
Abstract
Two important polymorphisms of folate cycle enzymes, methylenetetrahydrofolate reductase (MTHFR) C677T and thymidylate synthase (TS) enhancer region (TSER) 28-bp tandem repeat, are related to risk of various types of cancer, including brain tumors, although there are few studies on this subject. A case–control study of these two polymorphisms in astrocytomas of different grades was carried out using polymerase chain reaction–restriction fragment length polymorphism, also determining the immunohistochemical expression of TS. The MTHFR 677 TT genotype was less associated with astrocytic tumors (odds ratio [OR]=0.00; p=0.0238), but the TSER polymorphism did not show any significant association. Combined genotype TT-double repeats/triple repeats (2R/3R) had a protective effect against astrocytomas (OR=0.00; p=0.0388). Expression of TS protein was observed in the majority of cases, with grade IV tumors being the exception. Moreover, the median H-score for the pilocytic astrocytomas was significantly higher when compared with that for diffuse tumors. There was an inverse correlation between the 2R/2R genotype and the highest TS-expressing tumors, and 3R/3R was relatively more frequent among the tumors grouped in the third and fourth quartiles. Our results provide support for the role of MTHFR and TS polymorphism in gliomagenesis, possibly because of the alteration of DNA methylation and repair status. Moreover, high levels of TS expression were detected in these tumors.
Introduction
Folate is an essential nutrient required for DNA synthesis, repair, and methylation. A folate-derived methyl group is transferred to a series of compounds, and this biological network is known as one-carbon metabolism (Kim et al., 1999). In particular, two enzymes, methylenetetrahydrofolate reductase (MTHFR) and thymidylate synthase (TS), have an interrelated role (Wajed et al., 2001). MTHFR is a key enzyme in folate metabolism, regulating the flow of folate groups. Its substrate, 5,10-methylenetetrahydofolate, is an intracellular form of folate required for de novo synthesis of thymidylate. The product of MTHFR activity, 5-methyltetrahydrofolate, the most important circulating form of folate, is the carbon donor for de novo methionine synthesis and, consequently, DNA methylation. On the other hand, TS competes with MTHFR for 5,10-methylenetetrahydofolate as a substrate for intracellular conversion of dUMP to dTMP, a rate-limiting step in DNA synthesis and repair (Blount et al., 1997). Therefore, individual genetic variation in these enzymes could influence the general balance between DNA synthesis, repair, and methylation. All these processes seem to be involved in gliomagenesis because of the genetic instability and the presence of methylated genes described by some studies (Gonzalez-Gomez et al., 2003).
A common polymorphism, MTHFR C677T, substituting alanine for valine at codon 222, in the N-terminal catalytic domain, results in an allozyme with decreased activity. The MTHFR 677 TT genotype is associated with lower levels of circulating folate (5-methyl-THF), accumulation of 5,10-methylene-THF, and substantially reduced levels of global DNA methylation in peripheral blood leukocytes (Kim et al., 1999; Chung et al., 2010). Also, a tandem 28-bp repeat sequence in the TS promoter enhancer region (ER), mostly double repeats (2R) or triple repeats (3R), has been identified (Horie et al., 1995). Studies in vitro and in vivo demonstrated that TS expression was genotype dependent, wherein the 3R allele was associated with higher TS expression (Horie et al., 1995; Kristensen et al., 2010).
Although there are published studies suggesting that the polymorphism of folate cycle enzymes may affect many human cancers, so far few papers relate this to brain tumor risk (Kafadar et al., 2006; Semmler et al., 2006; Bethke et al., 2008; Sirachainan et al., 2008). All these publications have the MTHFR C677T polymorphism in common, but only one paper addresses this subject with regard to TSER, without any reference to TS expression. Moreover, the number of astrocytomas was very limited, and no association between the two polymorphisms was shown. Additionally, these studies were from different populations: two from northern European countries, one from Thailand, and the other from Turkey, with one of them only involving glioblastomas. Thus, in the present study, we performed an analysis of MTHFR C677T and TSER individually and in combination in a case–control study with a series of astrocytic tumors from the state of Ceará, Brazil. We also investigated the relationship between TS genotype and gene expression.
Materials and Methods
The present study was approved by the Ethics Committee of the Hospital Complex of the Federal University of Ceará under the protocols no. 32/04 and no. 121/04, according to Resolution 196/96 of the National Council of Health, Ministry of Health, Brazil. We investigated 93 cases of astrocytic tumors of different grades (World Health Organization [WHO]) (17 grade I, 19 grade II, 14 grade III, and 43 grade IV) from BIOPSE® (Biomédica, Pesquisas e Serviços Ltda). All the samples were sectioned at 5 μm and processed for histopathological evaluation (hematoxylin–eosin staining) and immunostaining for TS. Control subjects were cancer-free individuals born and living in the same region as the cases. Among the 492 (general population) controls, 93 were matched by sex and age (±2 years) (matched population).
