
Abstract
Gene expression analysis requires the use of reference genes consistently expressed under various conditions. In many cases, however, the commonly used reference genes are not uniformly expressed independently of tissues or environmental conditions. To provide a set of reliable reference genes in pigs, we used quantitative polymerase chain reaction to examine expression of six common reference genes (GAPDH, ACTB, H3F3A, HPRT1, RPL32, and RPS18) in adult tissues and prenatal skeletal muscles at 33, 65, and 90 days postcopulation from Tongcheng (obese-type) and Landrace (lean-type) pigs. The expression stability of these reference genes was evaluated by NormFinder, BestKeeper, and geNorm methods. Our data suggest that the reference genes were expressed variably in different tissues, developmental stages and breeds. RPS18, PRL32, and H3F3A could be used as internal controls to normalize gene expression in pig tissues and developmental skeletal muscle. The combination of internal control genes was necessary for accurate expression normalization. During skeletal muscle development, H3F3A and RPS18 would be the most appropriate combination to normalize gene expression in Tongcheng pigs, whereas the combination of PRL32 and RPS18 would be more suitable in Landrace pigs. In different tissues, the expression of PRL32 and RPS18 was the most consistent, and the combination of three genes (RPL32, RPS18, and H3F3A) is the most suitable for accurate normalization.
Introduction
The reference genes should be validated, ideally with consistent expression under various conditions. Some common reference genes, including ACTB, GAPDH, HPRT1 and RPS18, have been tested in northern blots, conventional reverse transcription (RT)-polymerase chain reaction (PCR) assays, and semiquantitative techniques for many years (Dheda et al., 2004; Huggett et al., 2005). These genes are supposed to have constant expression in different samples and experimental conditions. However, many studies prove that the expression of reference genes vary in different tissues, different cells, and different development stages (Thellin et al., 1999; Jain et al., 2006; Wan et al., 2010). Therefore, some work has been done to identify the most consistent reference genes by using quantitative PCR (qPCR) in humans (Thellin et al., 1999; Warrington et al., 2000), animals (Perez et al., 2008; Tatsumi et al., 2008), and plants (Jain et al., 2006; Hu et al., 2009). In some cases, since the use of a single gene as internal standards could lead to relatively errors, two or more reference genes are necessary for accurate normalization (Ohl et al., 2005; Jian et al., 2008).
The pig is an important agricultural animal and an ideal model organism. In our previous work, we have generated transcriptomic profiles of skeletal muscles from Tongcheng and Landrace pigs at 33, 65, and 90 days postcopulation (dpc) using long serial analysis of gene expression (LongSAGE). According to the LongSAGE analysis, we found that Histone 3 mRNA: H3 histone, family 3A (H3F3A) was consistently expressed in all LongSAGE libraries but the expression of GAPDH and ACTB was varied (Tang et al., 2007). After the LongSAGE analysis, we selected six commonly used reference genes (GAPDH, ACTB, H3F3A, HPRT1, RPL32, and RPS18) and evaluated them for gene normalization in pigs tissues and skeletal muscle development. Expression of mRNA encoding GAPDH, ACTB, H3F3A, HPRT1, RPL32, and RPS18 was examined in different tissues and skeletal muscles at 33, 65, and 90 dpc of Landrace and Tongcheng pigs by qPCR. The expression stability of these reference genes was evaluated by NormFinder (Andersen et al., 2004), BestKeeper (Pfaffl et al., 2004), and geNorm (Vandesompele et al., 2002) methods.
Materials and Methods
Sample collection and RNA extraction
Animal euthanasia was performed according to protocols approved by Hubei Province, PR China Animal Care and Use Committee for Biological Studies. Longissimus dorsi (LD) muscle samples were collected from Landrace and Tongcheng fetuses at the time points: 33, 65, and 90 dpc (three for each breed at each time point). Moreover, three adult pigs per breed were sacrificed for the tissues collection. Eleven different tissues (dorsal muscles, limb muscle, abdominal adipose, heart, liver, spleen, kidney, large intestine, small intestine, ovary, and oviduct) per Landrace pig were collected for RNA extraction, whereas ten different tissues (backfat, abdominal adipose, heart, liver, spleen, kidney, lung, small intestine, and ovary) per Tongcheng pig were collected for RNA extraction. All samples were immediately frozen in liquid nitrogen and stored at −80°C until use.
