
Abstract
DNA vaccines are known to be lacking in immunogenicity in humans. Presently, electroporation (EP) is thought to overcome this limitation. Here, we investigate whether human papillomavirus 16 E7 DNA vaccines delivered by EP might elicit potent antitumor activity in animal cervical cancer models, with a focus on the underlying mechanism(s). Intramuscular (IM)-EP delivery of E7 DNA vaccines induced more potent antitumor therapeutic and antimetastatic activity compared with IM delivery. Moreover, the tumor-controlled animals by IM-EP possessed long-term memory responses to parental tumor cells. This improved antitumor effect was concomitant with augmented Ag-specific CTL activities. IM-EP also induced IgG and Th-cell responses higher than IM delivery. Finally, IM-EP resulted in more antigen production in and more attraction of immune cells into the site of DNA injection, suggesting that these biological and immunological changes made by IM-EP might be responsible for enhanced CTL activities and antitumor resistance. Thus, this study shows that IM-EP can induce more potent antitumor activity by augmenting CTL responses possibly through more antigen production in and more attraction of immune cells into the muscle sites. This study also suggests that IM-EP of E7 DNA vaccines might be a potential approach toward treating patients with cervical cancer.
Introduction
Electroporation (EP), the administration of electrical pulses to muscle following DNA injection, has been shown to enhance the immunogenicity of DNA vaccines in a wide variety of small and large animal models (Han et al., 2010). The potential of EP delivery for clinical applications has been also suggested (Luxembourg et al., 2007; Bodles-Brakhop et al., 2009). DNA vaccines are less immunogenic in humans than in small animals (MacGregor et al., 1998; Wang et al., 1998; Barouch, 2006). The main reason for this might be resulting from a lower amount of plasmid relative to body mass or a lack of antigen-presenting cells (APCs) at the DNA injection site (Barouch et al., 2002). It has been reported that EP recruits dendritic cells and macrophages to the site of inoculation (McKay et al., 2004; Sumida et al., 2004; Liu et al., 2008). These localized effects were speculated to be associated with the strength and duration of the responses to vaccines in larger animals, including rabbits and humans (Ahlén et al., 2007). EP is also known to function by increasing myocyte permeability and thus facilitating plasmid uptake and antigen expression by host cells (Aihara and Miyazaki, 1998; Rizzuto et al., 1999; Widera et al., 2000; Gronevik et al., 2005).
In this study, we investigate whether E7 DNA vaccines delivered by EP might elicit more potent antitumor therapeutic and antimetastatic activity in an animal cervical cancer model, with a focus on the underlying mechanism(s). We observed that IM-EP of HPV 16 E7 DNA vaccines induces more potent antitumor therapeutic and antimetastatic activity, compared with IM delivery. This appeared to be mediated by augmenting Ag-specific CTL responses possibly through more antigen production in and more attraction of immune cells to the DNA injection sites. Further, IM-EP of E7 DNA vaccines displayed a significant degree of complete tumor regression in tumor-bearing mice. Thus, this study implies that IM-EP of E7 DNA vaccines might be a potential approach toward treating patients with cervical cancer.
Materials and Methods
Animals
Female C57BL/6 mice (5 weeks old) were purchased from Daehan Biolink (Chungbuk, Korea). Their care was performed under the guidelines of Institutional Animal Care and Use Committee–approved protocols.
DNA and immunization of mice
For IM delivery, the tibialis muscles of 6-week-old mice were injected at 1-week intervals with 50 μg of E7 DNA vaccines (pcDNA3-Sig/sE7/LAMP, pE7) (Kim and Sin, 2005) in a final volume of 50 μL of 0.25% bupivacaine-containing phosphate-buffered saline (PBS) using a 28-gauge needle (BD, Franklin Lakes, NJ). For IM-EP delivery, animals were injected IM as above with 50 μg of pE7 in a final volume of 50 μL of PBS, followed by EP at 0.2 A for 4 s using Cellectra® (equipped with three needle probes) from VGX International Inc. (Seoul, Korea). DNA vaccines were injected into the right and left leg muscles in sequence. pCR3.1-luciferase was kindly provided by D.B. Weiner (University of Pennsylvania School of Medicine). Plasmid DNA was produced in bacteria and purified by endotoxin-free Qiagen kits according to the manufacturer's protocol (Qiagen, Valencia, CA).
Tumor cell challenge and antitumor therapeutic study
For antitumor therapeutic studies between IM and IM-EP delivery methods of E7 DNA vaccines, 2×104 TC-1 cells were subcutaneously (s.c.) injected into the right mid-flank of C57BL/6 mice. For tumor rechallenge studies, 4×105 TC-1 cells per mouse were s.c. injected into the left flank of C57BL/6 mice and 4×105 control tumor cells (B16) per mouse were s.c. injected into the right flank of C57BL/6 mice. For tumor regression studies in tumor-bearing animals, 2×105 TC-1 cells per mouse were s.c. injected into the right mid-flank of C57BL/6 mice. When tumor sizes reached approximately 3–4 mm in diameter, the animals were given injections into the right and left leg muscles in sequence with E7 DNA vaccines by IM-EP. TC-1 tumor cells (a kind gift from T.-C. Wu, Johns Hopkins University, Baltimore, MD) were grown in cRPMI supplemented with 400 μg/mL of G418. The tumor cells were washed two times with PBS and injected into mice. Mice were monitored twice per week for tumor growth. Tumor growth was measured in millimeters using a caliper and was recorded as mean diameter [longest surface length (a) and width (b); (a+b)/2]. Mice were euthanized when tumor sizes reached more than 2 cm in mean diameter.
Antimetastatic effects
Animals were immunized twice at 2-week intervals with 50 μg of E7 DNA vaccines by IM and IM-EP delivery methods. At 4 weeks, the animals were intravenously (i.v.) challenged with 3×104 TC-1 cells per mouse. After 3 weeks following tumor cell challenge, animals were sacrificed to count metastatic nodules in the lung.
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) to detect Abs against recombinant E7 proteins (Kim et al., 2002) was performed as previously described (Kim et al., 2002; Ahn et al., 2003). In particular, recombinant E7 protein (1 μg/mL in PBS) was used as a coating antigen. To determine ELISA titers, equally pooled sera per group were twofold serially diluted from 1:50 and reacted with E7 protein. The titers were determined as the reverse of the highest sera dilution showing an optical density value similar to that of negative control.
