
Abstract
Most known DNA replication origins in vertebrate genomes have been found to occur close to transcriptional promoters. The origin of bidirectional DNA replication of the chicken lysozyme GAS41 locus was identified in a CpG island covering the GAS41 gene promoter. In this study, we generated an α-Orc2 antibody from rabbits immunized with the C-terminal half of Orc2 for studying in vivo Orc2 binding to the lysozyme-GAS41 origin. Using the chromatin immunoprecipitation technique and quantitative real-time polymerase chain reaction, we were able to map the Orc2-binding site to a region of the lysozyme GAS41 origin that contains multiple Sp1/Sp3-binding sites comapping with two DNase I hypersensitive sites. Further, knockdown of endogenous Sp1 by RNA interference reduced specific Orc2 binding to the lysozyme GAS41 origin. These results suggest that Sp1 participates in recruiting Orc2 to the origin.
Introduction
An origin of bidirectional DNA replication was localized to a small region between the chicken genes for lysozyme and GAS41. This origin is embedded in a ∼1.8-kb largely nonmethylated CpG island spanning the 3′ end of the lysozyme gene and the 5′ end of the GAS41 gene and displays a high initiation activity mapped closely to two DNase I hypersensitive (HS) sites of the GAS41 promoter (Phi-van et al., 1998; Phi-van and Strätling, 1999; Zimmermann et al., 2002, 2007). In this study, using a chromatin immunoprecipitation (ChIP) protocol in combination with quantitative real-time polymerase chain reaction (PCR), we investigated the in vivo interaction of Orc2 and Sp1 with chromatin of the lysozyme GAS41 origin. The binding sites of these factors were mapped to the DNase I HS sites containing Sp1 motifs, and Sp1 seems to be required for the specific Orc2 binding to the lysozyme GAS41 origin.
Materials and Methods
Preparation of α-Orc2 antibodies
cDNA coding the C-terminal half of chicken Orc2 (residues 353–588) was generated from plasmid riken1_17f11 containing the full-length Orc2 cDNA (GenBank accession no. AJ720422) by PCR using primers 5′-TCTGTTCTCAAC
Cell culture, cell synchronization, and transfection
Chicken myelomonocytic HD11 (Beug et al., 1979) and hepatic DU249 cells (Langlois et al., 1974) were maintained in Iscove's modified Dulbecco's medium (IMDM) supplemented with 8% fetal calf serum, 2% chicken serum, 100 units/mL penicillin, and 100 μg/mL streptomycin at 37°C in humidified 95% air and 5% CO2. Human hepatoma HepG2 cells were grown in Dulbecco's modified Eagle's medium supplemented with 10% FCS (Goethe et al., 2007). Cells were synchronized in G1 and S phase as previously described (Zimmermann et al., 2007). To collect G2-phase cells, G1/S-arrested cells were released in fresh IMDM medium for 6 h. Small interfering RNA was transfected into DU249 cells using TurboFect siRNA Transfection Reagent (Fermentas, St. Leon-Rot, Germany) according to the instructions of the manufacturer. The sequences to silence Sp1 were 5′-ACGAGAACUCCAACAGUUCcc-3′ and 5′-GGCCUCCAGACUAUUAACCuc-3′. A nontargeting siRNA (MWG-Biotech) was used as a negative control.
Nucleoprotein preparation and ChIP
DU249 cells (3×108) grown on 145-mm dishes were treated with 1% formaldehyde (Merck) for 10 min at 37°C. After removal of the medium, cells were washed twice with cold phosphate-buffered saline (PBS), scraped off in PBS supplemented with 10 mM sodium bisulfite (pH 7.2), and centrifuged at 600 g for 5 min at 4°C. The cells were suspended in 10 mL of hypotonic buffer A containing 10 mM Tris (pH 8.0), 3 mM MgCl2, 10 mM NaCl, and 10 mM sodium bisulfite, incubated for 15 min at 4°C, and disrupted by 15 strokes of Dounce homogenization using a tight pestle. After centrifugation, the nuclear pellet was washed twice with 20 mL of hypotonic buffer A and twice with 20 mL of high salt buffer B containing 1 M NaCl, 10 mM Tris (pH 8.0), 0.1% Nonidet P40, 5 mM EDTA, and 10 mM sodium bisulfite and subsequently suspended in 1 mL ChIP buffer (150 mM NaCl, 50 mM Tris [pH 8.0], 0.5% Nonidet P40, 5 mM EDTA, and 10 mM sodium bisulfite). The sample was then sonicated by ten 30-s pulses at 4°C in a Branson Sonifier 450 and digested with 25 units of micrococcal nuclease (Fermentas) per mg DNA in the presence of 3 mM CaCl2 at 37°C for 15 min. The reaction was stopped by adding 10 mM EDTA. Following centrifugation at 12,000 g for 3 min, the supernatant containing crosslinked nucleoprotein complexes was transferred to a fresh tube. The fragment length of the nucleoprotein complexes was controlled by 2% agarose gel electrophoresis. ChIP and DNA preparation were performed as previously described (Hübner and Phi-van, 2010).
Quantitative real-time PCR
DNA purified from the immunoprecipitated nucleoprotein complexes was analyzed by quantitative real-time PCR with SYBR Green I using a 7500 Real Time PCR System (Applied Biosystems) as previously described (Hübner and Phi-van, 2010).
