
Abstract
Mitogen/extracellular signal-regulated kinase kinase-5 (MEK5), which belongs to a network of mitogen-activated protein kinase pathways, play a pivotal role in carcinogenesis. The purpose of this study was to investigate whether variants in the MEK5 gene promoter were involved in susceptivity of individuals to sporadic colorectal cancer (CRC). In the present hospital-based case–control study of 737 patients with sporadic CRC and 703 healthy control subjects in a southern Chinese population, the two polymorphisms of MEK5 promoter (i.e., rs7172582C>T and rs3743354T>C) were genotyped by TaqMan assay. There were significant differences between cases and controls in the genotype and allele distribution of the MEK5 gene rs3743354T>C polymorphism. The rs3743354 CC genotype was associated with a significantly decreased risk of CRC when compared with the TT genotype (adjusted odds ratios [ORs]=0.43; 95% confidence interval [CI], 0.24–0.77). Compared to the T allele, a significant correlation was detected between the presence of the C allele and decreased risk of CRC (adjusted OR=0.79; 95% CI, 0.61–0.94). The decreased risk of CRC associated with rs3743354 variant genotypes (i.e., CT+CC) was found in the smoker subgroup (adjusted OR=0.63; 95% CI=0.45–0.88). Further, environmental factors, including smoking and drinking, interacted with rs3743354C variant genotypes to reduce CRC risk. Western blot analysis showed that the levels of MEK5 protein in sporadic CRC neoplastic tissues and adjacent normal colorectal epithelium tissues were lower in the carriers of rs3743354 CC genotypes than that in those with rs3743354 TT genotypes or those with rs3743354 TC genotypes. However, no significant association was found between the rs7172582C>T polymorphism and risk of CRC. These data indicate that the rs3743354 polymorphism in the MEK5 promoter may affect the risk of developing CRC.
Introduction
Mitogen/extracellular signal-regulated kinase kinase-5 (MEK5), also known as MAP2K5, belongs to a network of MAPK pathways. MEK5 in turn specifically phosphorylates and activates extracellular signal-regulated kinase-5 (ERK5) (Zhou et al., 1995), which directly phosphorylates and activates several transcription factors including c-Myc, Sap-1, c-Fos, Fra-1, and myocyte enhancer factor family members (Kato et al., 1997; English et al., 1998), eliciting a variety of biological responses to extracellular signals that include cytokines, growth factors, and various stress stimuli (Roberts et al., 2009). As the only kinase upstream of ERK5, MEK5 has two splice variants MEK5α and MEK5β. The expression of the MEK5β protein is greater than that of MEK5α in terminally differentiated tissues, whereas MEK5α expression is greater in mitotically active tissues such as the liver. MEK5α directly stimulates ERK5 activity, whereas MEK5β plays a kinase-dead dominant-negative role that suppresses ERK5 signaling (Seyfried et al., 2005). A growing number of studies showed that overexpression of MEK5α is associated with tumorgenesis and malignancies (Cameron et al., 2004; Song et al., 2004; Ghosh et al., 2005) and the expression ratio of MEK5α to MEK5β is higher in cancer cell lines, whereas overexpression of MEK5β inhibits serum-induced DNA synthesis (Cameron et al., 2004). Therefore, alternative splicing of MEK5α and MEK5β may play a pivotal role in ERK5 activation and subsequent carcinogenesis. Further, it is thought that MEK5 is directly involved in the progress of malignancies and tumorigenesis (Song et al., 2004). There are many studies suggesting that MEK5 plays a critical role in cancer occurrence and progression, such as prostate cancer (Mehta et al., 2003; McCracken et al., 2008), breast cancer (Li et al., 2008), hepatocellular cancer (Yan et al., 2009), and lung cancer (Winn et al., 2006). However, the association between MEK5 and CRC remains unclear.