DNA extraction from paraffin-embedded specimens and blood
All specimens were taken from paraffin-embedded tissues. Ten sections from each sample were obtained from the blocks with adequate precaution to prevent contamination between cases, including replacement of blades between each block. Microtome holders were cleaned using xylene between cases to prevent contamination of tissue from one block to the next. Deparaffinization was done through xylol baths at 65°C and rehydrated with solutions of decreasing concentrations of ethanol and deionized water. DNA was extracted using a solution containing 20% Chelex® and 10 mg/mL proteinase K, followed by incubation at 55°C overnight. The supernatant containing DNA was collected after centrifugation in a new sterile tube.
Blood samples were obtained from the control population individuals by standard venipuncture in test tubes containing ethylenediaminetetraacetic acid (EDTA) and then genomic DNA was extracted using a salting-out method. Agarose gel electrophoresis with ethidium bromide staining was performed to ensure DNA quality. Information on the demographic characteristics and familial history of cancer was collected using a questionnaire by a trained interviewer.
Genotype analysis
MTHFR C677T: The fragment of 198 bp from exon 4 of the gene was amplified using the primer sequence and conditions described by Frosst et al. (1995). The restriction endonuclease HinfI was used to digest the 10-μL polymerase chain reaction (PCR) product to determine the genotype. The C to T transition at nucleotide 677 (valine variant) creates a new HinfI site, which generates fragments of 175 and 23 bp. PCR products were verified in a 1% agarose gel stained with ethidium bromide and the restriction digest fragments were visualized in a 7% polyacrylamide gel with silver staining (Fig. 1).
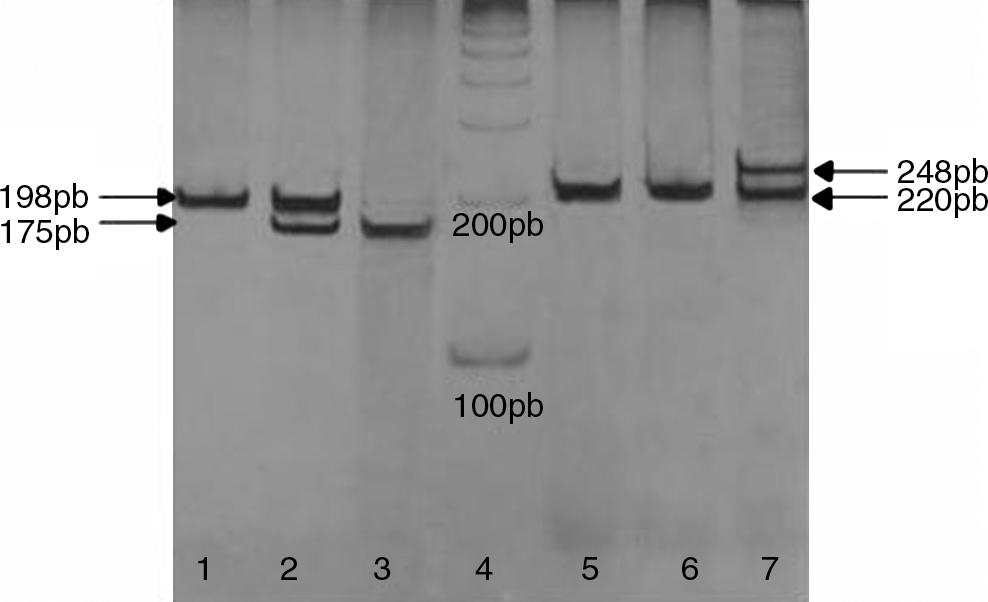
MTHFR (1, 2, and 3) TSER (5, 6, and 7) genotyping by polymerase chain reaction visualized by electrophoresis in 7% polyacrylamide gel. (1) Wild-type homozygote is identified by the presence of single 198-bp fragment, (2) polymorphic eterozygote is characterized by the presence of both 198- and 175-bp fragments, (3) polymorphic homozygote shows only the 175 bp, (4) 100-bp ladder, (5 and 6) 2R homozygote is identified by single 220-bp fragment, and (7) polymorphic heterozygote shows the presence of two fragments: 220 bp (2R) and 248 bp (3R). MTHFR, methylenetetrahydrofolate reductase; TSER, thymidylite synthase enhancer region.