Total RNA was extracted using TRIZOL Reagent® (Invitrogen), as recommended by the manufacturer. The quantity and quality of RNA was measured by the Evolution 60 spectrophotometer (Thermo Scientific). Only RNA preparations with an A260/A280 ratio of 1.8–2.0 and an A260/A230 ratio >2.0 were used for subsequent analysis. We also checked the RNA integrity by analyzing the 28S/18S ribosomal RNA ratios on 1.5% agar gel electrophoresis.
cDNA synthesis
Before cDNA synthesis, RNA samples were treated with RNase-free DNase I (Promega) to prevent DNA contamination. The cDNA synthesis was carried out with 5 μg purified RNA in 20 μL final incubation volume using the Superscript III First-Strand Synthesis System (Invitrogen) for RT-PCR. The RT reaction was performed at 37°C for 5 min followed by 42°C for 1 h and 10 min at 70°C using a Mastercycler Gradient thermocycler (Eppendorf). The cDNA was diluted tenfold and stored at −20°C until use for the subsequent quantitative real-time PCR analysis.
Quantitative real-time PCR
qPCR was conducted using an iCycler iQ Multicolor Real-Time PCR Detection System with hydrolysis probe detection (BIO-RAD). Primer and probe sets were designed via the Applied Biosystems Primer Express™ 2.0 software. The gene accession numbers, primer and probe sequences, and amplicon lengths were shown in Supplementary Table S1 (Supplementary Data are available online at
Analysis of expression stability
qPCR data were exported into an Excel datasheet and analyzed using three separate reference gene stability analysis software programs: geNorm, BestKeeper, and NormFinder.
Results
Expression profiling of the candidate reference genes in Landrace and Tongcheng pigs
A real-time RT-PCR assay based on hydrolysis probe detection was used to measure the expression profiles of the six reference genes (GAPDH, ACTB, H3F3A, HPRT1, RPL32, and RPS18) in the LD muscle at distinct stages of embryonic development (33, 65, and 90 dpc) and adult tissues from Landrace and Tongcheng pigs. To minimize experimental error, three independent technical replicates were performed for each experiment. Cq values were used to quantify the expression levels of the candidate reference genes. A higher Cq value is indicative of a lower expression level. In all samples, the rank of the six reference genes at mRNA expression level was (from high to low) as follows: RPS18 (18.66), RPL32 (19.25), ACTB (20.73), H3F3A (26.24), HPRT1 (26.58), and GAPDH (28.15).
NomrFinder analysis
NomrFinder was used to rank the candidate reference genes according to their expression stability in a given sample set and given experimental design. The gene with the lowest stability value will be top ranked, suggesting the expression is most consistent. The expression stability of these reference genes was analyzed in the following five sample-sets: (I) all 27 type samples, including both fetal LD muscle and adult tissues from Landrace and Tongcheng pigs; (II) 11 tissue samples from adult Landrace pigs; (III) 10 tissue samples from adult Tongcheng pigs; (IV) the LD muscle samples of three embryonic periods (33, 65, and 90 dpc) from Landrace pigs; and (V) the LD muscle samples of three embryonic periods (33, 65, and 90 dpc) from Tongcheng pigs.
As seen in Table 1, the most consistently expressed gene for sample set I and set III was RPL32, whereas RPS18 had the most consistent gene expression in sample set II and IV. RPS18 and H3F3A showed the most stable expression in sample set V.
BestKeeper analysis
BestKeeper calculated variations (standard deviation [SD] [±Cq] and variance coefficient [%Cq]) for the six reference genes in the five sample-sets (Table 2), showing the overall stability in gene expression. The reference genes can be ordered from the most stably expressed to the least by their variation. Any studied gene with the SD higher than 1 can be considered inconsistent (Pfaffl et al., 2004). The inter-gene relations were assessed by pairwise correlation analysis. The coefficient of correlation (r) to the BestKeeper Index of each candidate gene was shown in Table 3.
GM (Ct), the geometric mean of Ct; SD (±Ct), Ct standard deviation; CV (%Ct), variance coefficient expressed as percentage of Ct level.