Single-nucleotide polymorphisms (SNPs, i.e., small genetic changes of single-base nucleotides in DNA) vary among individuals, and this variance is a susceptivity in cancer (Lo et al., 2007; Li et al., 2009), particularly when these SNPs are located within regulating regions (Pan et al., 2010). Genetic variations in the promoter region may affect its functions. In addition, some environmental factors can affect gene activity by acting on the promoter region. We previously demonstrated that the polymorphism rs3826392 (-1304T>G) in the promoter of the mitogen-activated protein kinase kinase 4 (MKK4) gene, which also belongs to a network of MAPK pathways, is associated with decreased risk of sporadic CRC. Further, the gene–environmental interaction analysis showed that the age and alcohol drinking status interacted with rs3826392 G variant genotypes to reduce cancer risk (Wei et al., 2009). Several SNPs in the promoter of the MEK5 gene have been described. We investigated whether the genetic variations within the promoter region of the MEK5 gene, and their possible interactions with environmental factors, play a role in the risk of sporadic CRC.
The aims of this case–control study were to clarify the impact of two common MEK5 SNPs in the promoter region on CRC risk in a Chinese population and the gene–environmental interactions between potential confounders and to investigate these SNPs. Moreover, we analyzed the correlation between a significant MEK5 promoter region SNP and protein expression levels in sporadic CRC neoplastic and adjacent normal colorectal epithelium tissues.
Materials and Methods
Study subjects and sample collection
This study population included 737 sporadic CRC patients and 703 cancer-free controls. All enrolled subjects were unrelated Chinese individuals and were from Guangzhou City and surrounding regions in South China. All patients were recruited between July 2002 and March 2009 at the Sixth Affiliated Hospital (Gastrointestinal and Anal Hospital) of Sun Yat-Sen University (Guangzhou, China), Sun Yat-Sen University Cancer Center (Guanghzhou, China), the First Affiliated Hospital of Sun Yat-Sen University (Guangzhou, China), the Affiliated Tumor Hospital of Guangzhou Medical College (Guangzhou, China), Guangdong Provincial People's Hospital (Guangzhou, China) and Panyu People's Hospital (Guangzhou, China). Cases suffering from hereditary nonpolyposis CRC, familial adenomatous polyposis, or inflammatory bowel disease and other related malignancies were excluded. All patients were untreated sporadic CRC cases and were histopathologically diagnosed and recruited with a response rate of about 95% in our study. Cancer-free controls were randomly selected from about 12,000 individuals who participated in the healthy checkup programs in the community health stations conducted in Guangzhou City during the same time period when the cases were recruited, with a response rate of about 85%. These control subjects were frequency-matched to the cases on age (65 years) and sex. Each participant first gave an informed consent and was scheduled for an interview, in which a structured questionnaire was administered by trained interviewers to collect data on smoking status, alcohol use, and other factors including sex, age, family history of cancer, and BMI. Smoking status, alcohol use, and family history of cancer were defined as previously described (Wei et al., 2009). The BMI was categorized by the cutoff points suggested by the Cooperative Meta-Analysis Group of Working Group on Obesity in China (Bei-Fan, 2002). Subjects whose BMI was ≤23.9 kg/m2 were categorized as being underweight or normal body weight, subjects whose BMI was from 24.0 to 27.9 kg/m2 were defined as being overweight, and those with BMI ≥28.0 kg/m2 were categorized as obese. Subjects who have any first- or second-degree relative (or both) with a history of cancer were defined as “Yes” and the remaining subjects as “No.” Each subject was asked to donate 5 mL of peripheral venous blood after giving informed consent. The study was approved by the institutional review boards of Sun Yat-Sen University.