Nuclear extract preparation and western blot analysis
Nuclear extracts from exponentially growing cells were prepared using the protocol of Schreiber et al. (1989) except that nuclear proteins were extracted by different salt concentrations. Western blot analysis was performed as previously described (Hübner and Phi-van, 2010).
Statistical analysis
Statistical significance was evaluated by the Student's t-test. p<0.05 was considered significant.
Results
Orc2 is tightly bound to chromatin
To study chicken Orc2, an α-Orc2 antibody was raised in rabbits using a bacterially expressed C-terminal half of chicken Orc2. In immunoblots this antibody as well as an antibody against human Orc2 recognized chicken Orc2 as a polypeptide band with an apparent molecular weight of ∼73 kDa (Fig. 1A), whereas human Orc2 displayed an apparent molecular weight of ∼67 kDa (Kreitz et al., 2001; Keller et al., 2002). To examine chromatin binding of Orc2, nuclei isolated from hepatic DU249 cells were sequentially treated with increasing salt concentrations. The resulting nuclear extracts were analyzed by immunoblotting. Figure 1B shows that Orc2 was mainly extracted by high salt with 0.4−0.8 M NaCl, whereas the transcription factor Sp1 was already mobilized from chromatin with 0.2 M NaCl. These results seem to be consistent with the fact that, in contrast to transcription factors, Orc2 is tightly bound to chromatin of different nuclear compartments including the highly packaged heterochromatin (Pak et al., 1997). Further, the requirement of high salt for the extraction of Orc2 is probably also due to large Orc2-containing protein complexes binding to chromatin.
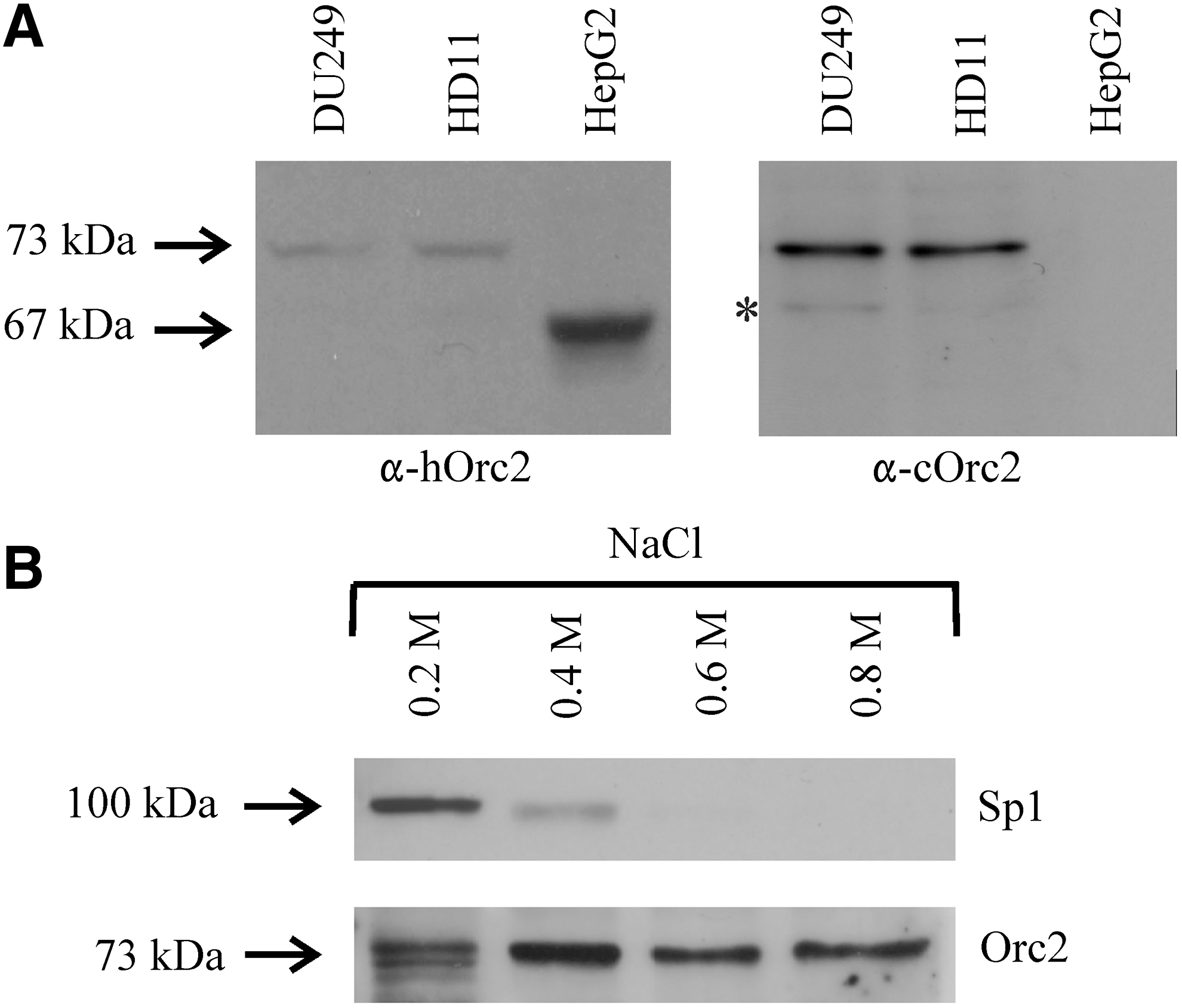
Differential mobilization of origin recognition complex (Orc) 2 from chromatin.