TSER: The presence of a 28-bp repeat polymorphism was determined using the primer sequences and PCR conditions described by Etienne et al. (2002, 2004) except for the addition of 10% of dimethyl sulfoxide. The expected fragment sizes were 220 bp (2R) and 248 bp (3R). PCR products were visualized in 7% polyacrylamide gels with silver staining (Fig. 1).
Quality control samples were included in all laboratory analyses. Random samples (10% of case and control samples) were reanalyzed for control of laboratory procedures with the identities unknown to the laboratory staff. Concordance in the analysis was 99.5% for both polymorphisms. For the discordant samples, the genotype assays were repeated by two independent researchers to achieve 100% concordance.
Immunohistochemistry and immunostaining analysis
Immunostaining was performed according to the protocol in an earlier report (Grunda et al., 2006) and is briefly described here. Antigen retrieval was performed by pretreating deparaffinized sections with 10 mM EDTA (pH 8.0) in a pressure cooker for 5 min. After cooling, the sections were immersed in phosphate-buffered saline (PBS) containing 3% hydrogen peroxide for 10 min to block endogenous peroxidase activity. Sections were then incubated in a humidified chamber overnight at 4°C with TS primary antibody (clone TS106; dilution 1:80; Neomarkers®). After rinsing with PBS, the slides were incubated with secondary antibody followed by streptavidin–biotin–peroxidase complex (LSAB+ system; DakoCytomation®), both for 30 min at room temperature with a PBS wash between steps. The reaction was revealed with diaminobenzidine–H2O2 (DAB+ system; DakoCytomation), counterstained with Harry's hematoxylin, and mounted. A confirmed case of TS-positive human breast carcinoma was used as a positive control. Controls for primary antibody specificity included omission of primary antiserum or its substitution by normal bovine serum.
The slides were independently evaluated by three experienced technicians. Immunostaining analysis was carried out using direct light microscopy in 5–10 different fields at 400×magnification. Positivity index (PI) represents the percentage of tumors that express TS protein in each group (histological grade); the labeling index (LI) expresses the percentage of positive cells in each tumor sample; the H-score takes into account the intensity of the TS stain expressed in values ranging from 0 to 3 (0=no stain; 1=weak; 2=moderate; and 3=strong), following the methods described by McCarty et al. (1986). For both the LI and H-score, we examined at least 1000 astrocytic cells counted in several fields of the same sample with high-power magnification.
Statistical analyses
Statistical analysis was performed with the use of SPSS version 14.0. Descriptive data were expressed as frequency distributions and medians. Comparison between quantitative variables was performed by nonparametric approaches (Mann–Whitney U-test and Spearman's rank correlation). Hardy–Weinberg equilibrium for case and control populations was assessed using the chi-square test. The odds ratio (OR) with 95% confidence interval (CI), adjusted for age and sex, was calculated to estimate the relative risk of development of astrocytic tumors for the MTHFR C677T and TSER genotypes. Statistical significance was set at p<0.05.
Results
The frequency of the polymorphic genotypes in the general population was 7.7% for MTHFR 677 TT and 28.9% for TSER 3R/3R. The genetic polymorphisms in this population were in Hardy–Weinberg equilibrium. Comparison between general and matched populations showed that the two populations did not differ (Supplementary Tables 1–4; Supplementary Data are available online at
Differences in genotype distribution for MTHFR C677T and TSER polymorphisms, as well as the combined genotype of these polymorphisms, were tested between the total cases and control population (n=93). These data are presented in Tables 1 and 2, respectively. Although the genotype MTHFR 677 TT was not present in the patients, the heterozygotes were slightly more frequent, which can explain the similar allele frequencies between the two groups. A polyacrylamide gel with representative results from both polymorphisms studied is shown in Figure 1.
Reference allele.
p<0.05: statistically significant.
MTHFR, methylenetetrahydrofolate reductase; TSER, thymidylate synthase enhancer region; OR, odds ratio; CI, confidence interval; χ 2, chi-square test; NC, not calculated.
Reference allele.
p<0.05: statistically significant.
The MTHFR 677 TT genotype was absent in the astrocytic tumors. Considering the MTHFR 677 CC genotype as reference, despite that the OR cannot be considered because of a lack of TT genotype in the cases generating a zero risk, the significance found by the chi-square test (p=0.0238) is a strong indication of the association of this genotype with protective effect against the development of astrocytoma.
On the other hand, the TSER polymorphism did not show any significant associations despite a decrease in the incidence of the 3R/3R genotype observed in the patients compared with the controls (Table 1). However, when the MTHFR C677T and TSER genotypes were combined, a potential protective effect was found with the TT-2R/3R genotype in relation to astrocytic tumors (p=0.0388) (Table 2).