SNP selection
Based on the GenBank dbSNP database (
Genotyping
Genomic DNA was extracted from peripheral blood using a QIAamp DNA Blood Mini Kit (Qiagen) according to the manufacturer's protocol. DNA purity and concentrations were determined by measurement of spectrophotometric absorbance at 260 and 280 nm. The corresponding primers and probes were designed using Primer Express 1.5 software (Applied Biosystems) and were purchased from Shanghai GeneCore BioTechnologies Co (Table S1). All samples were genotyped using an allelic discrimination assay on the Applied Biosystems 7500 FAST Real Time PCR instrument with TaqMan® probes. Real-time PCR was performed using 10 μL of TaqMan 2×universal master mix (Applied Biosystems), 2 μL of primer-probe (470 nM and 100 μM, respectively), 7 μL of DNase- and RNase-free water (Sigma), and 1 μL of sample DNA, in a total volume of 20 μL per single tube reaction. DNase-free water was used as a nontemplate control and DNA of the known MEK5 genotype (appropriate to the SNP of interest) was used as a positive control in each assay run. Assay conditions were as follows: 2 min at 50°C, 10 min at 95°C, and 55 cycles of 95°C for 15 s and 60°C for 1 min. First, the SNP assay was set up using sequence detection systems (SDS), version 2.1, software (Applied Biosystems) as an absolute quantification assay, and then the plate was read using the allelic discrimination settings. Eventually, postassay analysis was performed using the SDS software (Fig. S1). Ten percent of the samples were randomly repeated for quality control, and the concordance rate was 100%. The PCR products of every genotype were purified and confirmed by direct sequencing.
Western blot analysis
To analyze the correlation between observed significant SNP and MEK5 protein expression levels in vivo, fresh tissue samples of CRC and the adjacent normal colorectal epithelium were collected at the Sixth Affiliated Hospital of Sun Yat-Sen University. All tumor samples were histologically confirmed.
The frozen tissues were homogenized in T-PER tissue protein extraction (Thermo) lysis buffer, which contained a protease inhibitor. The mixture was centrifuged at 12,000 g for 15 min at 4°C and the supernatant was obtained. Sixty micrograms of a protein extract were separated by 10% SDS–polyacrylamide gel electrophoresis and transferred onto Polyvinylidene fluoride (PVDF) filters (Millipore Company). The membranes were blocked at 4°C overnight with blocking buffer (pH 7.6) containing 5% nonfat dry milk and incubated with rabbit anti-human monoclonal antibody against MEK5 (1:2000) purchased from BD Transduction Laboratories™. Immunoreactive proteins were stained using a chemiluminescence detection system. Membranes were then washed with stripping solution for 1 h and treated as described earlier but with mouse β-actin antibody (ProteinTech Group) as an internal control. The MEK5 protein expression levels were normalized to that of β-actin by calculating the relative expression levels.
Statistical analysis
Baseline sociodemographic characteristics between cases and controls were analyzed using the two-sided chi-square tests. The expected frequency of control genotypes was checked by the Hardy–Weinberg equilibrium test, which was tested by a goodness-of-fit chi-square test. LD between MEK5 polymorphisms was assessed by the 2LD program and the PROC ALLELE statistical procedure in SAS/Genetics (SAS Institute, Inc.) software. Odds ratio (OR) and 95% confidence intervals (CIs) were calculated to assess the relative risk conferred by a particular allele and genotype using an unconditional logistic regression model, with and without adjustment for age, sex, smoking status, alcohol drinking status, BMI, and family history of cancer. Logistic regression modeling was also used for the trend test. For the stratification analysis, we assessed the main effect of MEK5 in each subgroup and the interaction among the MEK5 polymorphisms and selected variables on cancer risk. A multiplicative interaction was indicated when OR11>OR10×OR01, in which OR11=the OR when both factors were present, OR01=the OR when only factor 1 was present, and OR10=the OR when only factor 2 was present. Interaction terms between variables were modeled using standard unconditional logistic regression to assess evidence for departure from a multiplicative model (Wang et al., 2005). All statistical analyses were two-sided and p<0.05 was considered statistically significant.
Results
Characteristics of the study population
The distribution of demographic characteristics of the 737 sporadic CRC cases and 703 matched controls is presented in Table 1. The proportion of current smokers and current drinkers in CRC cases was higher than that in the controls (current smokers: 38.0% vs. 30.6%; current drinkers: 41.8% vs. 18.9%), suggesting the matching was not adequate enough. Therefore, these variables were further adjusted for in the multivariate logistic regression model, and the adjusted variables were used in the later stratification and gene–environment interaction analysis.