Orc2 and Sp1 bind to the HS sites located within the lysozyme-GAS41 origin
Previous studies using a technique of indirect end labeling have shown that two DNase I HS sites contained in a region of the origin and GAS41 promoter comap exactly with a cluster of Sp1/Sp3-binding sites (Hübner and Phi-van, 2010). To determine whether Orc2 binds to the HS sites in vivo, we established a ChIP assay. For this assay, cells were treated with formaldehyde to crosslink chromatin proteins to DNA. After removing noncrosslinked proteins by high-salt extraction, the crosslinked high-salt-resistant chromatin was fragmented by a combination of ultrasonic pulses and digestion with micrococcal nuclease to produce chromatin fragments of <0.4 kb in length (data not shown). These chromatin fragments were used for immunoprecipitation with antibodies against Orc2 and Sp1 or with IgG from preimmune serum, and the resulting immunoprecipitates were then analyzed by immunoblotting. As shown in Figure 2, Orc2 and Sp1 were found in the fraction containing sheared chromatin fragments, which were resistant to the high-salt extraction (input), indicating that treatment of cells with formaldehyde efficiently crosslinked Orc2 to chromatin. Figure 2 shows further that Orc2 and Sp1 were contained in the chromatin precipitated by α-Orc2 and α-Sp1 antibodies, but not present in the immunoprecipitates with IgG from preimmune serum.
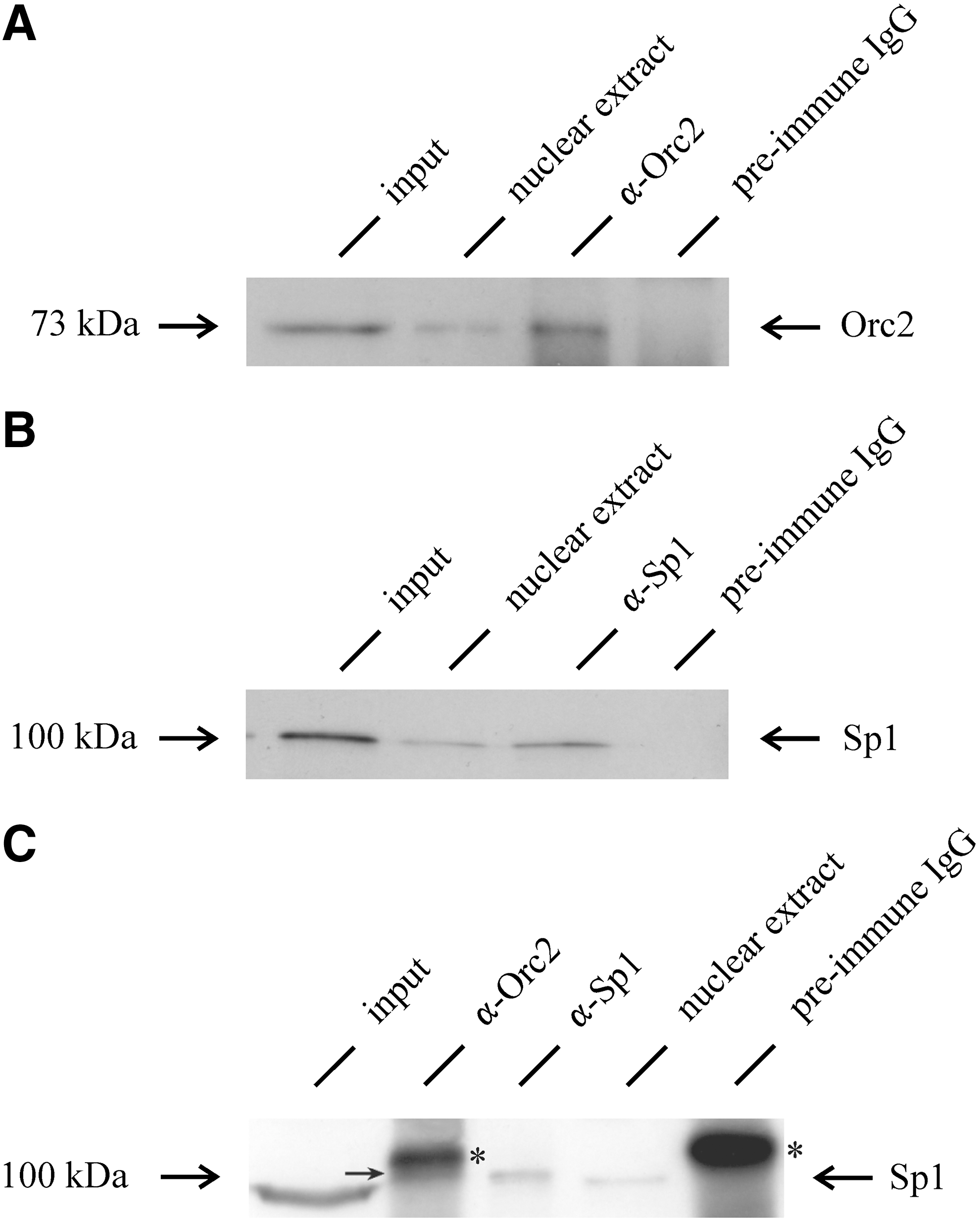
Chromatin immunoprecipitation (ChIP) with Orc2 and Sp1. Crosslinked chromatin fragments (input) from DU249 cells were immunoprecipitated with 3.75 μg α-Orc2 and 3.75 μg of preimmune IgG, or 0.2 μg α-Sp1 antibodies and 0.2 μg of preimmune IgG, for every 25 μg chromatin. α-Orc2, α-Sp1, and preimmune IgG precipitates were analyzed by western blotting with α-Orc2 antibody
Further, we analyzed the chromatin immunoprecipitated by α-Orc2 and α-Sp1 antibodies by immunoblotting with each other. Interestingly, we also found that Orc2 immunoprecipitates contain Sp1 (Fig. 2C), indicating that Sp1 and Orc2 bind to very closely adjacent sequences. This result is thus consistent with the previous finding that a fraction of Sp1 coprecipitates with Orc2-bearing chromatin (Keller et al., 2002). However, for unknown reasons we did not detect Orc2 in Sp1 immunoprecipitates (data not shown). Perhaps, in relation to total Sp1, there might be only a small Sp1 fraction that binds to origins, implicating that little Orc2 is coprecipitated with Sp1 by Sp1-specific antibodies and therefore was undetectable by immunoblotting used in this study.
