
Abstract
It has been shown that DNA repair is reduced in patients with systemic lupus erythematosus (SLE) and that the X-ray repair cross-complementing (XRCC1) Arg399Gln (rs25487) polymorphism may contribute to DNA repair. We evaluated the frequency of the XRCC1 Arg399Gln substitution in patients with SLE (n=265) and controls (n=360) in a sample of the Polish population. The odds ratio (OR) for SLE patients with the Gln/Gln versus Gln/Arg or Arg/Arg genotypes was 1.553 (95% confidence interval [CI]=0.9573–2.520; p=0.0729). OR for the Gln/Gln or Gln/Arg versus Arg/Arg genotype was 1.551 (95% CI=1.122–2.144, p=0.0077). The OR for the 399 Gln allele in patients with SLE was 1.406 (95% CI=1.111–1.779, p=0.0045). There was also a statistically significant p-value of the χ 2 test for the trend observed in the XRCC1 Arg399Gln polymorphism (ptrend=0.0048). We also found a significant contribution of the Gln/Gln or Arg/Gln versus Arg/Arg genotype to the presence of either the malar rash or phototosensitivity manifestations of SLE OR=2.241 (1.328–3.781, p=0.0023, pcorr=0.0414). Moreover, the meta-analysis of Taiwanese Han Chinese, Brazilian, and Polish populations showed that the Gln/Gln or Gln/Arg genotype and Gln allele were associated with SLE incidence. OR for the Gln/Gln or Gln/Arg versus Arg/Arg genotype was 1.440 (95% CI=1.15–1.80, p=0.0019) and OR for the Gln allele was 1.27 (95% CI=1.08–1.51, p=0.0051). Our studies may confirm that the XRCC1 Arg399Gln polymorphism may increase the risk of incidence of SLE and the occurrence of some SLE manifestations.
Introduction
Genetic studies have demonstrated a strong contribution of many gene variants to SLE incidence and the clinical manifestations of SLE (Wong and Tsao, 2006; Hom et al., 2008; Flesher et al., 2010). Moreover, the induction of SLE development can be initiated by various environmental factors, drugs, chemicals, viral infections, and exposure to ultraviolet (UV) light (Mok and Lau, 2003; Sarzi-Puttini et al., 2005; Jönsen et al., 2007).
An accumulation of breaks in DNA combined with nuclear proteins may generate immunogenic antigens, which induce the production of auto-reactive antibodies (Abs) and an autoimmune response in susceptible individuals (Takeda and Dynan, 2001; Lee et al., 2002). Some studies have shown impaired DNA repair in patients with SLE, which suggests the involvement of various polymorphic variants of genes encoding the proteins responsible for DNA repair (Herrick et al., 1995; McCurdy et al., 1997).
In mammals, DNA is protected from ionizing radiation by proteins encoded by the X-ray repair cross-complementing (XRCC) gene family (Thacker and Zdzienicka, 2004). Some XRCC proteins have demonstrated their role in the incidence of various autoimmune disorders by their presence at DNA damage sites (Takeda and Dynan, 2001; Lee et al., 2002; Tajouri et al., 2003). XRCC1 functions as a platform protein to complex several enzymes responsible repairing DNA breaks (Tudek, 2007). Three single-nucleotide polymorphisms (SNPs) resulting in amino acids changes have been found at the XRCC1 gene (Hong et al., 2005). Recently, it has been reported that the XRCC1Arg399Gln (rs25487) polymorphism may play a role in an individual's susceptibility to SLE (Lin et al., 2009). The XRCC1 Arg399Gln minor allele exhibited relatively high frequency in the Polish population we studied for the distribution of Arg399Gln genotype and allelic frequencies in patients with SLE. As SLE is a heterogeneous disease, we also evaluated the association of these polymorphisms with varied SLE clinical symptoms in this group of patients.
Patients and Methods
Patients and controls
Two hundred sixty-five patients (women only) fulfilling the American College of Rheumatology Classification criteria for SLE (Tan et al., 1982; Hochberg, 1997) were selected randomly at Institute of Rheumatology in Warsaw, Poland (Table 1). The 360 controls consisted of randomly selected unrelated healthy female blood donors and healthy women who had a comprehensive examination at the Institute of Mother and Child in Warsaw, Poland. Both patients and control groups were of Polish Caucasian origin. The protocol of the study was approved by the Local Ethics Committee of Poznań University of Medical Sciences. Written agreement was obtained from all participating individuals. The mean age of SLE patients at diagnosis was 37±12 years, and of controls 36±11 years.