MEK5 genotype distribution and risk of sporadic CRC
Both the SNPs (rs7172582 and rs3743354) were successfully genotyped. The genotype and allele distributions of the rs7172582 and rs3743354 polymorphisms between the cases and controls are presented in Table 2. The genotype distributions of the two SNPs among the participants in the control sample did not deviate from the Hardy–Weinberg equilibrium. The LD analyses showed that the linkage among the two SNP loci in controls were relatively weak (D′=0.532, γ 2=0.206), indicating every locus may have an independent effect on risk of sporadic CRC.
The observed genotype frequency among the control subjects was in agreement with the Hardy–Weinberg equilibrium (p 2+2pq+q 2=1) (χ 2=0.007, p=0.932 for rs7172582; χ 2=0.690, p=0.406 for rs3743354).
Two-sided χ 2 test for the distribution of either genotype or allele frequency.
ORs were adjusted in a logistic regression model that included age, sex, smoking status, drinking status, family history of cancer, and BMI.
CRC, colorectal cancer; CI, confidence interval; OR, odds ratio.
As revealed in Table 3, the frequency of the rs3743354 genotypes in the CRC patients (TT 64.5%, TC 32.8%, CC 2.7%) was significantly different from that in the control groups (TT 59.9%, TC 34.3%, CC 5.8%). Using the rs3743354 TT genotype as a reference, rs3743354 CC homozygotes have a significant 0.57-fold decreased risk of sporadic CRC (adjusted 95% CI, 0.24–0.77; p=0.004). Compared to the T allele, a significant correlation was detected between the presence of the C allele and a decreased risk of developing CRC (OR=0.79; 95% CI, 0.61–0.94; p=0.011). In addition, although our results for rs3743354 C variant genotypes (i.e., rs3743354 TC+CC) and the decreased risk of CRC did not reach statistical significance, the observed trend (adjusted OR=0.83; 95% CI, 0.66–1.03; p=0.095) indicates a somewhat protective effect. For the other SNP rs7172582, the genotype and allele frequencies did not significantly differ between the cases and the controls.
ORs were adjusted for age, sex, smoking status, alcohol use, family history of cancer and BMI in a logistic regression model.
p Value of test for the multiplicative interaction between MEK5 polymorphisms rs3743354T>C and selected variables on cancer risk were calculated using standard unconditional logistic regression models.
Stratification analysis of MEK5 genotypes and risk of sporadic CRC
A stratification analysis of the associations was conducted between the two SNPs' variant genotypes and the risk of sporadic CRC in subgroups by age, sex, smoking and drinking status, family history of cancer, and BMI. As shown in Table 3, a protected risk of the rs3743354 C variant genotypes associated with CRC achieved statistical significance in the subgroups of smokers (adjusted OR=0.63; 95% CI, 0.45–0.88; p=0.007). In the gene–environment interaction analysis, interactions between rs3743354 variant genotypes and smoking and drinking (p interaction=0.006 and 0.001; Bonferroni correction, p interaction=0.012 and 0.002) were observed, suggesting a possibility of more than multiplicative effects between rs3743354 C variant genotypes and smoking or alcohol drinking status. For the rs7172582 variant genotypes (i.e., rs7172582 CT+TT), however, no significant association with the risk of CRC was found in the stratification analysis.
Association of the rs3743354 polymorphism with MEK5 protein expression
To further analyze the association between rs3743354 polymorphism and the level of MEK5 protein expression, we collected 40 fresh tissue samples of CRC and the adjacent normal colorectal epithelium from the untreated sporadic CRC patients and found only one patient carrying the rs3743354 CC genotype. Therefore, we chose two patients carrying the rs3743354 TT genotype and two patients carrying the rs3743354 TC genotype randomly and the patient with the rs3743354 CC genotype and then detected the MEK5 protein levels of these tissue samples of these patients by western blot. We found that the levels of MEK5 protein were lower in the individual carrying the rs3743354 CC genotype than that in those with the rs3743354 TT or rs3743354 TC genotypes (Fig. 1).
