
Abstract
Several reports have suggested the importance of p73 polymorphisms in tumor behavior. We investigated the role of a p73 gene polymorphism in the susceptibility to cervical lesions in a southwestern European population. Peripheral blood samples were obtained from Radiotherapy and Gynaecology Departments of Portuguese Institute of Oncology (Porto, Portugal), from 1998 to 2002. We analyzed the p73 cytosine thymine polymorphism in peripheral blood DNA of 176 healthy donors, 38 high-grade squamous intraepithelial lesions (HSIL), and 141 patients with primary untreated invasive cervical cancers (ICC), by polymerase chain reaction–restriction fragment length polymorphism. Our results demonstrate a twofold increased susceptibility to the development of HSIL in women carrying the p73 AT allele (OR=2.39; p=0.022). Further, this association seems to be more evident in women with high parity (OR=12.53; p=0.007). This is in agreement with the possible role of p73 in cervical carcinogenesis, namely, in human papillomavirus-infected transition zone subjected to the action of estrogens and in conjunction with disruption of differentiation program of this squamous epithelium that occurs in HSIL phase before the next step to invasiveness and squamous cervical cancer.
Introduction
Infection per se is not sufficient to induce malignant conversion (zur Hausen, 2000; De Villiers, 2003) and only a minority of HPV-infected lesions progress to invasive cancer over a long period, suggesting that additional events are necessary for malignant transformation. In fact, cofactors like early age at sexual initiation, hormonal factors such as high parity and use of oral contraceptives, cigarette smoking, other sexually transmitted agents, deficiencies in certain micronutrients, an immunocompromised state, and various measures of socioeconomic status are relevant factors that modulate the oncogenic effect of HPV infection and promote infection persistence and carcinogenicity (Thomas et al., 2001; Matos et al., 2005).
A complex interrelation between viral and cellular genes is necessary for cell-cycle control deregulation, affecting the differentiation program and inducing the progressive proliferation and dysplasia of the epithelial cells, followed by progression to malignant conversion toward immortalization (Chu et al., 1999; De Villiers, 2003). Integration of HPV DNA may play a role in progression and maintenance of neoplasia and it has been demonstrated that protein products of HPV early genes (E6, E7) interact with growth-regulatory proteins of the human cell, p53 and pRb, contributing to a possible mechanism for an HPV oncogenic effect (Schiffman and Brinton, 1995). The development of cervical cancer and subsequent metastases clearly require more changes, involving the induction of chromosomal instability, namely, mutations and allelic loss of heterozygosity (Cai et al., 2000; zur Hausen, 2000), with the subsequent inactivation and deletion of multiple tumor suppressor genes.
p73, a gene structurally similar to p53, is localized on human chromosome 1p36.3, a region often deleted by loss of heterozygosity in a wide range of human cancers, including those of the skin, colon, breast, and liver, as well as neuroblastoma (Han et al., 1999). When overexpressed, it could activate the transcription of p53-responsive genes, such as p21, Bax, Mdm2, and GADD45, and inhibit cell growth in a p53-like manner, by inducing apoptosis (Liu et al., 2004). Data concerning allele-specific expression analysis of p73 gene polymorphisms is heterogeneous, ranging from 5.3% (marker in intron 9) in prostatic carcinomas to 42% (marker in exon 1) in lung cancers (Cai et al., 2000). As it has been reported with p53 (Koushik et al., 2005; Santos et al., 2005, 2006; Sousa et al., 2006), the importance of p73 polymorphisms in tumor behavior has been suggested (Ryan et al., 2001; Li et al., 2004a, 2004b; Niwa et al., 2005; Lee et al., 2010). p73 has a C/T polymorphism in exon 2 with two naturally occurring alleles consisting of a double-nucleotide substitution G→A and C→T at positions 4 and 14: the GC and AT alleles. The AT allele has an additional StyI restriction site, which does not exist in GC allele (Mai et al., 1998; Cai et al., 2000). The presence of this polymorphism was already reported in informative lung squamous cancers (Mai et al., 1998), esophageal squamous cell carcinomas (Cai et al., 2000), and cervical cancer (Craveiro et al., 2004; Liu et al., 2004; Niwa et al., 2004). The possible role of the p73 gene in differentiation and progression toward carcinogenesis in cervical cancer is known from immunocytochemical results (Brooks et al., 2002; Nenutil et al., 2003; Cheung et al., 2010). However, no reports have been published on a Caucasian population regarding the susceptibility to cervical lesions due to the presence of this polymorphism, which could be responsible for differences in p73 expression of tumors versus normal tissue. The aim of the present case–control study was to investigate the role of this p73 gene polymorphism in the susceptibility to cervical lesions in a southwestern European population.