The most consistently expressed gene for sample set I was RPL32 with the lowest variation (SD=0.97) and the highest correlation (r=0.947). In sample set II, the lowest variation was seen for the gene RPS18 (SD=0.78). GAPDH, H3F3A ACTB, and HPRT1 were considered inconsistent, as the genes with SD value higher than 1.0. In sample set III, RPL32 had the lowest variation and the highest correlation. In sample set IV, ACTB, RPS18, and RPL32 showed similar consistency with SD values of 0.17, 0.18, and 0.19, respectively. However, ACTB showed weaker correlation (r=0.587) than RPS18 (r=0.967) and RPL32 (r=0.909). Therefore, RPS18 and RPL32 could be used as internal controls to normalize gene expression. In sample set V, GAPDH was the most variable gene, because the SD value was higher than 1.0. Both RPS18 and RPL32 had the most consistent expression (SD=0.07). However, RPL32 demonstrated the lowest correlation with BestKeeper Index (r=−0.147). Therefore, GAPDH and RPL32 were eliminated from the candidate list. RPS18 was the most suitable reference gene for normalization in this sample set.
GeNorm analysis
The expression consistency of the six reference genes was evaluated using the geNorm software program (version 3.5). The geNorm VBA applet for Microsoft Excel determines the most consistently expressed reference gene in a set of biological samples by calculating the gene expression stability measure (M) for a reference gene and the average pairwise variation (V) for that gene with all other candidate genes (Currie et al., 1991). Genes with lower M values have more consistent expression patterns. The data were shown in Figure 1. When all samples were analyzed as one data set, the M value of RPL32 and RPS18 was the lowest, whereas the M value of GAPDH was the highest. This indicates that RPL32 and RPS18 were the most consistently expressed genes and that GAPDH was the most variably expressed gene (Fig. 1A). In the adult tissue samples from Landrace pigs (Fig. 1B) or Tongcheng pigs (Fig. 1C), RPL32 and RPS18 also had the least variable expression level and the expression pattern of GAPDH remained the most variable. In the LD muscle samples at the 33, 65, and 90 dpc of Landrace pigs (Fig. 1D), the expression pattern of H3F3A and HPRT1 was much more consistent than the other four reference genes. In contrast, the expression of H3F3A and RPS18 was the least variable in the LD muscle at the 33, 65 and 90 dpc of Tongcheng pigs (Fig. 1E), whereas