Th-cell proliferative responses
Spleens were aseptically removed from each group and pooled together. Lymphocytes were harvested from spleens and prepared as the effector cells by removing the erythrocytes and by washing several times with cRPMI (RPMI 1640 supplemented with 10% fetal bovine serum, 1% penicillin/streptomycin, and 1%
IFN-γ assay
A 1 mL aliquot containing 6×106 splenocytes was added to wells of 24-well plates. Then, the cells were stimulated with 1 μg/mL E7 peptides (containing the MHC Class I epitope at amino acids 49–57) (Feltkamp et al., 1993). The E7 CTL peptide (RAHYNIVTF) was purchased from Peptron (Taejon, Korea). After 3 days of incubation at 37°C in 5% CO2, cell supernatants were secured and then used for detecting levels of IFN-γ using commercial cytokine kits (Biosource Intl., Camarillo, CA) by adding the extracellular fluids to the IFN-γ-specific ELISA plates.
CTL assay
CTL assays were performed as previously described (Sin et al., 2006; Ye et al., 2007). In brief, splenocytes were cultured in cRPMI supplemented with 1 mM sodium pyruvate and 5.5×10−5 M β-mercaptoethanol and stimulated in vitro for 5 days with 1 μg/mL E7 peptides (containing the MHC Class I epitope at amino acids 49–57) (Feltkamp et al., 1993) and 25 units/mL rIL-2 (BD, Mountain View, CA). The stimulated splenocytes were collected and used as effector cells against target cells (TC-1 cells). After background subtraction, the percentage of lysis was calculated using the following formula: 100×[(experimental release−effector spontaneous release−target spontaneous release)/(target maximum release−target spontaneous release)].
Histological analysis
Tumor-bearing animals were sacrificed and tumors were surgically excised. The tumor mass was placed in 4% formalin solution. The fixed tumor mass was bisected longitudinally or transversely and embedded in paraffin. Four to 5-μm sections of the embedded tumor tissues were made and then stained with hematoxylin and eosin (H&E). Light microscopy was used to identify the status of lymphocyte infiltration or inflammation.
SDS-PAGE, western blot assay, and sandwich ELISA
DNA-injected muscles were collected in 300 μL of lysis buffer (0.1 M Tris-HCl [pH 7.8], 0.5% Triton X-100, and protease inhibitor cocktail), followed by tissue tearing. Cell supernatant was collected after microcentrifugation. Muscle cell proteins were analyzed by 12% SDS-PAGE and then electrophoretically transferred to nitrocellulose membranes (Amersham, Piscataway, NJ). The membrane was preequilibrated with TBST solution (10 mM Tris-HCl [pH 8.0], 150 mM NaCl, and 0.1% Tween 20) containing 2% bovine serum albumin for 1 h and then reacted with anti-HPV 16 E7 monoclonal antibodies (Oncogene, Boston, MA). After three washes with TBST, the membrane was incubated with anti-mouse IgG-horseradish peroxidase (HRP) (Sigma-Aldrich) for 1 h at room temperature. The immunoreactive protein bands were visualized using the ECL detection reagents (Amersham). For sandwich ELISA, DNA-injected muscles were collected in 300 μL of lysis buffer (25 mM Tris [pH7.4], 50 mM NaCl, 0.5% Na-doxycholate, 2% NP-40, 0.2% SDS, and protease inhibitor cocktail), followed by tissue tearing. Muscle cell supernatant was collected after microcentrifugation. One hundred microliters of cell supernatants was added to a 96-well plate previously coated with anti-E7 antibodies raised in rats (previously generated in our laboratory). The plates were washed and then added with 100 μL of anti-E7 raised in mice (previously generated in our laboratory), followed by addition of anti-mouse IgG conjugated with HRP (Sigma-Aldrich). After addition of substrate, the absorbance was read at 405 nm. Recombinant E7 proteins (Kim et al., 2002; Ahn et al., 2003) were used as a positive control and were also serially diluted to obtain a standard curve.
Bioluminescence assay
Animals were injected with pCR3.1-luciferase by IM versus IM-EP delivery methods. At 0, 5, 7, 11, 15, 18, 26, and 33 days postinjection, animals were intraperitoneally (i.p.) administered D-luciferin in accordance with the manufacturer's protocol (Xenogen, Alameda, CA). Then, the bioluminescent intensity was measured using the IVIS Imaging System Series 200 (Caliper, Hopkinton, MA). Luminescence signals were acquired for 10 s. For detection of luciferase activity in spleen and the inguinal lymph nodes proximal and isolateral to the site of injection, animals were sacrificed at 7 days following IM versus IM-EP delivery of pcDNA3 (as a control) and pCR3.1-luciferase. The lymphoid organs were cut into several pieces and then dipped into a D-luciferin solution prior to detecting luminescence signals.
Statistical analysis
Statistical analysis was done by the independent-samples t-test using the SPSS 13.0 software program. The values of experimental groups were compared with the values of control group. A p-value of <0.05 was considered significant.
Results
Antitumor therapeutic effects of E7 DNA vaccines by IM versus IM-EP delivery methods
To investigate the antitumor therapeutic effects of IM versus IM-EP delivery of HPV 16 E7 DNA vaccines, animals were s.c. challenged with 2×104 TC-1 tumor cells and then treated with E7 DNA vaccines by IM and IM-EP delivery methods. As shown in Figure 1A, IM delivery of E7 DNA vaccines resulted in a significant degree of tumor growth suppression over time compared with negative control. However, IM-EP delivery of E7 DNA vaccines displayed complete suppression of tumor growth compared with IM delivery. In the IM-EP-treated animal group, however, some tested animals showed tumor formation within 15 days after the first treatment. The formed tumor regressed thereafter. Most of these animals showed no recurrence of tumors over 115 days posttreatment. These data show that IM-EP delivery is more effective at inducing antitumor therapeutic activity against E7-expressing tumor cells compared with IM delivery. We were next interested in testing whether these tumor-controlled animals by IM-EP might have long-term antitumor memory responses to a parental tumor cell rechallenge. Therefore, the tumor-controlled animals of Figure 1A were s.c. rechallenged with 4×105 TC-1 cells (20-fold more tumor cells than the first-challenged tumor cells) per mouse on the left flank and with 4×105 B16 cells (control tumor cells) per mouse on the right flank at 115 days following the first treatment (Fig. 1A). As shown in Figure 1B and C, age-matched naive mice showed formation of both B16 and TC-1 tumor cells over time. In contrast, the tumor-controlled animals by IM-EP completely inhibited TC-1 tumor formation, but failed to suppress control B16 tumor formation. This analysis shows that animals treated with E7 DNA vaccines by IM-EP can induce tumor antigen-specific long-term antitumor memory responses.