DNA was isolated from fragmented immunoprecipitated chromatin and analyzed by quantitative real-time PCR to detect the sequences that were specifically enriched in immunoprecipitated chromatin. Trying to map Orc2 binding to the lysozyme GAS41 origin more exactly, we used five primer pairs designed for analysis of ∼0.8 kb including the GAS41 promoter and the origin and two primer pairs located ∼1.7 kb upstream and downstream of the GAS41 promoter (Fig. 3 and Table 1). PCR performed with these primers and genomic DNA as template showed equal amplification efficiency for each primer pair (data not shown). Then, we addressed the question of whether some sequences of the lysozyme-GAS41 locus were enriched in the fragmented chromatin fraction (input), which was generated by the high-salt extraction, ultrasonic pulses, and digestion with micrococcal nuclease, before it was used for immunoprecipitation with the indicated antibodies. DNA purified from this fraction was analyzed by the real-time PCR. As shown in Figure 4A, PCR with each primer pair resulted in approximately equal amplification, which indicates that none of the selected sequences was enriched in the fraction. Next, we analyzed the DNA purified from fragmented immunoprecipitated chromatin by real-time PCR. Figure 4B shows that the sequences LG3 and LG4 very close to the DNase I HS sites and Sp1 cluster were enriched 7.5-fold and 6.5-fold over the sequence LG1 or 2.7-/2.1-/2.8-fold and 2.3-/1.6-/2.4-fold over the adjacent sequences LG2, LG5, and LG6, respectively, in the α-ORC2 immunoprecipitated chromatin. Statistical analysis using the unpaired t-test revealed that these differences were statistically highly significant (p≤0.01). Interestingly, very similar results were obtained with DNA from α-Sp1 immunoprecipitated chromatin. The enrichment of sequences LG3 and LG4 was 8.9- and 8-fold, respectively, higher than that of LG1 in the α-Sp1 immunoprecipitated chromatin (Fig. 4C). In contrast, none of the selected sequences was significantly enriched in immunoprecipitate with preimmune IgG (Fig. 4B, C). Taken together, these results suggest that Orc2 as well as Sp1 bind to the DNase I HS sites within the lysozyme GAS41 origin in vivo.