Table 3 shows the distribution for MTHFR and TSER genotypes according to the histological subtypes of astrocytoma. No statistically significant differences were found between these polymorphisms with regard to histological classification.
Examples of immunostaining for TS in astrocytic tumors are illustrated in Figure 2. Expression of TS protein was observed in the majority of cases (PI>90%), but grade IV tumors showed a slightly lower PI (∼80%). Moreover, the median H-scores for the pilocytic tumors (WHO grade I) was significantly higher when compared with the diffuse tumors (Fig. 3). Among the diffuse tumors, no difference was found between grades II and III, but grade IV tumors had significantly lower median H-scores.
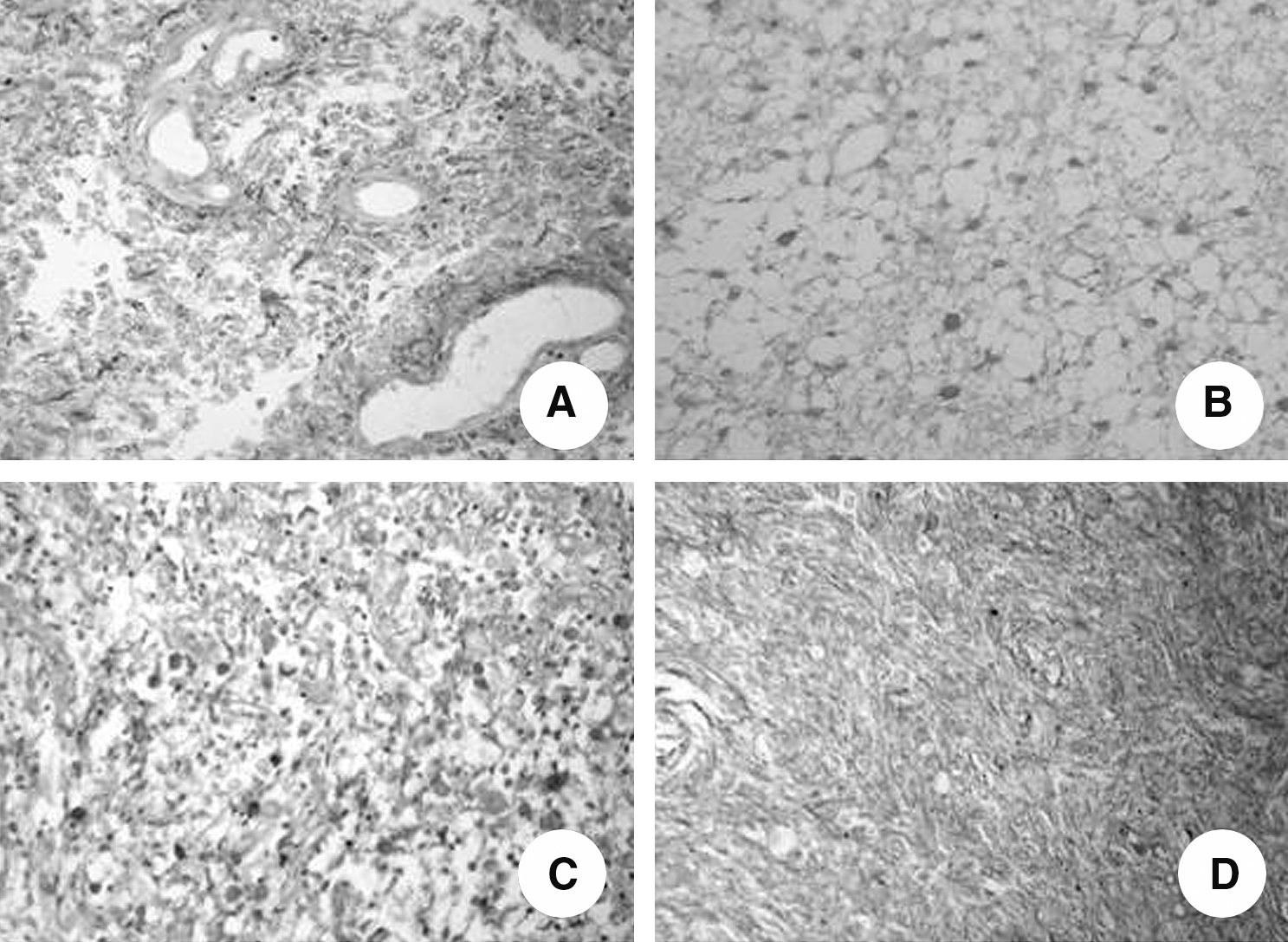
Immunohistochemistry for thymidylate synthase protein in formalin-fixed, paraffin-embedded astrocytic tumors (×400).