Antitumor therapeutic effects of E7 DNA vaccines by IM versus IM-EP delivery methods, and memory responses to a parental tumor cell rechallenge. Each group (n=5) of mice (C57 BL/6) was s.c. challenged with 2×104 TC-1 cells per mouse. Next day, mice were treated at the right leg muscle with 50 μg of E7 DNA vaccine (pE7) by IM versus IM-EP methods, followed by two more treatments at the left and right leg muscles in sequence at 1-week intervals. Control mice were treated with 50 μg of pcDNA3 by IM (right leg) and IM-EP (left leg) methods at the same time. Tumor size was measured twice every week and mean tumor size was recorded
Antimetastatic effects of E7 DNA vaccines by IM versus IM-EP delivery methods
To investigate the antimetastatic effects of IM versus IM-EP delivery of E7 DNA vaccines, animals were injected with E7 DNA vaccines by IM versus IM-EP delivery methods and then i.v. challenged with 3×104 TC-1 tumor cells per mouse. Figure 2A shows white and transparent metastatic nodules of a TC-1 tumor cell origin on the surface of the lungs. As shown in Figure 2B, IM delivery of E7 DNA vaccines resulted in a significant decrease in metastatic tumor nodule formation on the surface of the lungs compared with negative control. Furthermore, IM-EP delivery of E7 DNA vaccines resulted in more suppression of metastatic tumor nodule formation on the surface of the lungs, compared with IM delivery of E7 DNA vaccines. These data show that IM-EP delivery of E7 DNA vaccines can induce more effective antimetastatic activity against TC-1 cells than IM delivery. This is in line with the antitumor therapeutic effects of E7 DNA vaccines delivered by IM-EP observed here.
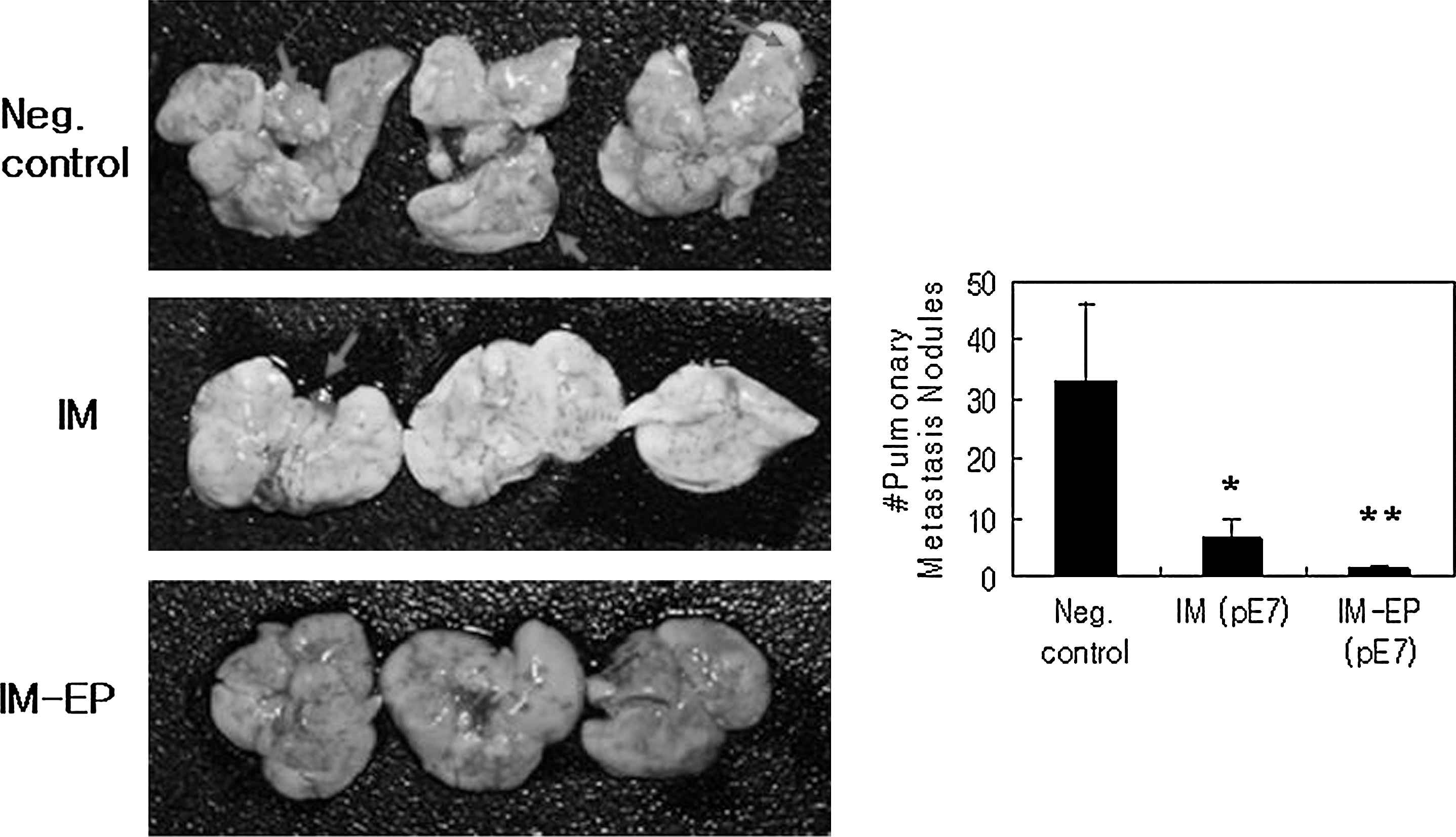
Antimetastatic effects of E7 DNA vaccines by IM versus IM-EP delivery methods. Each group of mice (n=5) was injected with 50 μg of E7 DNA vaccines (pE7) by IM or IM-EP methods twice at 2-week intervals. Control mice were injected with 50 μg of pcDNA3 by IM (right leg) and IM-EP (left leg) methods. At 4 weeks, the animals were i.v. challenged with TC-1 cells (3×104 cells/mouse). Mice were then sacrificed at 21 days after tumor cell challenge and the lung was obtained. Finally, metastasis nodules on the entire lung surface of five tested mice per group were counted. The picture shows three representative lungs per group. Values and bars represent mean of pulmonary metastasis nodule counts and the SD, respectively. These experiments were repeated with similar results. *p<0.05 compared with negative control; **p<0.05 compared with IM (pE7). Arrows indicate metastatic nodules.