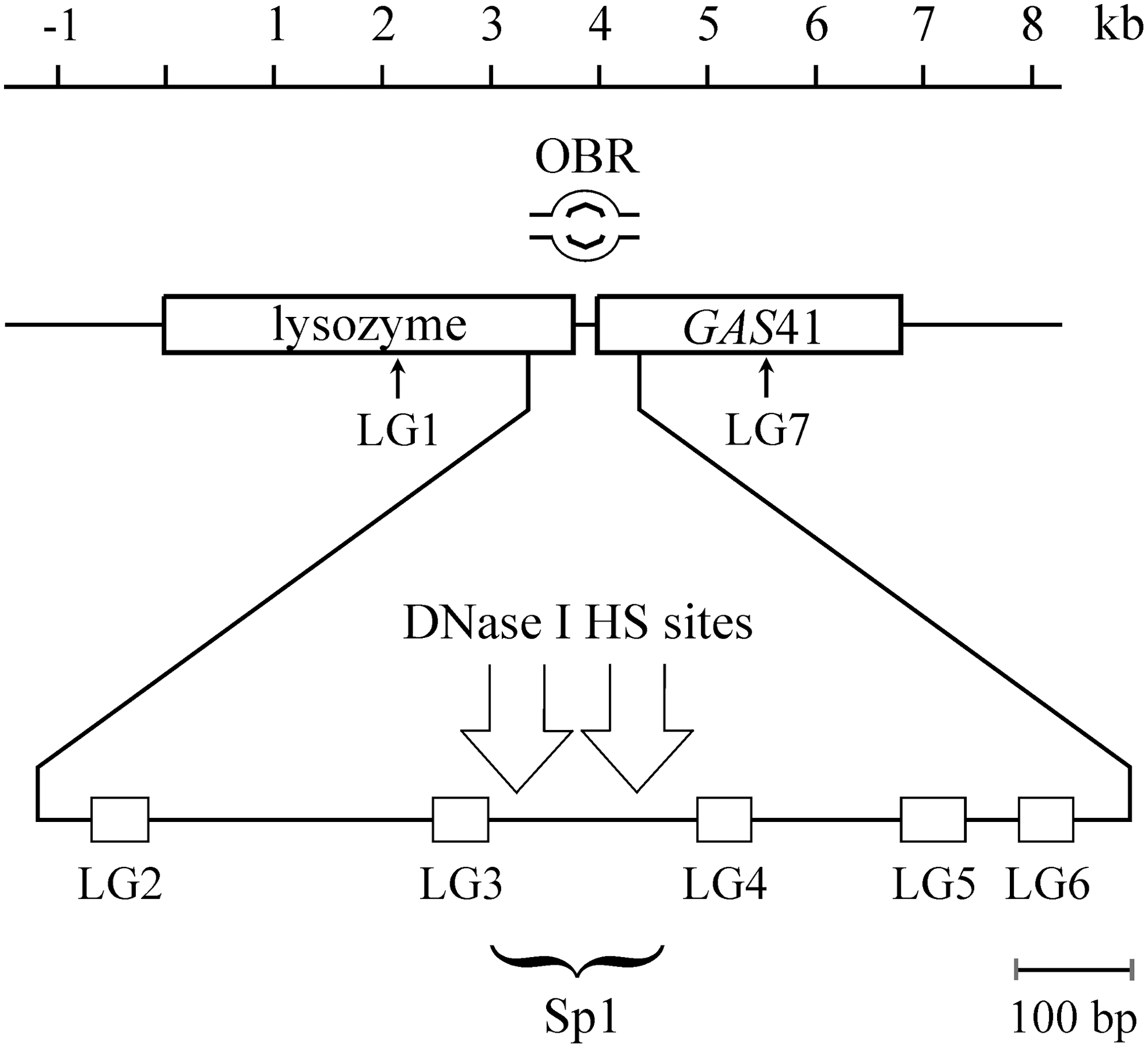
The lysozyme GAS41 locus with the coding sequences for lysozyme and GAS41 and the origin of bidirectional DNA replication (OBR). Two thin arrows and five open boxes indicate the sequences analyzed by real-time polymerase chain reaction (PCR) (LG1−7). Two DNase I hypersensitive (HS) sites comap with the Sp1 cluster (Hübner and Phi-van, 2010).
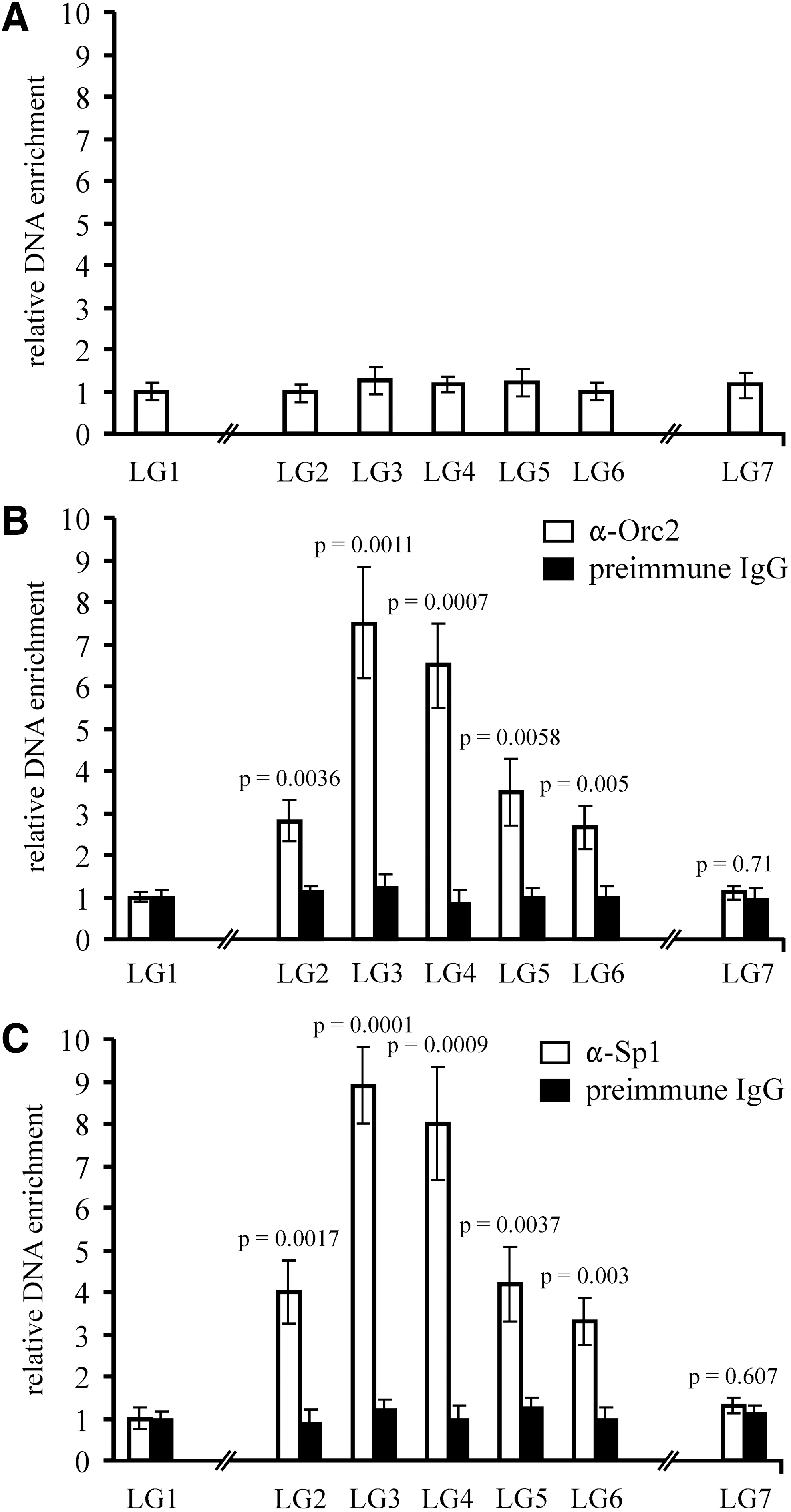
Enrichment of the lysozyme GAS41 origin sequences by antibodies against Orc2 and Sp1. DNA was purified from crosslinked chromatin fragments (input)
Fragments analyzed by real-time PCR are indicated in Figure 3.
PCR, polymerase chain reaction.
siRNA-mediated repression of Sp1 affects the Orc2 binding to the lysozyme GAS41 origin
To investigate whether Sp1 binding is a prerequisite for the Orc2 binding to the lysozyme GAS41 origin, we knocked down Sp1 by transfection of DU249 cells with Sp1 siRNA and then analyzed the effect of Sp1 silencing on Orc2 binding by ChIP assay. Western blot analysis of nuclear extracts from DU249 cells transfected with Sp1 siRNA shows a drastic decrease in endogenous Sp1 expression (Fig. 5A). Consequently, we examined the binding of Sp1 and Orc2 to the lysozyme GAS41 origin by calculating the enrichment of the origin sequence LG3 in Sp1 and Orc2 immunoprecipitates, which was normalized to the far upstream located fragment LG1. Figure 5B and C show that transfection with Sp1 siRNA also caused a significant decrease of the Sp1 binding and reduced the specific Orc2 binding to the origin by 35% in comparison to the negative control siRNA. Thus, these data suggest that Sp1 is required for binding of Orc2 to the lysozyme GAS41 origin. However, the reduction of the Orc2 binding is not strongly correlated with the siRNA-mediated decrease of Sp1 binding. One possible explanation is that Orc2, after recruiting to origins, is tightly bound to chromatin, which is possibly independent of Sp1.