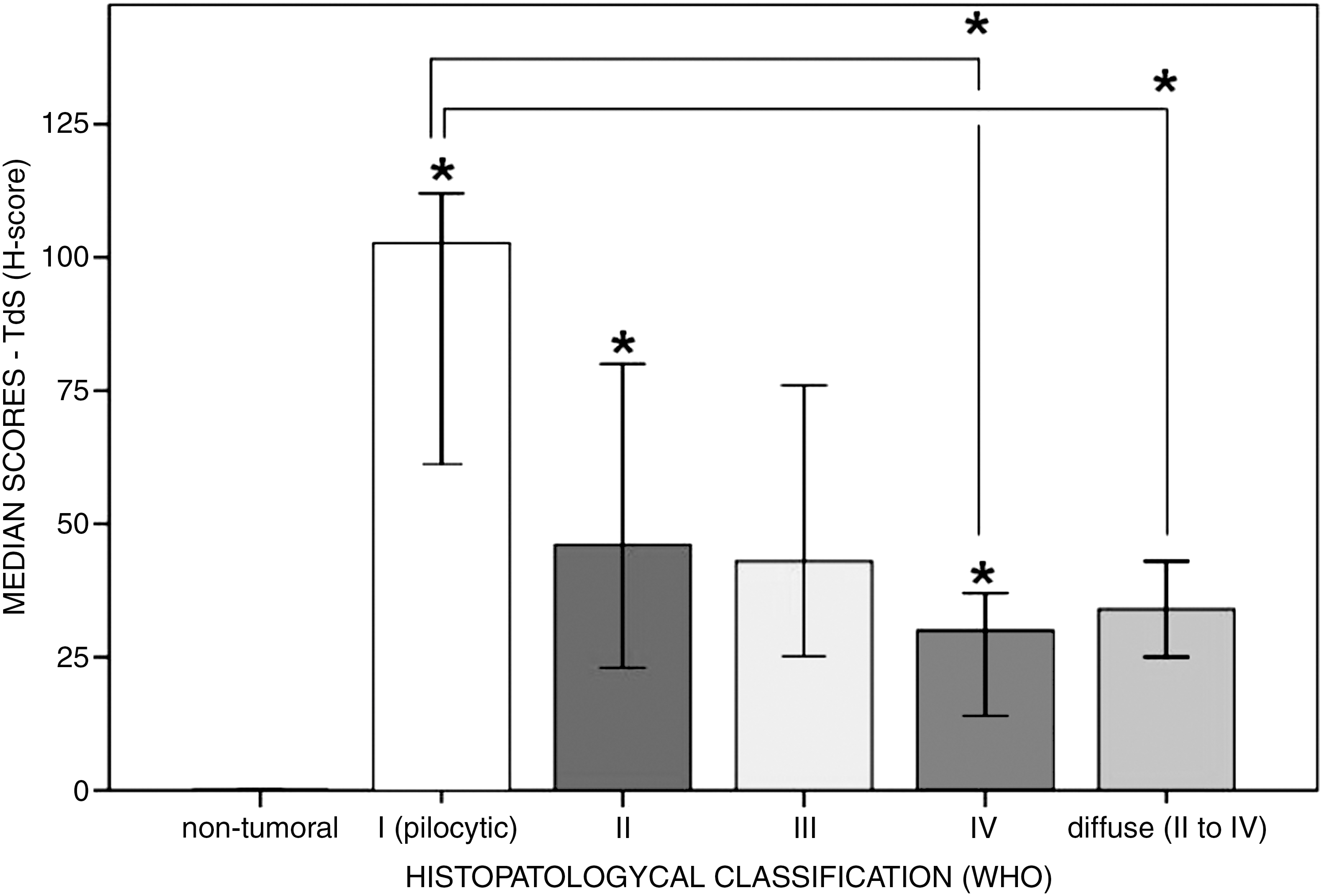
Median H-score attributed to thymidylate synthase immunostaining according to histopathological classification of the cases studied (n=93). *p<0.05 when compared with immediately preceding group or group referred to by the bar (Mann–Whitney U-test; error bars 95% CI). CI, confidence interval.
When these results were correlated to the TS genotype, a negative correlation was found between genotype 3R/3R and TS expression (R=−0.282 and p=0.002). In view of this contradictory finding, TS expression was categorized into quartiles, based on the LI scores (Fig. 4). This made it possible to demonstrate an inverse correlation between the wild-type homozygous genotype (2R/2R) and the highest TS-expressing tumors as well as a greater presence of the heterozygous genotype (2R/3R) among cases of median TS expression (LI=25%–49%). The distribution of the polymorphic homozygote (3R/3R) did not display a clear pattern, although this genotype was relatively more frequent among the tumors grouped in the third (LI=50%–74%) and fourth (LI=75%–100%) quartiles. Interestingly, the 3R/3R was the predominant genotype in cases negative for TS expression (LI<5%), which were represented by glioblastomas (grade IV), except for a single case of grade III tumor.