Induction of antigen-specific Ab and cellular responses by IM versus IM-EP delivery of E7 DNA vaccines
We were next interested in testing immune responses to E7 antigens following IM versus IM-EP delivery of E7 DNA vaccines. For this, animals were immunized with E7 DNA vaccines by IM versus IM-EP methods and then bled for reacting with E7 proteins in ELISA. As shown in Figure 3A, IM delivery resulted in more Ag-specific IgG production at 14 and 21 days than negative control. Over these time points, IM-EP delivery displayed Ag-specific IgG production significantly more than IM delivery. The ELISA titers of equally pooled sera collected at 21 days after the first injection were determined as 800 (for IM) and 6400 (for IM-EP). We next tested Th-cell proliferative responses to E7 antigens. As shown in Figure 3B, IM-EP delivery of E7 DNA vaccines resulted in induction of Th-cell proliferative responses in an Ag dose-dependent manner. However, negative control and IM delivery showed no significant induction of Ag-specific Th-cell proliferative responses. In particular, counts per minute (CPM) by treatment with concanavalin A as a polyclonal stimulator were determined to be 1179±156 (for negative control), 1941±129 (for IM), and 1924±68 (for IM-EP), suggesting that the tested immune cells were healthy. This analysis shows that IM-EP delivery can induce Ag-specific Th-cell proliferative responses more effectively. We also tested IFN-γ production levels from CD8+ T cells (Fig. 3C). When the immune cells of animals injected with E7 DNA vaccines by IM delivery were stimulated in vitro with E7 CD8+ T-cell (CTL) epitopes, they showed IFN-γ production of 19 pg/mL, which was slightly higher than those of negative controls (14 pg/mL). When the immune cells of animals injected with E7 DNA vaccines by IM-EP were stimulated in vitro with CD8+ CTL epitopes, they showed IFN-γ production of 555 pg/mL. This is a 29-fold increase in IFN-γ production from CD8 + T cells by IM-EP delivery compared with IM delivery. This production profile of IFN-γ from CD8+ T cells was compatible with data of direct CTL assay (Fig. 3D).
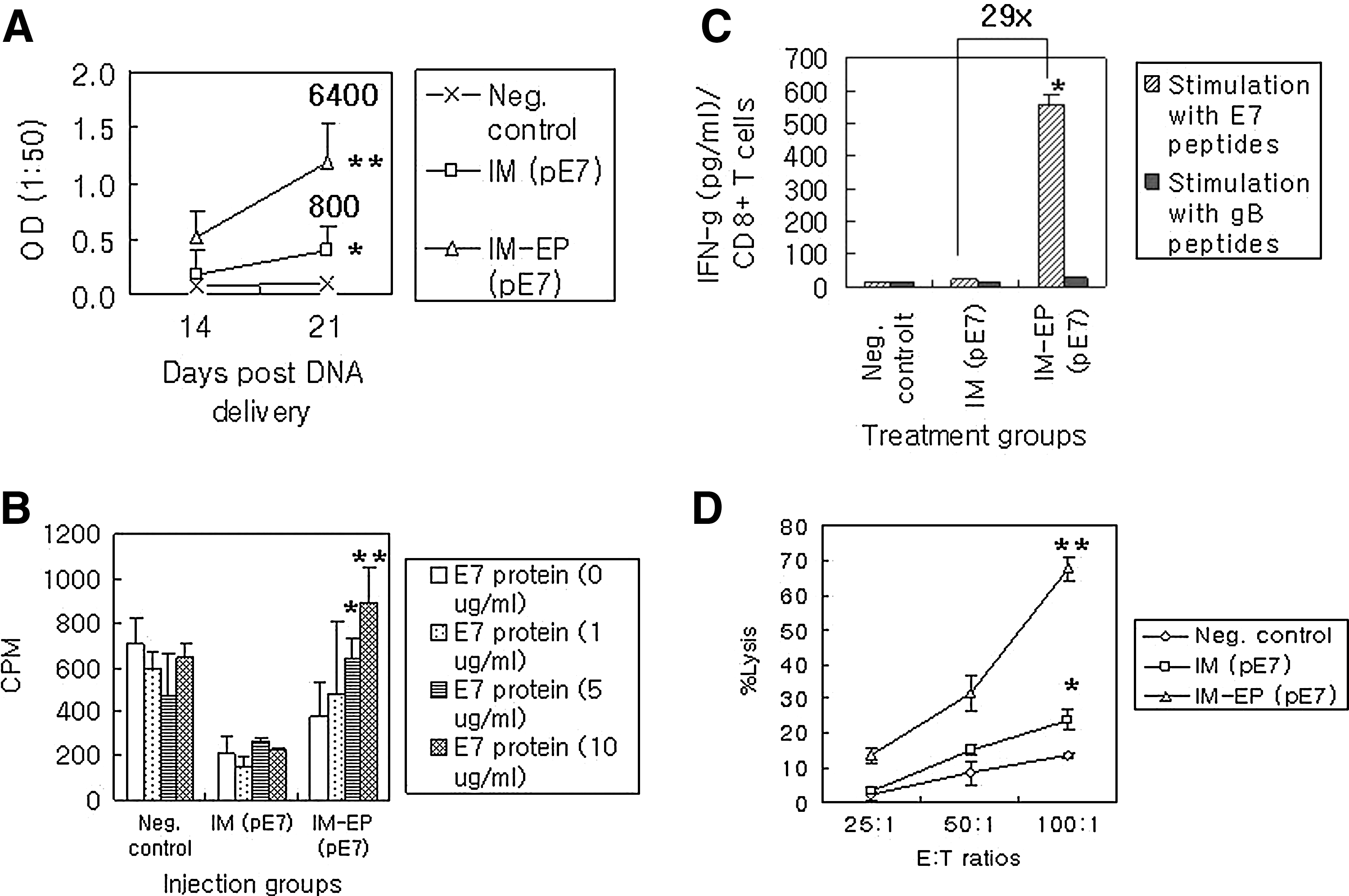
Ag-specific IgG
Immune-cell infiltration levels in the muscles by IM versus IM-EP delivery of E7 DNA vaccines
We next tested immune-cell infiltration levels at the muscle site of E7 DNA injection by IM versus IM-EP methods. Figure 4 shows H&E staining of the muscles obtained at 5 and 7 days postdelivery. Naive mice showed normal histology of skeletal muscles (Fig. 4). In particular, the upper and lower pictures in naive mice show longitudinal and transverse muscle sections, respectively. However, IM delivery resulted in infiltration of mild focal inflammatory cells and lymphocytes into the muscle site at both 5 and 7 days postdelivery. On the other hand, IM-EP delivery showed dramatically more infiltration of lymphocytes, plasma cells, and macrophages into the muscle site as well as muscle degeneration compared with IM delivery. This analysis shows that IM-EP delivery can attract immune cells (lymphocytes and APCs) into the muscle site dramatically more than IM delivery. This phenomenon might be associated with increased Ag-specific immune and antitumor responses observed here.