GAPDH showed the most variable expression. Further, when the LD muscle samples at each time point from both breeds were analyzed as one data set, the expression pattern of H3F3A and RPS18 remained the most stable (Fig. 1F). The geNorm analysis demonstrated that GAPDH was the most variably expressed gene in all of the sample sets. This suggests that GAPDH was not a suitable gene for normalization. In contrast, RPL32 and RPS18 are suitable as internal controls. When gene expression was analyzed in skeletal muscle development, H3F3A was the most suitable gene for normalization.
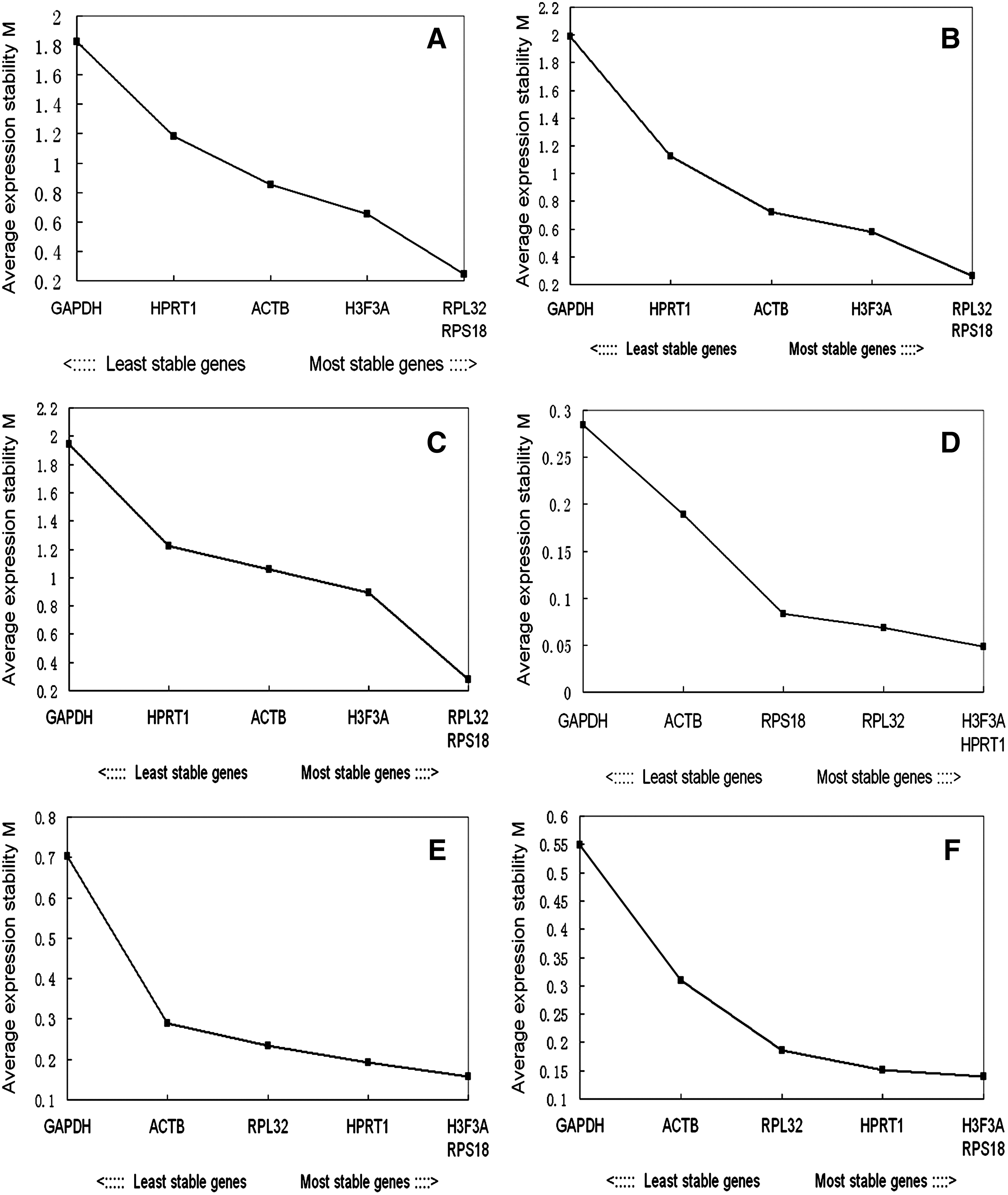
Average expression stability six reference genes calculated using geNorm. The expression stability and ranking of the six candidate reference genes in all 27 samples
A single reference gene alone could not serve as an appropriate internal control in the experiment; therefore, two or more reference genes are necessary for accurate normalization. geNorm determines the number of reference genes necessary for normalization by calculating the average pairwise variation (V) value. It proposes a cut-off value for the pairwise variation of 0.15, below which the inclusion of an additional reference gene is not required (Vandesompele et al., 2002). As shown in Figure 2A, when all samples were considered as a single data set, the pairwise variations V2/3, V3/4, V4/5, and V5/6 were all higher than the cut-off value of 0.15, indicating that the addition of a sixth reference gene for normalization is necessary. The situation was similar for the data sets composed of the different adult tissues from each breed (Fig. 2B, C). The expression of GAPDH, HPRT1, and ACTB; however, was very variable as shown in Figure 1A–C. Therefore, GAPDH, HPRT1, and ACTB could not be used as internal controls. The combination of RPL32, RPS18i, and H3F3A is suitable for accurate expression normalization in pig tissues. When the LD muscle samples from the three different embryonic stages of Landrace pigs were analyzed (Fig. 2D), the V2/3, V3/4, V4/5, and V5/6 were 0.025, 0.024, 0.069, and 0.078, all falling below 0.15. Similar results were seen with the LD muscle samples from the three embryonic periods of Tongcheng pigs (V2/3=0.064; Fig. 2E) and when the LD muscle samples at each embryonic period from both breeds were combined (V2/3=0.047; Fig. 2F). This indicated that there was no need to include a third reference gene in the normalization factor.
