (SGSC Sscrofa9.2/susScr2) pig BLAT search (
SNP, single-nucleotide polymorphisms.
Discussion
The results of cDNA cloning, expression, and homology analysis by this study proved the genomic and structural conservation of PASP among mammals. The encoded pig PASP precursor (by TV1) shared identities of 85%, 62.9%, and 62.5% with its counterparts of human, mouse, and rat, respectively. The pig PASP protein also comprised four functional domains of saposins A, B, C, and D, which are necessary for sphingolipids hydrolysis (Fig. 2). Like human and mouse, the pig PASP gene also consists of 15 exons and 14 introns, and all introns followed the GT-AG rule (Fig. 3). In addition, two isoforms of pig PASP (TV1 and TV2) were found in this study (Fig. 1). Alternative splicing for TV1 and TV2 occurred in exon 8 (CAGGATCAG), encoding for three amino acids (Gln-Asp-Gln) of the saposin B domain. These two isoforms were also reported in human (Nakano et al., 1989; Holtschmidt et al., 1991a, 1991b; Lamontagne and Potier, 1994), mice (Zhao et al., 1998; Cohen et al., 2005), rats (Hiraiwa et al., 2003), chickens, and zebrafish (Cohen et al., 2004) (Fig. 3).
The high expression of pig PASP in reproductive and nervous tissues suggested that it functions in these systems. The pig PASP gene was found to abundantly express in nerve center (cerebellum, cerebrum, hypothalamus, and pituitary), fat tissues (subcutaneous fat and abdominal fat), immune system tissues (spleen and lymphnode), and organs of reproduction (ovary and oviduct). In mouse, the PASP gene also highly expressed in the nervous and reproductive systems in both adult and embryonic stage animals (Sprecher-Levy et al., 1993). This feature was probably related to the reported important roles of PASP in reproductive system (Sprecher-Levy et al., 1993), nervous system (O'Brien et al., 1994), and during sphingolipid catabolism (O'Brien and Kishimoto, 1991; Kishimoto et al., 1992). However, in the cerebellum, cerebrum, hypothalamus, spleen, lymphnode, subcutaneous fat, and back muscle, PASP mRNA levels were different between male and female pigs; whether this sex bias was related to a different internal environment between male and female remained unclear.
TV1 and TV2 expressed differently in various tissues. PASP TV1 abundantly expressed in cerebrum, cerebellum, hypothalamus, pituitary, heart, subcutaneous fat, and foreleg muscle, but a hig level of PASP TV2 mRNA was detected in the liver, spleen, lung, kidney, and lymphnode. In adult rats, it was reported that a PASP transcript equivalent to TV1 preferentially expressed in brain, heart, and muscle, whereas a 9 bp shorter PASP transcript (equivalent to TV2) was preferentially expressed in lung, liver, and kidney tissues (Hiraiwa et al., 2003). In mouse and chicken brain, expression of PASP with 9 bp increased during development (Cohen et al., 2004). In normal brain, the ratio of two transcripts (TV1/TV2) was 85:15; however, it decreased to 5:95 after ischemic injury (Hiraiwa et al., 2003). These findings indicated that the two PASP isoforms play important roles in neuron regeneration (Chen et al., 2008). Moreover, PASP isoforms were reported to change binding specificity of saposin B presumably to adapt to the variable sphingolipid composition of tissues (Lamontagne and Potier, 1994). Another study, however, reported that mice with an exon 8 deletion were healthy and fertile, and no changes were detected in PASP secretion or in accumulation and metabolism of gangliosides, sulfatides, or neutral glycosphingolipids (Cohen et al., 2004).
Online prediction of potential motifs was useful for investigation of regulatory elements of the pig PASP gene. In 5′ flanking region (3000 bp) of the pig PASP gene, we predicted binding sites for RORE, Sp1, SRY, and other positive and negative regulatory elements (oct-1, Cdx A, and cap) (Fig. 6). We also found a TATA box at −322 bp. Studies in vitro indicated that the mouse PASP promoter lost its TATA box but had positive and negative regulatory elements at −2400 bp (Sun et al., 1997). An important region at −310 bp might modulate the expression of PASP in mice (Jin et al., 1998). In transgenic mice created by deletion of the 5′-flanking region of PASP, it was reported that the modifiers (ROR alpha- and Sp1-binding sites) residing within 310 bp of the 5′-flanking region mediated the spatio-temporal and differential expression of PASP in the central nervous system and eye (Sun et al., 2000, 2003). As in other species, the identified SNPs of the PASP gene have potential effects on pig production traits. In humans, the deletion of valine (11Val) in saposin A protein caused the abnormal galactosylceramide metabolism in infants (Spiegel et al., 2005), and the N215K substitution in saposin B led to the loss of the single N-glycosylation site in a patient with metachromatic leukodystrophy (Regis et al., 1999). A single bp deletion (c.803delG) in the saposin B domain led to a complete deficiency of PASP and saposins, and was associated with a complex sphingolipidosis dominated by lactosylceramide accumulation (Hulková et al., 2001). Saposin D (−/−) mice generated by C509S mutation developed progressive polyuria at around 2 months and ataxia at around 4 months (Matsuda et al., 2004). Other variations of this gene were related to Gaucher disease (Horowitz and Zimran, 1994). All these SNPs identified by this study provided candidate markers for further functional investigation of the pig PASP gene.
Conclusion
We identified two transcripts (TV1 and TV2) of the pig PASP gene and found that they expressed differently among various tissues and between males and females. The genomic organization of the pig PASP gene was described, and a total of 68 SNPs found by this study.
Footnotes
Acknowledgments
This study was supported by the National Major Special Projects on New Varieties Cultivation for Transgenic Organisms (2008ZX08006–005 and 2009ZX08009–145B) and the Important Projects in Key Fields in Guangdong and Hongkong, 2008 (2008A02).
Disclosure Statement
No competing financial interests exist.