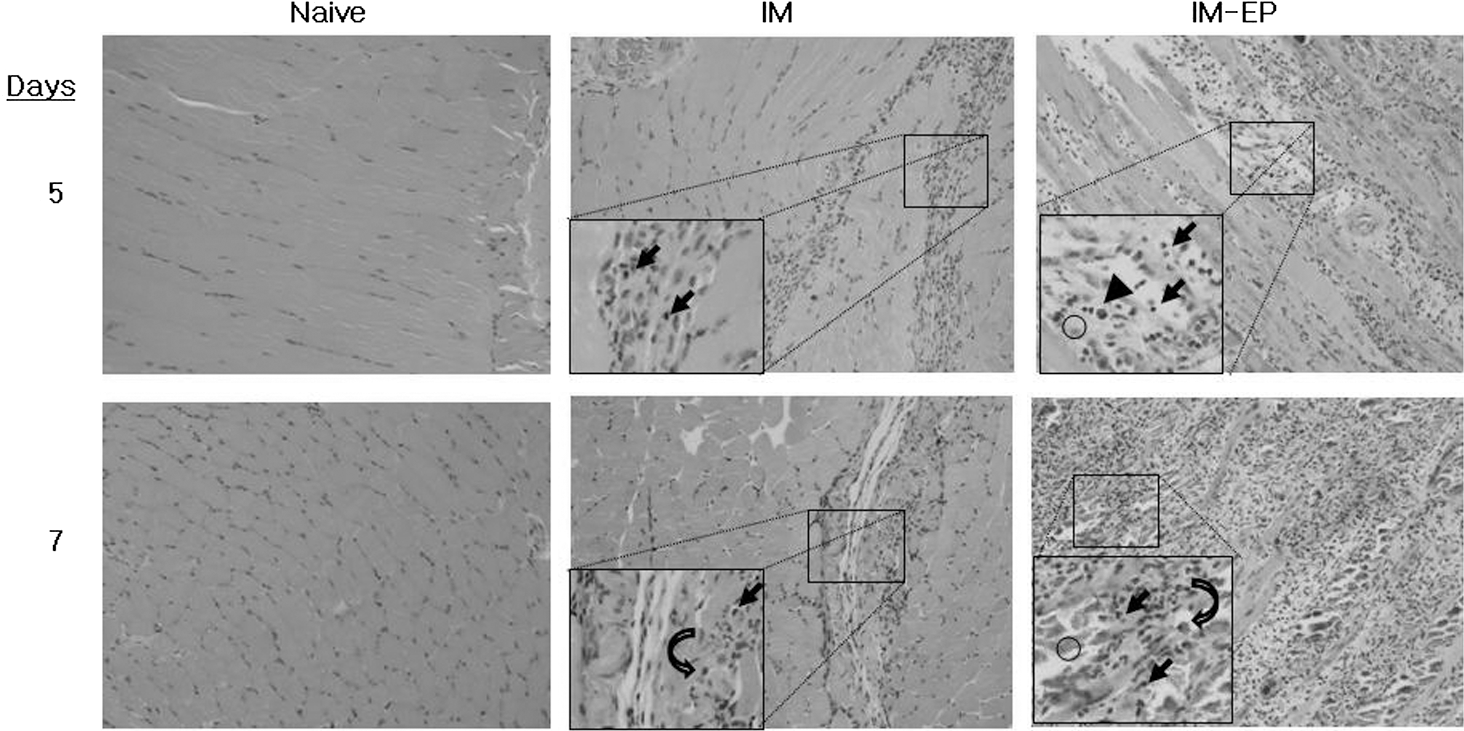
Immune cell infiltration levels at the DNA-injected site by IM versus IM-EP delivery methods. Each group (n=5) of mice (C57 BL/6) was injected with 50 μg of E7 DNA vaccines (pE7) by IM versus IM-EP methods. Animals were sacrificed at 5 and 7 days postdelivery and then the injected muscles were obtained for H&E staining. One representative slide is shown (×200). Arrow, lymphocyte; arrow head, plasma cell; curved arrow, macrophage; circle, degenerated muscle.
Antigen expression levels in the muscles and lymphoid organs by IM versus IM-EP delivery
To confirm whether IM-EP of E7 DNA vaccines results in more antigen production in the muscle, compared with IM delivery, animals were injected with E7 DNA vaccines by IM versus IM-EP methods, and then the muscles were obtained for detecting antigen expression levels by SDS-PAGE and Western blot assays as well as sandwich ELISA. However, we were unable to detect an E7 protein expression in the test samples obtained on days 5 and 7 postinjection. To further measure the difference in antigen expression levels by the two delivery methods, we used an indicator gene, luciferase. Animals were injected with pCR3.1-luciferase by IM versus IM-EP methods. Mice were then imaged on days 5, 7, 11, 15, 18, 26, and 33 postinjection using the IVIS imaging system Series 200 (Fig. 5A). Figure 5B shows luciferase activity measured by luminescent signals. Animals injected with pCR3.1-luciferase by IM-EP showed significantly more luciferase activity at the DNA injection site over time, compared with IM delivery. The luminescent signals of negative control were between 103 and 104. The luminescent signals of IM delivery at 5, 7, 11, 15, 18, 26, and 33 days postinjection were detected as 4.5×105, 1.76×106, 1.01×106, 7.02×105, 4.42×105, 2.79×105, and 4.9×104, respectively. Moreover, the luminescent signals of IM-EP at 5, 7, 11, 15, 18, 26, and 33 days postinjection were measured to be 2.02×108, 1.92×108, 1.49×108, 1.6×107, 6×106, 8.27×106, and 4.59×106, respectively. This is a more than 100-fold increase in protein expression by IM-EP compared with IM delivery. In particular, luciferase expression peaked at 7 days by both IM and IM-EP delivery methods. In IM-EP, luciferase expression was decreased dramatically at 15 days postinjection, and the decreased expression level was maintained even at 33 days postinjection. We next measured antigen expression levels in spleen and inguinal lymph nodes at 7 days postinjection when the luciferase expression was found to be highest by the two delivery methods. Negative control, IM, and IM-EP showed no luciferase activity in spleen and the inguinal lymph nodes proximal and isolateral to the site of DNA injection (Fig. 5C). In contrast, the muscle injected with pCR3.1-luciferase by IM-EP delivery (used as a positive control) displayed strong luminescent signals. Thus, these collective data suggest that IM-EP can induce an antigen production at the muscle site significantly more than IM delivery, which might be responsible for more induction of Ag-specific immune responses and antitumor therapeutic activity observed here.