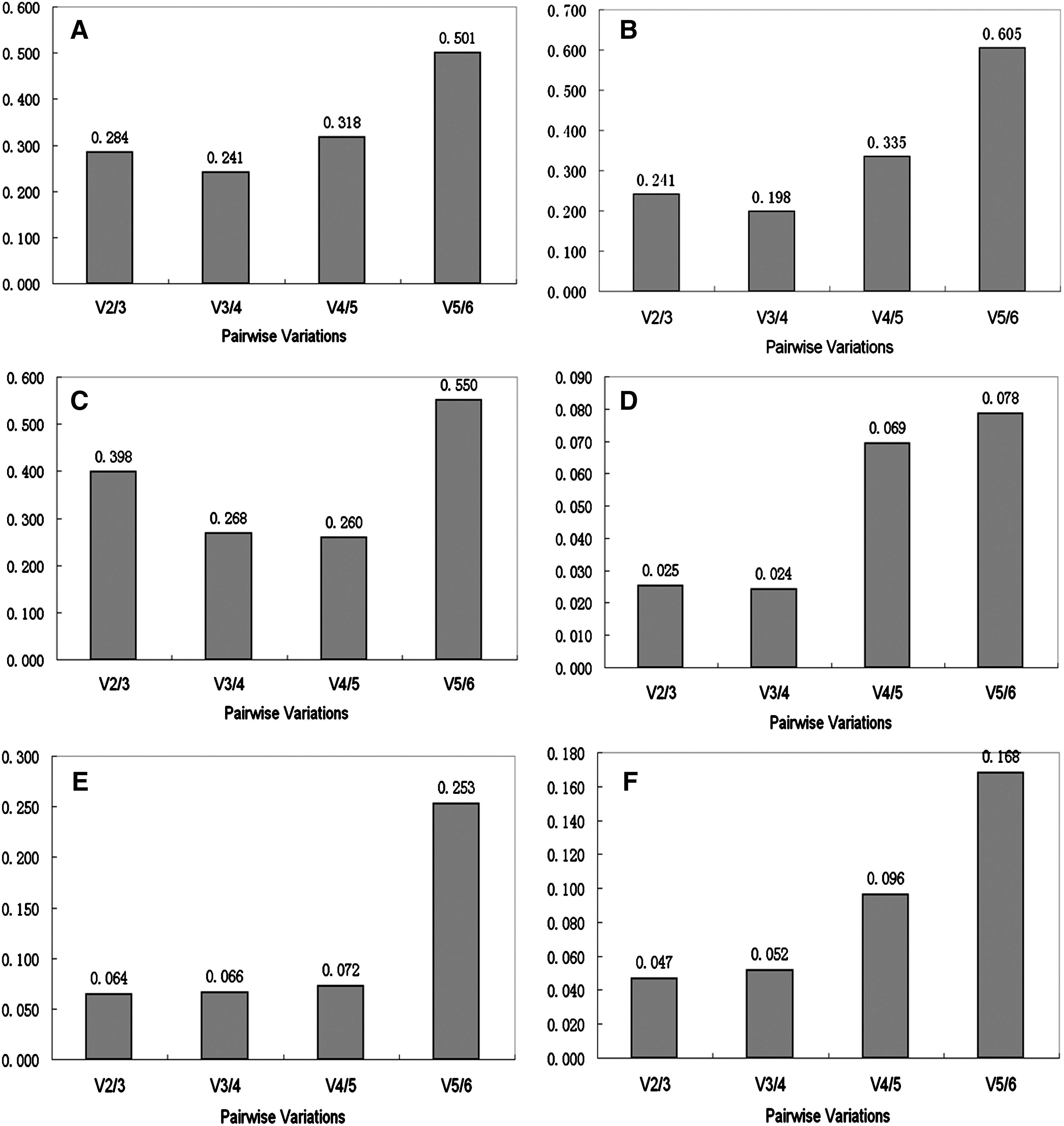
The optimal number of housekeeping genes for normalization as determined using pairwise variation. Pairwise variation to determine the optimal number of housekeeping genes for normalization in all 27 samples
Discussion
Gene expression analysis is the most common strategy applied to identify the genes relevant to new signaling and metabolic pathways that underlie many biological processes. It is critical to select appropriate reference gene for data normalization in gene expression analysis. Ideally, reference genes should show consistent expression levels in different samples and under different experimental conditions. However, evidence has shown that reference genes are variably expressed in different tissues, cell lines, and developmental stages. Therefore, it is essential to select a suitable reference gene for normalization. Some studies have been done in mice and human tissue using qPCR. Zhang et al. (2005) suggested that GNB2L1, HPRT1, RPL32, ACTB, and B2M may be suitable reference genes in gene expression studies of human neutrophils. Tatsumi et al. (2008) selected PPIA and TBP as the ideal reference genes for accurate normalization in mouse livers. In cattle studies, Perez et al. (2008) proposed SF3A1, EEF1A2, and HMBS as reference genes for normalization of gene expression studies in bovine muscle tissues. Goossens et al. (2005) recommend YWHAZ, GAPD, and SDHA as suitable internal controls for normalization of qPCR in bovine preimplantation embryos.