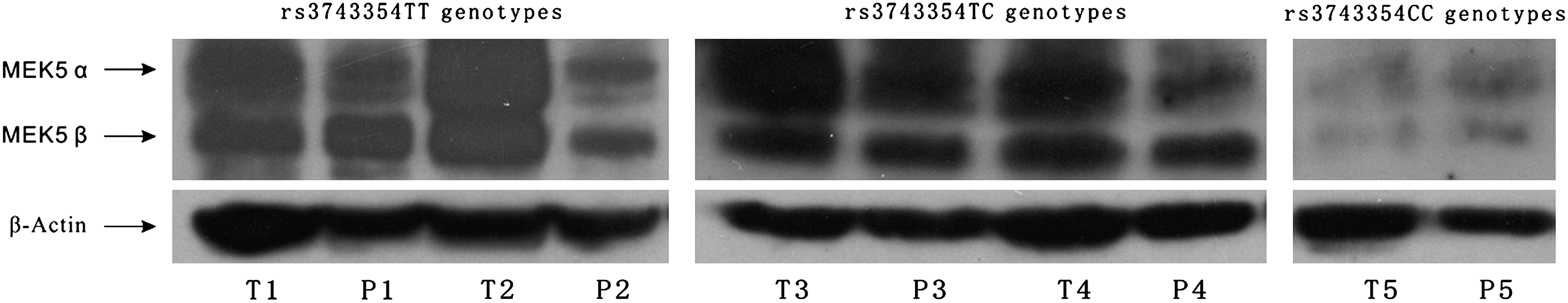
The association of rs3743354 polymorphism and mitogen/extracellular signal-regulated kinase kinase-5 (MEK5) protein expression. T, neoplastic tissue of colorectal cancer (CRC); P, adjacent normal colorectal epithelium tissue of CRC.
Discussion
To our knowledge, this is the first study to investigate whether MEK5 gene polymorphisms are related to the occurrence of sporadic CRC in a Chinese population. We found that the polymorphism rs3743354T>C in the MEK5 promoter is associated with decreased risk of CRC, particularly in the smoker subgroup. For the other SNP rs7172582C>T, no evidence was found for any association between this SNP and sporadic CRC risk. Further, we found that the MEK5 protein was downregulated in vivo in individuals who carried the rs3743354 CC allele, suggesting that rs3743354T→C substitution may decrease MEK5 protein expression in sporadic CRC tissues.
The signaling pathways that regulate cell proliferation, survival, and transformation, such as MAPK pathways, are of great importance in cancer biology. A number of studies demonstrated that the MAPK pathway was aberrantly activated in CRC (Shan et al., 2009). The MEK5/ERK5 pathway is sequentially regulated by a series of upstream MAP kinase kinases (MEKs) in a signaling cascade. Upon activation, the MAPKs selectively phosphorylate cellular targets, leading to regulation of gene expression and biologic events such as proliferation, differentiation, and apoptosis (Ghosh et al., 2005). There are reports that demonstrate that the MEK5/ERK5 pathway was involved in tumorgenesis and malignancies (Mehta et al., 2003; Dudderidge et al., 2007; Li et al., 2008). Dudderidge et al. (2007) showed that the induction of MEK5/ERK5 signaling is associated with activation of the DNA replication licensing pathway in prostate cancer. As the key upstream BMK1 kinase, MEK5 has two naturally occurring splice variants, MEK5α and MEK5β. Song et al. (2004) observed that MEK5α may be Stat3-regulated and play a critical role in oncogenesis mediated by aberrantly activated Stat3 signaling in breast carcinomatosis and malignancies. Ghosh et al. (2005) showed that exogenous expression of c-myc promoter-binding protein 1 induces prostate cancer cell death by downregulating the expression of MEK5α and upregulating the level of MEK5β. There are also some studies indicating that MEK5 plays an important role in the carcinogenesis of hepatocellular cancer (Yan et al., 2009) and lung cancer (Winn et al., 2006). However, few studies have investigated the role of MEK5 in sporadic CRC. Our finding reveals that the rs3743354 polymorphism in the MEK5 promoter region may be biologically important in the tumorigenesis of sporadic CRC.