Significant results are highlighted in bold font.
Represents the absolute number of positive patients for Arg/Arg, Arg/Gln, and Gln/Gln genotypes or Gln MAF.
SLE was performed by χ 2 test.
χ 2 test for trend.
Genotype comparison (Gln/Gln vs. Arg/Gln or Arg/Arg) between patients with and patients without clinical manifestations of SLE.
Genotype comparison (Gln/Gln or Arg/Gln vs. Arg/Arg) between patients with and patients without clinical manifestations of SLE.
MAF, minor allelic frequency; SLE, systemic lupus erythematosus; CI, confidence interval; OR, odds ratio.
Genotyping
DNA was obtained from peripheral leukocytes using a salting out procedure. The presence of the XRCC1 Arg399Gln (rs25487) variant was identified by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). PCR was carried out using primer pair 5′ ACCTTGTGCTTTCTCTGTGTC 3′ and 5′ TAGTCTGCTGGCTCTGGGCT 3′. The PCR-amplified fragments 509 bp in length containing the XRCC1 Arg399Gln polymorphism were digested with endonuclease NciI (CC/SGG) (New England Biolabs, Ipswich, MA). The XRCC1 399 Arg allele was cleaved into 378 bp and 131 bp fragments, whereas the XRCC1 399 Gln allele was not cleaved (Fig. 1). The DNA fragments were separated according to size by electrophoresis on 2% agarose gel and observed by ethidium bromide staining. The XRCC1 Arg399Gln polymorphism was also confirmed by repeated PCR-RFLP analysis and commercial sequencing.
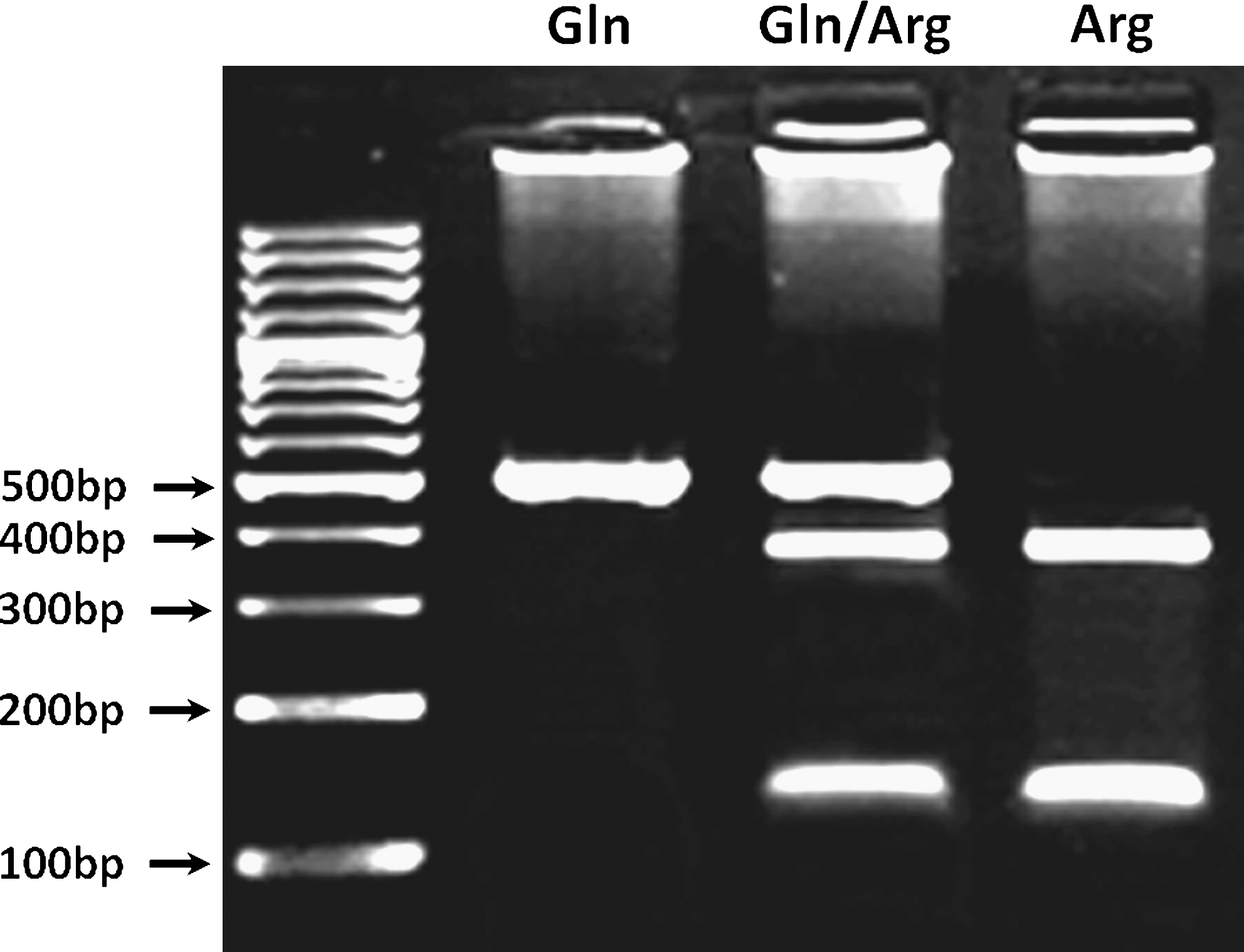
Representative picture of PCR–restriction fragment length polymorphism analysis of the XRCC1 Arg399Gln polymorphism. The 509 bp PCR-amplified fragments were digested with endonuclease NciI (CC/SGG). The XRCC1 399 Gln allele was not cleaved, whereas the XRCC1 399 Arg allele was cleaved into 378 bp and 131 bp fragments. The DNA fragments were separated by electrophoresis on 2% agarose gel and observed by ethidium bromide staining. PCR, polymerase chain reaction.
Statistical analysis
The distribution of genotypes in patients and controls was tested for deviation from Hardy–Weinberg equilibrium. Differences in genotypic and allelic distribution between patients and controls and associations between clinical manifestations, production of Abs, and polymorphism distribution in patients were determined by χ
2 or Fisher exact test. Bonferroni correction for multiple comparisons was used and both p-values, before (p) and after correction (p