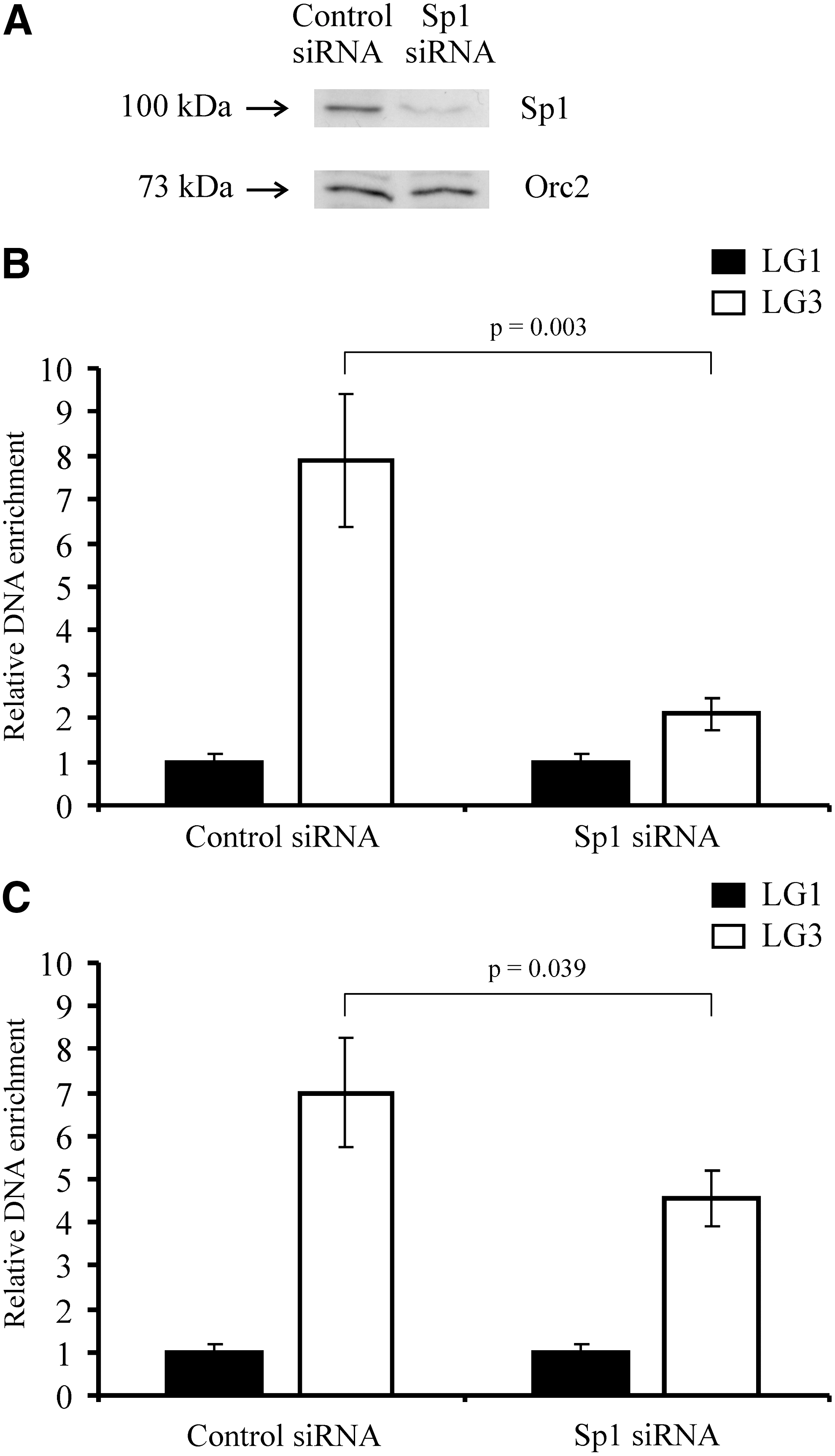
Involvement of Sp1 in Orc2 binding to the lysozyme GAS41 origin. DU249 cells were transfected with either control siRNA or Sp1 siRNA. Nuclear extracts from transfected cells were analyzed by western blotting using α-Sp1 and α-Orc2 antibodies
Orc2 is bound to the lysozyme GAS41 origin in the G1, S, and G2 phases of the cell cycle
To examine whether Sp1 and Orc2 binding to the lysozyme GAS41 origin is cell cycle regulated, we analyzed the Sp1 and Orc2 binding in synchronized DU249 cells by the ChIP assay. Figure 6A and B show the relative enrichment of LG3 in Sp1 and Orc2 immunoprecipitates, respectively. It is shown that the enrichment of LG3 by α-Sp1 as well as by Orc2 was equally high from cells of G1, S, and G2 phases, indicating that Sp1 and Orc2 were bound to the lysozyme GAS41 origin in G1, S, and G2 phases of the cell cycle. Thus, these results are consistent with the previous finding by Li and DePamphilis (2002) that Orc2 is tightly bound to chromatin throughout the cell cycle.
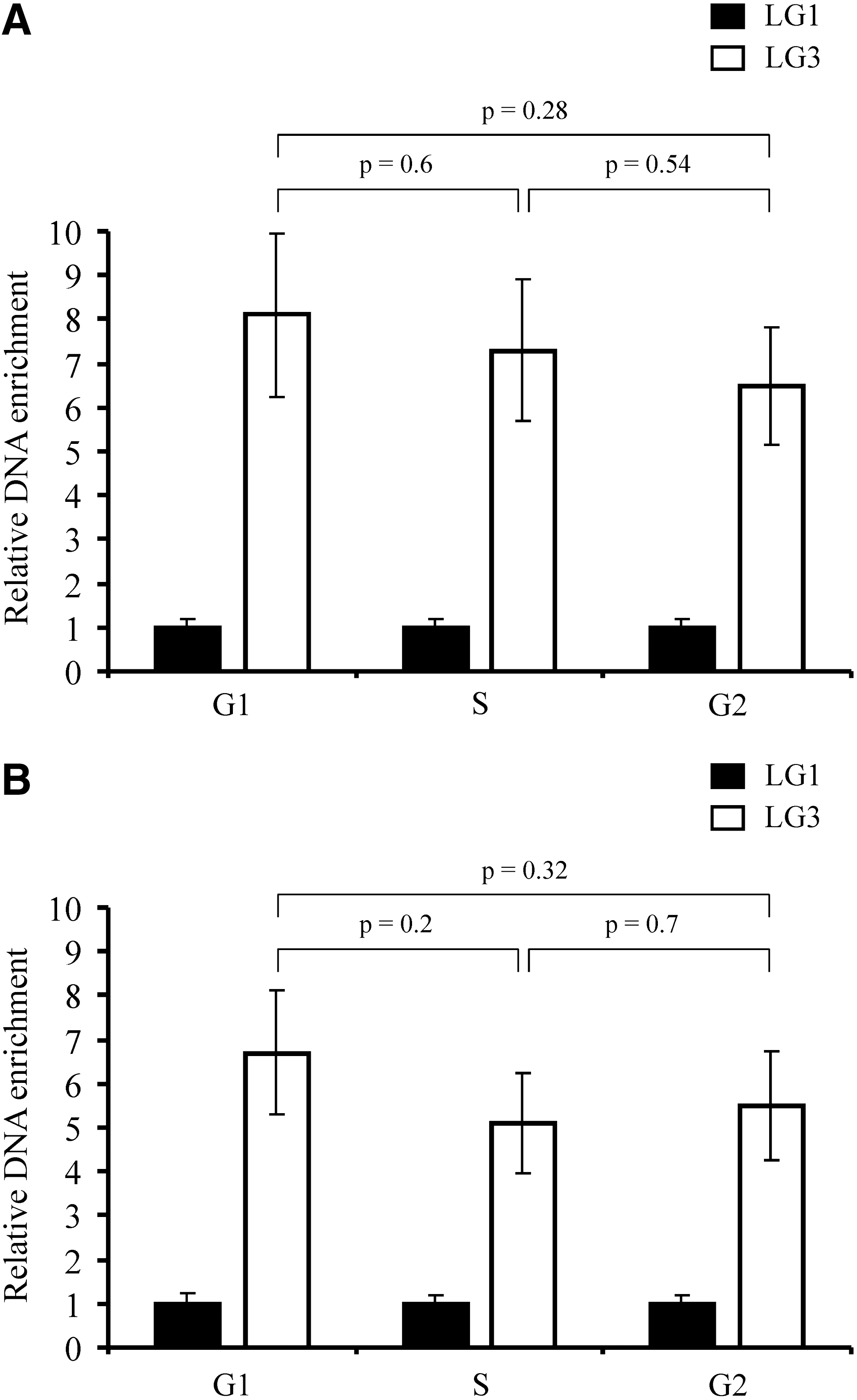
Orc2 binding to the lysozyme GAS41 origin during the cell cycle. Cells were synchronized for entering the G1, S, and G2 phase of the cell cycle. The Sp1 and Orc2 binding to the lysozyme GAS41 origin was analyzed by ChIP assay using α-Sp1
Discussion