We have previously argued that the polymorphism rs3826392 in the MKK4 gene promoter might modify the expression of the MKK4 protein and play a decrease role in risk of sporadic CRC, and the bioinformatics analysis showed that this may be due to altering the binding site of the transcriptional factor Nkx-2 (Wei et al., 2009). In this research, for the three SNPs, we also performed bioinformatics analysis with the TFSEARCH program (
CRC is a very complex disease and is unlikely to be significantly affected by a single genetic or environmental factor. Studies examining the associations between a single genetic variant and CRC risk may provide limited information, and statistical power is difficult to achieve. Therefore, approaches to assess the interactions of multiple genetic variants and gene–environment are very important. The stratification analysis showed that the decreased risk of CRC associated with the rs3743354 C variant genotypes was more pronounced in the subgroups of smokers and drinkers. Our data indicated that the protective effect of MEK5 rs3743354 genotype would mainly function under rigorous environmental and dietary influences. The interaction analysis showed that there were statistically significant interactions (or effect modifications) between MEK5 polymorphisms and smoking and drinking on cancer risk, indicating the gene–environment interaction in the etiology of sporadic CRC. This is coincident with the conclusion that occurrence of certain cancers is mediated by lots of relatively weak risk factors including genetic factors and environmental factors and the interactions among them (Ahmed, 2006; Torkamani et al., 2008).
Several potential limitations of the present study require consideration. First, our study focused on a Chinese population, and thus it is unclear whether our findings can be generalized to other populations. Second, in this hospital-based case–control study, all cases were recruited from six hospitals, whereas controls were selected from the communities, and thus selection bias may have occurred. Third, this study may have suffered from an information bias: although the questionnaires were completed before the diagnosis in our six hospitals, some patients referred from other institutions might have known their diagnosis. Another limitation is that the sample size may not be large enough, as the ratio of rs3743354 CC genotype is just 5.8% in healthy control and 2.7% in the CRC group. We achieved 83.2% study power (two-sided test, α=0.05) to detect an OR of 0.43 for the rs3743354 CC genotype compared with the rs3743354 TT genotype. Therefore, it appears that our finding that the rs3743354 CC genotypes associated with a decreased risk of sporadic CRC is unlikely to have been obtained by chance. Further, although we have recruited 40 fresh tissue samples of CRC and the adjacent normal colorectal epithelium, there was only one pair of tissues with rs3743354 CC genotype, and thus a larger study should be performed to conform this result.
In summary, the rs3743354T>C polymorphism was found to be associated with a moderate decreased risk of developing sporadic CRC in a population from southern China. Interactions between the rs3743354T>C polymorphism and smoking and drinking in the etiology of CRC were also found. However, further studies in different populations and with larger sample sizes, as well as well-designed mechanistic studies, are needed to confirm our finding.
Footnotes
Acknowledgments
This study was partly supported by the National Natural Scientific Foundation of China (Grant Nos. 81072042 and 30872488 to Dr. L. Wang), the Science Technology Development Program of Guangdong (Grant No. 2008B030301105 to Dr. J. Peng), and the Research Fund for the Doctoral Program of Higher Education of China (Grant No. 200805580074 to Dr. J. Wang). The authors thank Yang Jiao, for her assistance in recruiting the subjects, and Mr. Benjamin Greené, for revising the article.
Author Contributions
J. Wang and J. Peng designed the study; D. Diao and L. Wang performed all the experiments and wrote the paper; D. Chen, Y. Wei, and J. Lu partly performed population investigation, DNA extraction, and data management; J. Zhang performed the statistical analyses; and H. Liu revised the article.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.