Materials and Methods
Peripheral blood samples were obtained from 176 cancer-free control donors, 38 high-grade squamous intraepithelial lesions (HSIL), and 141 patients with primary untreated invasive cervical tumors from Radiotherapy and Gynecology Departments of Portuguese Institute of Oncology (Porto, Portugal), from 1998 to 2002. All samples were taken after informed consent according to the Declaration of Helsinki.
Clinical characteristics, including histological type and tumor stage (according to the International Federation of Gynecology and Obstetrics criteria), age at diagnosis, at menarche, and at menopause, number of pregnancies and of full-term pregnancies, and data about the use of oral contraceptives, were obtained from medical records. High parity women were defined as those with three or more full-term pregnancies.
DNA was extracted from the white blood cell fraction using a standard protocol. To evaluate the allelic expression of a C/T polymorphism in exon 2 of the p73 gene, polymerase chain reaction (PCR) amplification of DNA samples obtained was performed using two primers (5′ CAGGAGGACAGAGCACGAG 3′ and 5′CGAAGGTGGCTGAGGCTAG 3′), according to the method by Mai et al. (1998), with minor modifications (Invitrogen, Paisley, Scotland).
PCR was performed under the following conditions: 95°C for 9 min for activation of Taq DNA polymerase, followed by 95°C for 40 s, 58°C for 40 s, 72°C for 40 s, and 72°C for 5 min for 40 cycles. A specific 229 base pair (bp) fragment was obtained and electrophoresed on a 1.5% agarose gel stained with ethidium bromide.
The p73 gene has a polymorphism in exon 2 with two naturally occurring alleles: the GC and AT alleles. The AT allele has an additional StyI restriction site, which does not exist in GC allele. To identify heterozygous samples to this C/T polymorphism we analyzed DNA as follows: 15 μL of the PCR product were digested overnight at 37°C, with the restriction enzyme StyI. After digestion, the GC allele had one fragment of 229 bp and the AT allele had two fragments of 157 and 72 bp separated on a 3% agarose gel with ethidium bromide staining. Cases with 229, 157, and 72 bp StyI-digested fragments were judged to be heterozygous, having both GC and AT alleles—GC/AT genotype—whereas cases showing only one StyI-digested fragment were regarded as homozygous—GC/GC genotype and AT/AT genotype (Fig. 1).