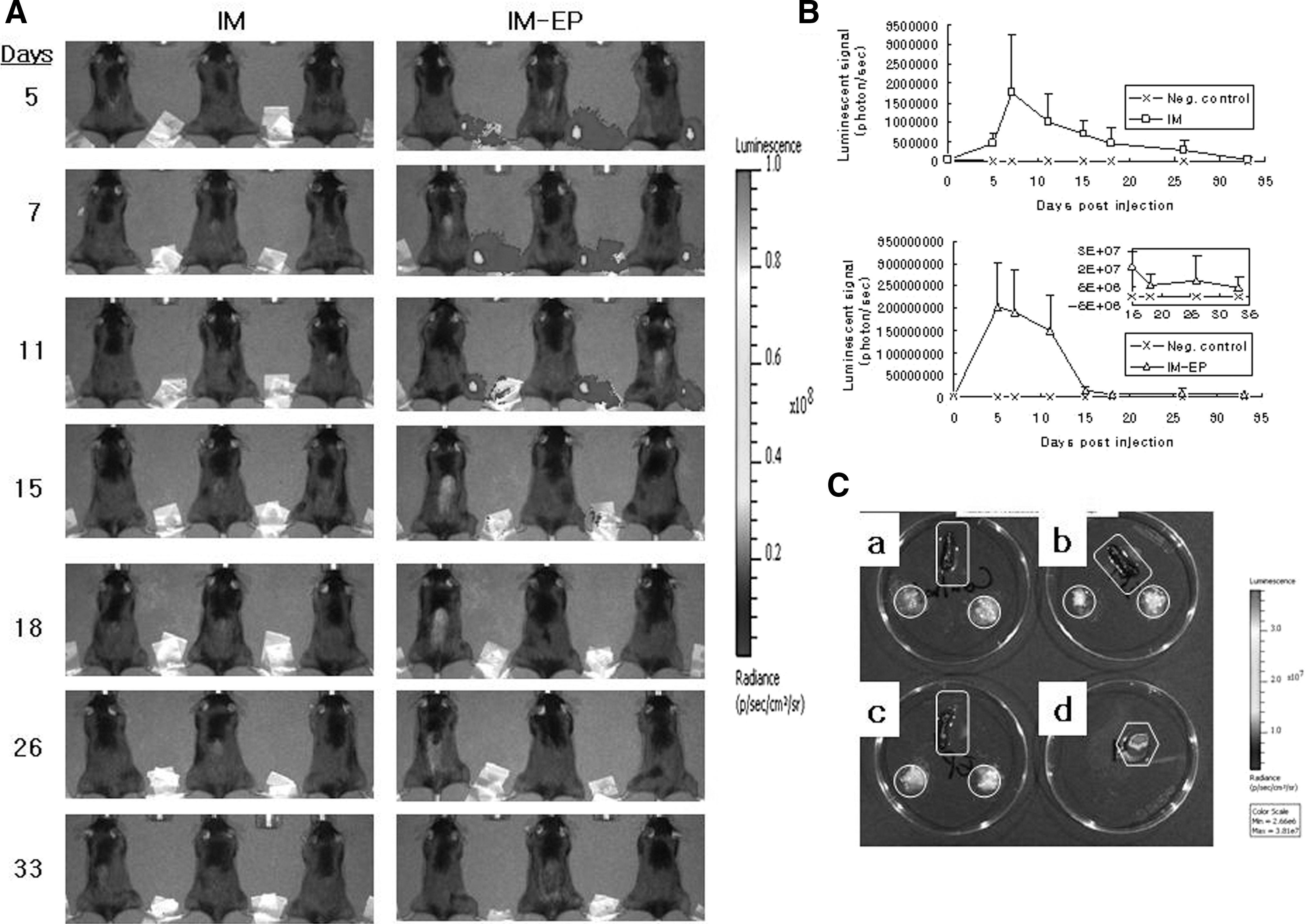
Antigen expression levels at the injection site and lymphoid organs by IM versus IM-EP delivery methods.
Antitumor therapeutic activity of E7 DNA vaccines by IM-EP in tumor-bearing animals
We previously reported that E7 DNA vaccines delivered by IM methods have little effects, if any, on eradicating large established tumors (Sin et al., 2006). To test whether E7 DNA vaccines delivered by IM-EP might have any dramatic effect on control of established tumor, animals with a tumor size of 3–4 mm in diameter were treated with E7 DNA vaccines by IM-EP. As shown in Figure 6, IM-EP delivery of E7 DNA vaccines resulted in a more dramatic inhibition of tumor growth over time, compared with control vector. In detail, none of control mice showed complete tumor regression, whereas of 11 mice treated with E7 DNA vaccines by IM-EP, 6 exhibited complete tumor regression (55%). These data show that IM-EP can lead to dramatic antitumor therapeutic activity even in tumor-bearing animals and that IM-EP might be a potent DNA vaccine delivery method for induction of antitumor therapeutic activity.