To study the interaction of Orc2 with the chicken lysozyme GAS41 origin, we prepared an α-cOrc2 antibody. Using the ChIP assay with this antibody we found that Orc2 preferentially binds to the open chromatin of two DNase I HS sites of the lysozyme GAS41 origin containing multiple Sp1-binding sites. These results are consistent with previous findings that ORC binds to nucleosome-free regions in both S. cerevisiae and Drosophila (Eaton et al., 2010; MacAlpine et al., 2010) or to regions of low nucleosome occupancy in the Chinese hamster dihydrofolate reductase locus (Lubelsky et al., 2011). On the other hand, using EMSA with oligonucleotides derived from sequences of the lysozyme GAS41 origin, we were not able to identify any Orc2-binding sites (data not shown), suggesting that Orc2 binding to this origin is not sequence specific.
Several studies have reported positive effects of transcription factors on DNA replication. For example, the cellular oncoprotein c-myc, by interacting with the prereplicative complex, binds to the c-myc gene origin and modulates DNA replication origin activity (Dominguez-Sola et al., 2007), or other transcription factors such as NF1 and Oct-1 are able to bend the Ad5 origin, leading to a synergistic enhancement of DNA replication (Mysiak et al., 2004). It has also been reported that transcription factor binding to promoter elements can induce origin activity (Ghosh et al., 2004) or site-specific initiation of DNA replication (Danis et al., 2004).
Sp1 is a zinc finger transcription factor and has been already shown to be required for viral replication of simian virus 40, polyomavirus, papillomavirus, EBV, and cytomegalovirus (Guo and DePamphilis, 1992; Demeret et al., 1995; Baumann et al., 1999; Isomura et al., 2005). A possible role of Sp1 in DNA replication of Metazoa is also conceivable. Sp1 is essential for transcription of the house-keeping gene GAS41, which possesses a promoter without the typical TATA box as part of an upstream located CpG island (Hübner and Phi-van, 2010). Our