In molecular genetic studies of pigs, GAPDH and ACTB were the most frequently used reference genes for gene expression normalization. However, there is evidence that the expression of GAPDH is not consistent under various conditions (Glare et al., 2002; Erkens et al., 2006; Perez et al., 2008). Erkens et al. (2006) proposed ACTB, TBP, and TOP2B as reference genes for normalization of qPCR data in porcine backfat and LD muscle. Kuijk et al. (2007) reported that GAPDH, PGK1, S18, and UBC showed high stability in early porcine embryonic development. Foss et al. (1998) reported that the expression of GAPDH was more stable than ACTB in immune cells and tissues using northern blot and HPRT could be used as the endogenous control with qPCR. Nygard et al. (2007) investigated nine reference genes in different porcine tissues and reported that ACTB, RPL4, TPB, and HPRT1 were found to have the highest stability across tissues. To identify the more suitable reference genes for normalization in multiple tissues and skeletal muscle development of different porcine breeds, we investigated some other reference genes expression in 27 type samples, representing skeletal muscles at three embryonic period (33, 65, and 90 dpc), and various adult tissues of Landrace pigs and Tongcheng pigs. In agreement with the findings of Erkens et al. (2006) and Nygard et al. (2007), GAPDH was expressed unstably and not suitable as a reference gene in pig tissues. However, our results suggested that the use of ACTB as a reference gene would not be appropriate, as it showed unstable expression in our samples. In addition, RPL32 and RPS18 showed consistent expression among the studied samples and appeared more suitable for data normalization.
To evaluate these six reference genes for expression normalization, we used qPCR to examine mRNA expression of these genes and applied three methods (NormFinder, BestKeeper, and geNorm) to evaluate the expression consistency in the five sample-sets as described above. When using NormFinder, BestKeeper, and geNorm, it is necessary to pay attention to the foundation and prerequisites of the application: (1) high-quality total RNA; (2) the same amount of total RNA reverse transcribed under the same conditions; and (3) minimal experimental errors.
According to geNorm analysis, when all samples were considered together, RPL32 and RPS18 were the most consistently expressed genes. In the adult tissue series from each breed, RPL32 and RPS18 were also the most consistently expressed. These results were also supported by NormFinder and BestKeeper analysis. During the skeletal muscle development of Landrace pigs, geNorm analysis revealed that the least variably expressed genes were H3F3A and HPRT1. However, NormFinder and BestKeeper indentified that RPL32 and RPS18 were the most consistent genes. During the skeletal muscle development of Tongcheng pigs, geNorm and NormFinder results suggested that H3F3A and RPS18 had more consistent expression than the other four reference genes.
geNorm could also be used to determine the number of reference genes necessary for normalization by calculating the average pairwise variation (V) value. When the V value is below 0.15, the inclusion of an additional reference gene is not required. When analyzing the LD muscle samples at the three embryonic periods of Landrace pigs, the V2/3 was 0.025, indicating that a third reference gene was not necessary for normalizing gene expression. According to NormFinder and BestKeeper result, RPL32 and RPS18, instead of H3F3A and HPRT1, were considered as the best candidate reference genes to normalize the gene expression. When considering the LD muscle sample set of Tongcheng pigs and both breeds at the three embryonic periods, the V2/3 were below 0.15, showing H3F3A and RPS18 would be enough to normalize gene expression. When all samples were considered together and the various adult tissue samples of the two breeds were considered separately, the V2/3–V5/6 were all >0.15, indicating that a sixth reference gene was necessary for normalization. However, the GAPDH, HPRTI, and ACTB genes were not suitable as internal controls, due to their highly variable expression. We found that the V3/4 was the smallest V obtained, indicating that a fourth reference gene did not improve the accuracy of the normalization. Therefore, the combination of RPL32, RPS18, and H3F3A was most suitable for the normalization of gene expression data across all the samples and tissues of different breeds.
Conclusion
In conclusion, our study revealed that RPL32, RPS18, and H3F3A were expressed more consistently than the other three candidate genes across samples. In different tissues, the expression of PRL32 and RPS18 was the most consistent. The combination of RPL32, RPS18, and H3F3A is the most suitable for accurate expression normalization. During skeletal muscle development in Landrace pigs, RPL32 and RPS18 could be used to normalize gene expression, whereas H3F3A and RPS18 could be used as internal controls for Tongcheng pigs. These results will aid in the selection of reference genes in gene expression studies in pig tissues and skeletal muscle development.
Footnotes
Acknowledgments
We are grateful to Sanping Xu for help in collecting the animal material. This research was supported by the National Key Scientific Program (2009CB011600), the National Natural Science Foundation of China (30830080), the National Key Project (2008ZX08009-001), and the Project of Science and Technology Innovation Team “Research and Improvement of Domestic Animal Germplasm,” Institute of Animal Science, Chinese Academy of Agricultural Science (ywf-td-1).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.