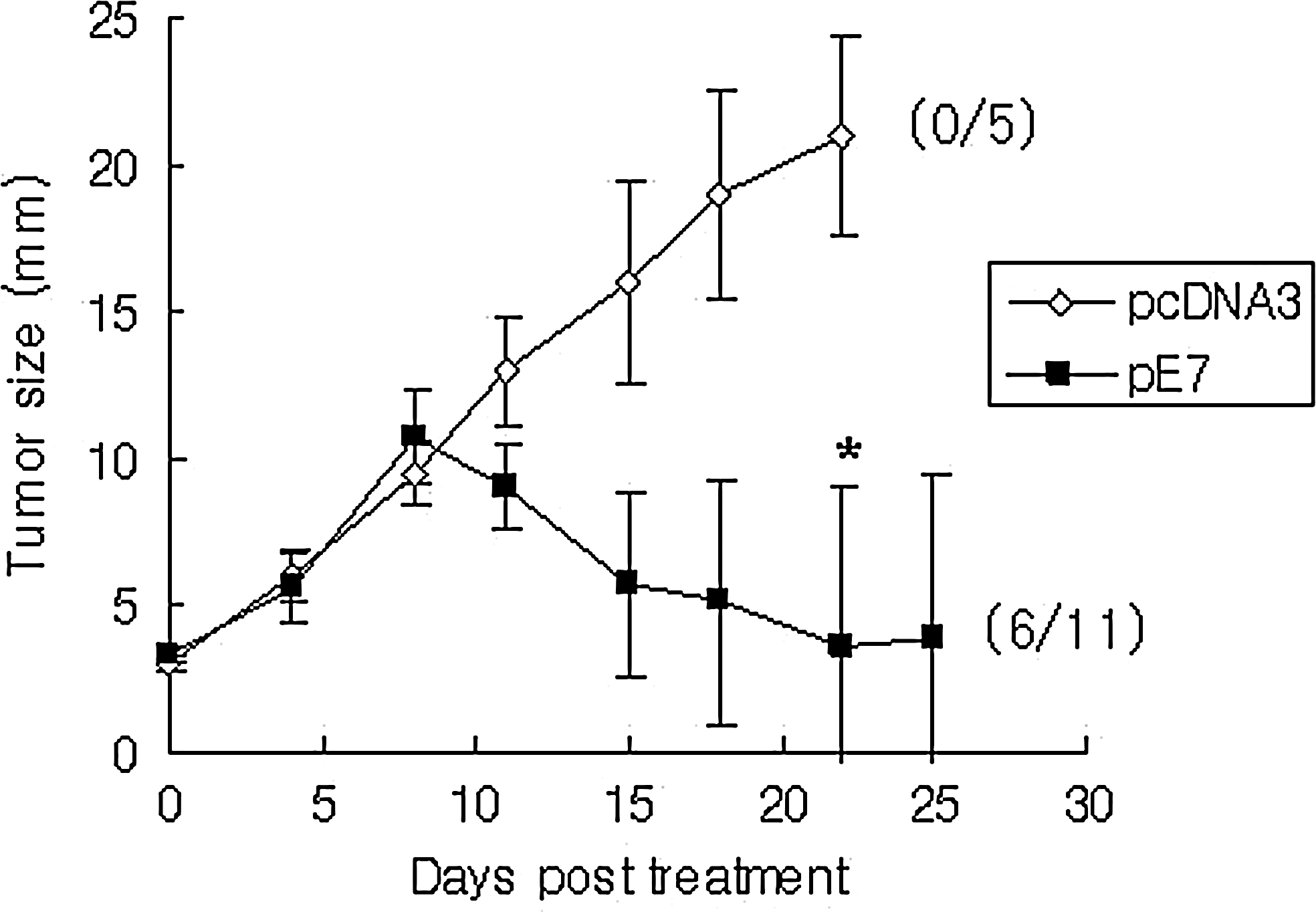
Tumor regression activity of E7 DNA vaccines by IM-EP in tumor (3–4 mm)-bearing mice. Mice were s.c. injected with 2×105 TC-1 cells per mouse. When tumors reached about 3–4 mm in mean diameter, each group of tumor-bearing mice (n=5, 11) was treated with E7 DNA vaccines (pE7) or pcDNA3 by IM-EP at 0, 4, and 11 days. Tumor size was measured over time and mean tumor size was recorded. Values and bars represent mean tumor sizes and the SD, respectively. The numbers in “(/)” indicate the number of mice showing complete tumor regression/the number of mice tested. *p<0.05 compared with negative control.
Discussion
In this study, we observed that E7 DNA vaccines delivered by IM-EP are capable of inducing more potent antitumor therapeutic and antimetastatic activity than those by IM delivery. Long-term memory responses to parental tumor cell rechallenges were also observed in tumor-controlled animals treated with E7 DNA vaccines by IM-EP. These data suggest that IM-EP of DNA vaccines can induce a dramatic level of antitumor therapeutic and antimetastatic activity as well as tumor antigen-specific long-term memory immunity. This antitumor activity was consistent with our subsequent findings that IM-EP resulted in dramatically more production of IFN-γ from CD8+ T cells compared with IM delivery. This IFN-γ production also directly correlated with the induction levels of direct CTL lytic activity against TC-1 tumor cells by IM-EP compared with IM delivery. In view of our previous findings using E7 DNA vaccines (Kim and Sin, 2005; Sin et al., 2006; Sin, 2009b), E7 DNA vaccines delivered by IM methods in this study elicited a far lower level of Ag-specific immune responses. This is likely due to the differences in DNA vaccination schemes and time points for immune cell assays. Thus, these collective data are in line with previous findings including ours that improvement of E7-specific CTL responses is associated with control of E7-expressing tumor (Cheng et al., 2002; Kim et al., 2002, 2004a, 2004b; Ahn et al., 2003; Zhang et al., 2003; Kim and Sin, 2005; Sin et al., 2006; Bae et al., 2007).