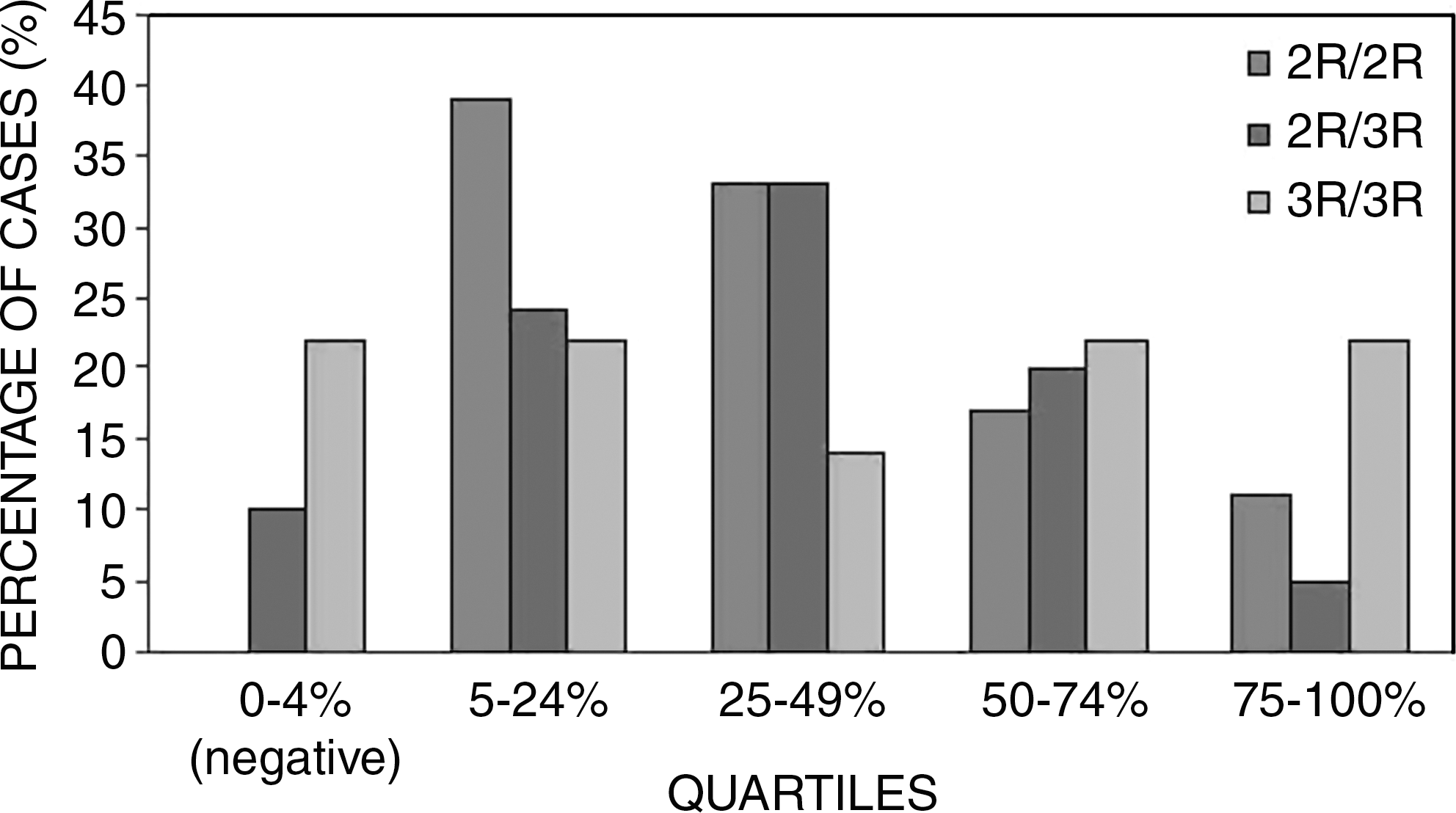
TSER genotype distribution according to quartiles of expression, based on labeling index scores of the cases studied (n=93).
Discussion
The present study analyzed the risk of the development of astrocytic tumors according to the presence of polymorphisms of two important enzymes of the folate cycle, MTHFR C677T and TSER, crossing data from DNA somatic tissue (controls) with the archived tumor tissue. This comparison between different sources of sample was in consonance with a previous analysis (Blomeke et al., 1997; Marsh et al., 2005; Weiss et al., 2007). Moreover, we assessed TS immunoexpression and performed a correlation with its polymorphic status.