corr), were determined. The XRCC1 Arg399Gln polymorphism was tested for association with SLE using the χ
2 test for trend (p
trend). The odds ratio (OR) and 95% confidence intervals (95% CI) were also determined. Statistical significance was interpreted as p-value <0.05. Power analysis was performed using χ
2 exact test, available at
Meta-analysis
The meta-analysis of three independent datasets (Bassi et al., 2008; Lin et al., 2009, and present study) was conducted using the Mantel-Haenszel fixed-effect model. The effect of allele contrast (Gln vs. Arg) and the contrast for the dominant (GlnGln/GlnArg vs. ArgArg) and recessive (GlnGln vs. GlnArg/ArgArg) models were estimated. The inter- and between-study heterogeneity was tested using the Breslow and Day method. The p-value for significance of association and heterogeneity tests was set to 0.10. All statistical calculations were conducted using the program EasyMA (Cucherat et al., 1997).
Results
Distribution of the XRCC1 Arg399Gln allelic and genotypic variant in SLE patients and controls
Genotype assessment of the XRCC1 Arg399Gln polymorphism did not exhibit significant aberration from the Hardy–Weinberg equilibrium in any group.
The frequency of the XRCC1 399 Gln/Gln genotype was approximately 1.5-fold times higher in SLE patients than in the controls, and reached 0.15 and 0.10 in those groups, respectively (Table 2). The prevalence of the XRCC1 399 Gln/Arg heterozygous genotype was also increased in individuals with SLE as compared to controls, and frequencies reached 0.48 and 0.42, respectively (Table 2). The OR for SLE patients with the Gln/Gln versus Gln/Arg or Arg/Arg genotypes was 1.553 (95% CI=0.9573–2.520; p=0.0729) (Table 2). OR for the Gln/Gln or Gln/Arg versus Arg/Arg genotype was 1.551 (95% CI=1.122–2.144, p=0.0077). We also found a significantly higher frequency of the XRCC1 399 Gln allele in patients than in controls, amounting to 0.39 and 0.31, respectively (Table 2). The OR for the 399 Gln allele in patients with SLE was 1.406 (95% CI=1.111–1.779, p=0.0045). The p-value of the χ 2 test for the trend observed for the XRCC1 Gln399Arg polymorphism was also statistically significant (ptrend=0.0048). The statistical power of this study was 76% for Gln/Gln Gln/Arg genotypes.