DNA vaccines have been demonstrated to be safe and tolerable in clinical trials. However, limitation of DNA vaccines' clinical applicability includes a lack of their immunogenicity possibly because of a lower expression of gene-coded proteins and a lower infiltration of APCs to DNA vaccine-injected sites in large animals (Barouch et al., 2002). Previously, we have demonstrated that Ag-specific CTL induction and antitumor resistance can be enhanced by increasing antigen expression through E7 codon optimization in an E7 DNA vaccine model (Kim and Sin, 2005). In the present study, however, we used an indicator gene, luciferase, to compare its in vivo expression levels by IM versus IM-EP delivery methods. We observed that compared with IM delivery, IM-EP led to dramatically more and longer antigen expression at the DNA-injected muscle site, as measured by luminescent signals. This level of antigen expression was detected even after 33 days postdelivery by IM-EP. However, we failed to detect antigen expression in spleen and inguinal lymph nodes proximal and isolateral to the site of DNA injection by IM and IM-EP. This is a bit surprising, based upon the finding that green fluorescent protein (GFP) was detectable in APCs of regional lymph nodes following IM delivery of GFP-expressing DNA vectors (Chattergoon et al., 1998). This discrepancy might be explained by the degree of antigen expression in APCs of the regional lymph nodes. For instance, an antigen can be detectable at the cellular level but not at the tissue level, possibly depending on the levels of uptakes of antigen-encoding DNA and its expression in APCs. Despite this, our data clearly show that IM-EP can lead to a more increased and longer antigen expression in myocytes. This likely contributes to augmentation of Ag-specific CTL induction, thereby increasing antitumor resistance. Presently, however, it remains unclear as to why E7 proteins were not detectable in vivo after IM and IM-EP delivery in this study. It is possible that E7 proteins expressed in vivo might be quickly degraded in the inflammatory environment, such that they were undetectable in vitro. This is based upon a previous report that E7 protein expression is sensitive to degradation in cells (Reinstein et al., 2000). It has been also reported that E7 protein expression is hardly detectable by immunoprecipitation methods and Western blot assay, but E7 mRNA expression is detectable to a significant level (Smotkin and Wettstein, 1986; Stoler et al., 1992). Even with a failure of E7 protein detection in myocytes, our in vivo analysis using a luciferase gene clearly demonstrates the superiority of IM-EP to IM delivery in local antigen production. Thus, IM-EP delivery of DNA vaccines might offer considerable potential to generate more antigenic proteins in vivo, which could be taken up by APCs for subsequent induction of Ag-specific immune responses. In accordance with this, we observed that IM-EP delivery induced infiltration of immune cells (lymphocytes, plasma cells, APCs, etc.) into the DNA-injected muscle site dramatically more than IM delivery. This finding is consistent with the previous findings of others that EP recruits large mixed cellular inflammatory infiltrates to the DNA injection site and these infiltrates contain 45-fold increased numbers of macrophages and 77-fold increased numbers of DCs as well as two- to sixfold increased numbers of B and T cells, compared with infiltrates following DNA vaccination alone (Liu et al., 2008). It is thought that danger signals caused by EP's needle probes and the electric pulses might be possibly associated with this immune cell infiltration. This localized effect might help to ensure that strong Ag-specific immune responses are induced and maintained. Thus, these collective data show that IM-EP of E7 DNA vaccines can induce more potent Ag-specific immune responses through more antigen production in host myocytes and, at the same time, more recruitment of immune cells to the injected muscle site.
To date, various types of vaccines (E7 peptide and heat shock protein-conjugated E7 protein vaccines, viral vector delivery of E2 proteins, and E7-fused VLP) have been tested for their effectiveness in treating precancerous cervical cancer lesions (Muderspach et al., 2000; Garcia-Hernandez et al., 2006; Kaufmann et al., 2007; Roman et al., 2007). In the studies, the vaccines were found to be effective at inducing Ag-specific CTL responses and exhibiting objective clinical benefits, such as regression of HPV-associated precancerous lesions. In cervical cancer patients, however, the vaccine efficacy was lacking. For example, therapeutic vaccines (peptide vaccines, viral vector delivery, and DC vaccines) were well tolerated without any clinical benefits, even in the presence of HPV-specific CTL in some patients (Borysiewicz et al., 1996; van Driel et al., 1999; Kaufmann et al., 2002; Ferrara et al., 2003; Santin et al., 2006). In particular, some of these patients were immunocompromised, for example, a lack or loss of MHC expression in cancer cells and an induction of immune tolerance. Thus, downregulation of the host immune system appears to be a definite negative factor for treating cervical cancer patients by therapeutic vaccination. Others including us recently reported that cytotoxic drugs and radiation can alter expression of tumor cell phenotypes (MHC class I, Fas, ICAM-1, tumor antigens) for better recognition by immune cells (Chakraborty et al., 2004; Reits et al., 2006; Ye et al., 2007; Sin et al., 2009). In the present study, E7 DNA vaccines delivered by IM-EP displayed more than 50% of complete tumor regression rates in tumor (3–4 mm)-bearing animals. This level of tumor regression is almost equivalent to that of E7 subunit vaccines previously tested (Sin et al., 2006). In this regard, DNA vaccines also have some advantages over subunit vaccines, such as cost-effectiveness in their production and longer shelf life. It is also noteworthy that the tumor-bearing animals tested here were treated by IM-EP on days 0, 4, and 11 days. However, this level of tumor eradication was not observed when tumor-bearing animals were treated by IM-EP on days 0, 7, and 14 (data not shown), suggesting the importance of IM-EP time schedules for achieving higher tumor regression rates. In particular, tumors allowed to grow for more days appear to be less therapeutically controlled, underscoring the importance of directing any therapeutic protocol toward eliminating early the kinetic disparity between tumor growth and the immune attack and maintaining the tumor-specific immune responses over time. It is also speculated that tumor regression rates might be augmented by adding either cytotoxic drugs or radiation to IM-EP delivery of E7 DNA vaccines. This is based upon our previous findings demonstrating therapeutic synergy between chemotherapy/radiotherapy and E7 subunit vaccine-based immune therapy (Bae et al., 2007; Ye et al., 2007; Sin et al., 2009). It is also known that CD73 expressed on certain tumor cell types catalyzes AMP breakdown to adenosine (Vannoni et al., 2004), which mediates suppression of T cells through the A2A adenosine receptor (A2AAR) (Huang et al., 1997). To date, blocking CD73 by adding anti-CD73 antibodies or inhibiting the activity of the A2AAR on immune cells has been proven to be effective for enhancing antitumor responses in animal tumor models (Jin et al., 2010; Stagg et al., 2010). In our unpublished data, moreover, CD73 was highly expressed on the surface of TC-1 tumor cells, suggesting that modulation of the activity of CD73 on TC-1 tumor cells possibly influences the outcome of antitumor T-cell responses generated by IM-EP of E7 DNA vaccines in controlling tumor in tumor-bearing mice. However, this remains to be further tested.
In summary, these data show that IM-EP delivery of HPV 16 E7 DNA vaccines induces more potent antitumor therapeutic and antimetastatic activity by enhancing Ag-specific CTL responses possibly through more Ag production in and more attraction of immune cells to the DNA injection site. Moreover, IM-EP of E7 DNA vaccines displayed a significant degree of tumor regression in tumor-bearing mice. Thus, this study suggests that IM-EP of E7 DNA vaccines might be a potential approach toward treating patients with cervical cancer.
Footnotes
Acknowledgments
The authors appreciate VGX International Inc. for providing Cellectra for this study. The authors also thank W.G. Ahn (Chuncheon Center, Korea Basic Science Institute) for his assistance in utilizing the IVIS 200 system. This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (2010-0008060). Moreover, this study was supported by a 2010 Research Grant from Kangwon National University.
Disclosure Statement
Jeong-Im Sin serves as a consultant to VGX International, Inc.