In basic cancer research, there is an increasing need to establish factors that predispose to the development of cancer. In recent years, genetic factors have been increasingly recognized as major contributors to cancer risk (Weber and Nathanson, 2000; Ponder, 2001; Dumitrescu and Cotarla, 2005). Several genetic polymorphisms, including the polymorphisms of folate pathway enzymes, are reported to have an important role in the modification of cancer susceptibility. Although there are some conflicting results, it seems to depend on the cancer type. In relation to brain tumor risk, there are still few studies in the literature.
There are many variations in the genotype frequency of the polymorphisms MTHFR C677T and TSER among world populations. The Hardy–Weinberg equilibrium analyses in the present study showed that the control population was in equilibrium, but not the brain tumor samples, demonstrating an imbalance in these specific patient populations. Analyses of MTHFR C677T showed that the frequency of genotype 677 TT was similar to that for American Caucasian (Yang et al., 2008), Portuguese (Castro et al., 2003), German (Semmler et al., 2006), and Finnish (Bethke et al., 2008) populations, but showing a lower frequency compared with Chinese (Shrubsole et al., 2004) and higher than in South African (Pegoraro et al., 2004) and Thai (Sirachainan et al., 2008) populations. The frequencies of TSER polymorphism appear to be similar to that of a US population (Ulrich et al., 2005), but differing from that of a Chinese population (Zhang et al., 2005), which had higher frequency of the 3R/3R genotype (Table 4).
CE, state of Ceará; RS, state of Rio Grande do Sul.
The present study that with MTHFR C677T, although the OR could not be considered, the significant absence of this genotype in the cases indicated that the genotype 677 TT may confer a potential protective effect, thereby decreasing the risk of this cancer. The reduction of risk related to this genotype has been described for other cancers, such as hepatocellular carcinoma (Yuan et al., 2007), colorectal cancer (Cui et al., 2010), non-Hodgkin lymphoma (Kim 2007), and prostate cancer (Safarinejad et al., 2010).
On the other hand, these findings are contradictory to those of some studies that assessed brain tumors. Sirachainan et al. (2008) studied 31 glial tumors and did not find any association with MTHFR C677T polymorphism in Thai children; however, only seven of them were astrocytic tumors, and this population had a genotypic frequency distribution that differed from that in the present study. Kafadar et al. (2006) also did not find any association, but they found a higher frequency of the TT genotype, possibly because these authors analyzed high-grade gliomas without distinction.
A major study on the polymorphism of folate enzymes was conducted by Benthke et al. (2008) in a northeast European population, which provided support for the role of folate metabolism in the development of primary brain tumors. In this study, genotypes associated with increased 5,10-MTHF levels were correlated with elevated risk of developing brain tumors. Although no statistically significant association was found with the folate metabolism polymorphisms studied, their data indicated a decrease in risk related to the MTHFR 677 TT genotype in the astrocytoma group. Also, the MTHFR 677 CC genotype was associated with risk of glioblastoma when in combination with MTHFR 1298 AC/CC polymorphism. It was a well-designed study, matching the case with control from the same origin; the frequency of the MTHFR 677 TT genotype shows large variation among the different studied populations, which can explain the differences relative to our data.
One possible explanation for the protective genotype against astrocytic tumors could be the decrease in methylation of tumor suppressor genes due to the low activity of the polymorphic enzyme MTHFR, compromising the supply of methyl groups. However, this hypothesis is contradictory to the study by Cadieux et al. (2006), which indicated that inadequate methyl production promotes genomic hypomethylation and instability, explaining the increased proliferative activity in these tumors. Another plausible explanation for this 677 TT MTHFR protective effect is the increased conversion rate of dUMP to dTMP that prevents dUMP misincorporation into DNA and promotes a more efficient repair and higher genomic stability (Duthie, 2010). The removal of dUMP leads to a temporary strand break, and when it happens repeatedly, repair becomes inefficient, which may cause DNA strand breaks, chromosomal damage, and malignant cell transformation (Reidy, 1987; Blount and Ames, 1994).
With regard to the TSER polymorphism, we did not find any correlation with the risk of developing astrocytic tumors. The same was found in the study of Semmler et al. (2006). Studies that analyzed this polymorphism regarding cancer susceptibility showed many variations, according to the localization of the tumor, indicating that there is no tendency toward establishing a consensual role about this polymorphism in human cancers (Table 5).
ALL, acute lymphoblastic leukemia; GC, gastric cancer; NHL, non-Hodgkin lymphoma.