χ 2 analysis.
χ 2 test for trend.
(Gln/Gln vs. Arg/Gln and Arg/Arg).
(Gln/Gln and Arg/Gln vs. Arg/Arg).
(Gln vs. Arg).
Association of XRCC1 Arg399Gln genotypes with clinical symptoms of SLE
The most significant differences in the distribution of XRCC1 Gln/Gln or Arg/Gln versus Arg/Arg genotype was found in SLE patients with malar rash and phototosensitivity manifestations. These values amounted to OR=1.693 (95% CI=1.024–2.801, p=0.0395) and OR=1.700 (95% CI=1.029–2.810, p=0.0376) (Table 1), respectively. However, neither p-value was statistically significant after Bonferroni correction: p corr=0.711 and p corr=0.677, respectively. However, there was a significant contribution of the Gln/Gln or Arg/Gln versus Arg/Arg genotype to the presence of either a malar rash or phototosensitivity manifestations OR=2.241 (1.328–3.78, p=0.0023, p corr=0.0414) (Table 1). We did not find a significant association between the XRCC1 Arg399Gln polymorphism with other SLE clinical manifestations (Table 1). There was also no contribution of the XRCC1 Arg399Gln polymorphism to the presence of anti-dsDNA, anti-Smith, anti-snRNP, anti-Ro, anti-Scl-70, or anti-phospholipid Abs. We did not find significant differences in SLE disease activity index (Bombardier et al., 1992) at diagnosis between XRCC1 Arg399Gln genotype groups.
Meta-analysis of the XRCC1 Arg399Gln polymorphism
The meta-analysis was performed using data pooled from three studies conducted in Taiwanese Han Chinese (Lin et al., 2009), Brazilian (Bassi et al., 2008), and Polish populations. There was no heterogeneity between these studies in the analysis of XRCC1 Arg399Gln genotypes and the Gln allele. The p-value for the heterogeneity tests was >0.10. The OR for SLE patients with the Gln/Gln versus Gln/Arg or Arg/Arg genotypes was 1.17 (95% CI=0.84–1.64; p=0.39), (Table 3). OR for the Gln/Gln or Gln/Arg versus Arg/Arg genotype was 1.44 (95% CI=1.15–1.80, p=0.0019) (Table 3). The OR for the 399 Gln allele in patients with SLE was 1.27 (95% CI=1.08–1.51, p=0.0051).
Significant results are highlighted in bold font.
Mantel-Haenszel fixed-effect model.
Breslow and Day method.
Discussion
It has been reported that DNA repair is diminished in patients with SLE (Benke and Belmar, 1991; Bashir et al., 1993; McCurdy et al., 1997). Leucocytes from these patients exhibited increased levels of spontaneous single strand breaks and accumulation of oxidative DNA lesions as compared to those of healthy individuals (Benke and Belmar, 1991; Bashir et al., 1993). Moreover, SLE lymphocytes exposed to γ-radiation or SLE neutrophils with oxidative DNA damages may display reduced DNA repair ability (McCurdy et al., 1997; McConnell et al., 2002).
We found that the XRCC1 399Gln gene variant can be a risk factor for SLE incidence in a sample of the Polish population. Moreover, meta-analysis of Taiwanese Han Chinese (Lin et al., 2009), Brazilian (Bassi et al., 2008), and Polish populations showed that this gene variant can be associated with SLE incidence.
Recent studies conducted by Lin et al. (2009) in the Taiwanese Han Chinese population demonstrated a statistically significant higher frequency of the XRCC1 399 Arg/Gln heterozygote in patients with SLE than in controls. In contrast, Bassi et al. (2008) did not confirm the contribution of the XRCC1 Arg399Gln polymorphism to SLE development in Brazilian populations. However they observed the association of XRCC1 Gln/Gln or Arg/Gln genotypes with presence of anti-dsDNA Ab (Bassi et al., 2008).
In our study, patients with the XRCC1 Gln/Gln or Arg/Gln genotypes exhibited a significantly increased risk of developing either malar rash or phototosensitivity. An association between the XRCC1 Arg/Gln heterozygotes and phototosensitivity or malar rash was observed in SLE patients from the Taiwanese Han Chinese population (Lin et al., 2009). Furthermore, in this population, the XRCC1 Arg/Gln heterozygote has also been associated with hematologic and arthritis manifestations and presence of antinuclear Abs (Lin et al., 2009).