results obtained from experiments using Sp1 siRNA support the assumption that Sp1 might play a role in the initiation process of the lysozyme GAS41 origin. Although the initiation at origins is inhibited by highly packaged chromatin, transcription activators may recruit chromatin remodelers, which in turn may open chromatin as access for the initiation and replication factors to the origins. The open chromatin structure of two DNase I HS sites at the lysozyme GAS41 origin is probably maintained by the Sp1/Sp3 binding (Hübner and Phi-van, 2010). In contrast to yeast, binding of Orc2 to origins in Metazoa is not sequence specific and therefore might require additional nuclear factors. Our results suggest that Sp1 plays such a role. The binding of Sp1 to many CG-boxes results in high concentrations of Sp1 at the lysozyme GAS41 origin, which might enable Orc2 to be recruited to the origin. Further, because Sp1 can form a tetramer, which causes DNA looping at promoters for recruiting transcription factors (Mastrangelo et al., 1991), it is also speculated that Sp1 could bend the chromatin at the lysozyme GAS41 origin to facilitate the assembly of preinitiation complexes. Therefore, it remains interesting to determine the exact role of Sp1 for the assembly of the prereplicative complex at the lysozyme GAS41 origin.
Footnotes
Acknowledgments
The authors thank Dr. W.-H. Strätling (University Medical Center Hamburg-Eppendorf) for helpful discussions, Drs. J.-M. Buerstedde and R.B. Caldwell (German Research Center for Environmental Health, Munich) for providing plasmid riken1_17f11, and Drs. R. Knippers and D. Schaarschmidt (University of Konstanz) for providing human Orc2-specific antibodies.
Disclosure Statement
No competing financial interests exist.