We did not observe significant tendencies in the risk analyses related to the distribution of the two genotypes in different histological grades. None of the previous studies reported its results considering each astrocytoma WHO grade and few reports performed a combined genotype analysis. Semmler et al. (2006) studied the association of glioblastoma development with MTHFR C677T, methionine synthase (MTR) A2756G, and transcobalamin 2 C776G polymorphisms; these authors found that only grade IV astrocytomas showed an increased risk related to the MTR G allele. When we performed a genotype association, we found that the TT-2R/3R genotype was significantly less associated with tumor frequency. It is important to observe that the combinations of MTHFR 677 TT and TSER 3R/3R genotypes were in low frequency among the tumor cases studied, demonstrating the complexity of genotypic combinations owing to genetic interactions and nutritional habits. The potential protective effect related to this genotype association can be due to a decreased activity of MTHFR in association with an increased activity of TS. This association would favor DNA synthesis and repair processes in detriment to methylation, promoting a more efficient repair and a higher genomic stability (Duthie, 2010).
The high TS immunohistochemical levels in the majority of tumor cases indicate the importance of this enzyme in the maintenance of genomic stability due to DNA synthesis, although some differences were observed when the number (LI) or intensity (H-score) of stained cells were assessed. The significant expression in pilocytic astrocytomas (grade I), differing from the other grades (II, III, and IV), could be explained by the common benign behavior of these cases associated with the well-differentiated status. We hypothesize that other genetic factors associated with malignancy, absent in grade I tumors, could be activated in diffuse tumors (especially in grade IV), perhaps inhibiting, at least partially, TS expression (Cadieux et al., 2006).
On the other hand, some studies that analyzed the in vitro correlation between genotype and TS expression demonstrated a higher enzyme expression associated with the genotype 3R/3R (Horie et al., 1995; Yu et al., 2008). In the present study, when immunohistochemical data were crossed with genotypes, a negative correlation was found between 3R/3R and TS expression. However, when the genotype was distributed according to expression score quartiles, we found that the 2R/2R genotype showed an inverse correlation with TS expression. Conversely, despite a variation in TS expression associated with the 3R3R genotype, its frequency was high in the higher quartiles. This lack of a clear pattern associated with the TS expression may be due to the presence of a single-nucleotide polymorphism (C-G) in the second repeat of 3R alleles, wherein the presence of 3G sequence has three to four times greater efficiency than the other sequences (2C, 2G, 3C) (Kawakami and Watanabe, 2003; Mandola et al., 2003).
Curiously, the few cases with negative TS expression were observed only among astrocytic tumors with 2R/3R and, notably, 3R/3R genotypes. Moreover, all these cases were high-grade tumors (predominantly grade IV), which are expected to show more genetic alterations. Thus, we suggest that the accumulation of molecular alterations could, at least partially, block TS expression, resulting in low DNA repair and predispose to more genetic instability. The slight decrease in TS expression observed in glioblastomas compared with other grades, demonstrated in the present study, also supports this hypothesis. These findings are in accordance with the extensive knowledge about glioblastomas, which show lower repair rates and marked genetic instability due to the sum of multiple genetic alterations that characterize their molecular and clinical malignancy (Collins, 2004).
The high levels of TS expression also raise speculation about the use of this enzyme as a target in chemotherapy for astrocytoma. Some studies have shown success in treating brain tumors with fluoropyrimidines, especially for radiosensitization of glioblastomas (5-fluorouracil and capecitabine) (Menei et al., 2005; Grunda et al., 2010). Recently, novel folate-based TS inhibitors derived from quinazolines (raltitrexed, pemetrexed, and nolatrexed) demonstrated a great potential in the treatment of gliomas (Van Triest et al., 2000; Clarke et al., 2010). A better understanding of the molecular pathway of gliomagenesis and translational studies will enhance our knowledge about these tumors and will certainly lead to further rapid improvements in clinical outcome.
In conclusion, our results provide support for the role of polymorphisms of the folate pathway enzymes MTHFR and TS in astrocytic tumorigenesis, possibly because of an alteration in methylation and repair status. Although the sample had a reasonable size, the lack of MTHFR 677 TT indicates the need for further analysis, with a higher number of cases to confirm these findings. Moreover, the high level of TS expression detected in these tumors, despite not being directly related to TSER, shows a potential targeted therapy, notably in high-grade astrocytomas. Unfortunately, we did not measure the plasma levels or dietary intake of folate and the homocysteine in our population as well as the MTHFR and TS enzyme activity in brain tumor tissues. However, it invites new speculation that would certainly warrant exploration in future studies.
Footnotes
Acknowledgment
This study was financially supported by CNPq.
Disclosure Statment
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.