These differences in the effect of the XRCC1 Arg399Gln genotypes on the incidence of SLE and clinical manifestations in various ethnicities may be due to the racial heterogeneity or size of the studied groups. This distinction may also result from different populations' exposure to various environmental chemicals interacting with the XRCC1 Arg399Gln genotypes. This eventually may lead to different effects on the risk of SLE incidence in the studied ethnicities (Sarzi-Puttini et al., 2005; Jönsen et al., 2007).
To date, the XRCC1 399 Gln polymorphic variant has also been recognized as a genetic modifier for increasing the formation of DNA adducts and DNA damage in individuals exposed to cigarette smoke, aflatoxin B1, 1,3-butadiene, vinyl chloride metabolites, or styrene (Lunn et al., 1999; Abdel-Rahman and El-Zein 2000); Godderis et al., 2004; Li et al., 2009; Weng et al., 2009; Wang et al., 2010). Moreover, the XRCC1 399 Gln gene variant has been associated with lymphoblastic leukemia and carcinomas of head and neck, stomach, lung, prostate, colon, and breast (Sturgis et al., 1999; Abdel-Rahman et al., 2000; Divine et al., 2001; Duarte et al., 2005; Chen et al., 2006; Silva et al., 2007; Tumer et al., 2010). The XRCC1 399 Gln gene variant may also contribute to pterygium, schizophrenia, artherosclerotic coronary artery disease, and asthma incidence (Saadat et al., 2008; Chiang et al., 2010; Bazo et al., 2011).
XRCC1 protein is a crucial component of the base excision repair (BER) pathway for DNA damage. Although XRCC1 does not display enzymatic activity, it functions as a scaffold to coordinate the activity of the other proteins in the BER machinery, including glycosylases, an endonuclease (APE1), DNA polymerase β, a ligase, and Poly (ADP-ribose) Polymerases (PARP1 and PARP2) (Lindahl and Wood, 1999; Batar et al., 2010). The Arg399Gln polymorphism is situated in the central part of the XRCC1 BRCT1 domain, associated with the functioning of PARP1, PARP2, and APE1 (Vidal et al., 2001; Schreiber et al., 2002). Monaco et al. (2007) showed that the XRCC1 Arg399Gln substitution produces significant conformational alterations at several other sites in the BRCT1 domain. These alterations can be crucial in protein–protein interactions and may subsequently reduce the capability of the BER machinery formation. The decreased ability of the XRCC1 399 Gln protein in DNA repair has recently been demonstrated. Li et al. (2009) showed that immortalized lymphoblasts from vinyl chloride workers bearing the XRCC1 399 Arg/Arg genotype and exposed to etheno-adenosine adduct exhibited an average repair efficiency of 82%. In contrast, immortalized lymphoblasts from workers having the XRCC1 399 Gln/Gln genotype and exposed to the same adduct displayed an average repair efficiency of 32% (Li et al., 2009).
DNA damage can be produced by endogenous factors, including reactive oxygen species, or indirectly during BER, or exogenous factors such as chemicals and ionizing radiation (Thompson and West, 2000; Rastogi et al., 2010; Sutandyo, 2010). UV irradiation may damage DNA and is a powerful inducer of SLE in susceptible individuals (Pirner et al., 1992; Sarzi-Puttini et al., 2005). Impaired DNA repair may induce an autoimmune response in susceptible individuals with the XRCC1 399 Gln protein variant that has been exposed to DNA damage factors. This DNA damage may produce nucleoprotein complexes containing new composites or conformation epitopes or their cleavage products, which may present immunocryptic peptides that stimulate autoreactive T lymphocytes (Takeda and Dynan, 2001; Beyer et al., 2002; Lee et al., 2002; Mihara et al., 2002; van Bavel et al., 2010).
Our genetic investigation suggests the possible contribution of the XRCC1 399 Gln variant to SLE development and to either a malar rash or phototosensitivity manifestations. However, our genetic studies did not reach a power of 80%, due to a small sample size. Therefore, to reinforce the role of the XRCC1 399 Gln gene variant as a risk factor in SLE development and clinical manifestations of SLE, further studies of this polymorphism's distribution in other sample populations would be valuable.
Footnotes
Acknowledgments
This study was supported by grant no. 502-01-01124182-07474, Poznań University of Medical Sciences.
Disclosure Statement
No competing financial interests exist.