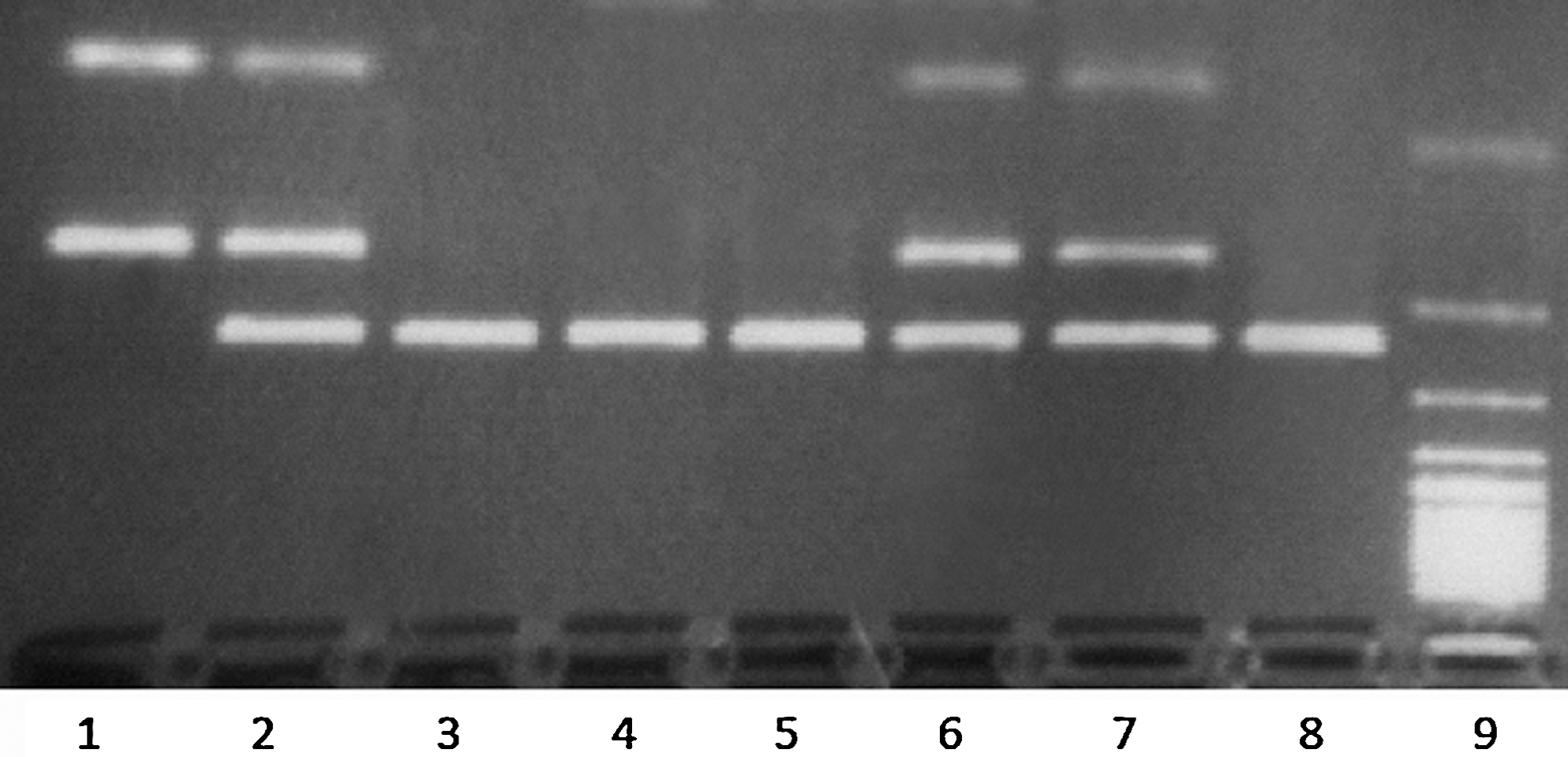
Analysis of the p73 gene polymorphism in exon 2- restriction fragment length polymorphism of the polymerase chain reaction products. Case 1, homozygous genotype AT/AT (72 and 157 bp); cases 2, 6, and 7, heterozygous genotype GC/AT (72, 157, and 229 bp); cases 3, 4, 5, and 8, homozygous genotype GC/GC (229 bp); 9, 100-bp ladder.
Statistical analysis of data was performed using computer software Statistical Package for the Social Sciences for Windows (Version 7.5). Chi-square analysis was used to compare categorical variables. The significance level was defined as p<0.05.
Results
The median age of the cases at diagnosis was 46 years, with a mean age of 47.7±12.6 years. The median age of normal donors was 48 years, with a mean age of 49.7±15.6 years. We found no significant differences in the age distribution among case and control individuals. The association between age at menarche or at menopause and the presence of the polymorphism was not significant.
The genotype frequencies in the control group of were 67.6% (119 out of 176) for homozygous genotype GC/GC, 27.3% (48 out of 176) for heterozygous genotype GC/AT, and 5.1% (9 out of 176) for homozygous genotype AT/AT. In HSIL group, the genotype frequencies were 50.0% (19 out of 38), 44.7% (17 out of 38), and 5.3% (2 out of 38), for GC/GC, GC/AT, and AT/AT, respectively. The distribution of genotypes in the group of invasive cervical cancer (ICC) patients was 67.4% (95 out of 141) for GC/GC, 26.9% (38 out of 141) for GC/AT, and 5.7% (8 out of 141) for AT/AT (Table 1).
High parity women were defined as those with three or more full-term pregnancies.
HSIL, high-grade squamous intraepithelial lesion; ICC, invasive cervical cancer.
In Table 2, we present the results of the statistical analysis of our data regarding the associations of the absence or presence of AT allele in DNA of peripheral blood samples of the three groups. Using normal donors as the reference we found a significant predominance of the presence of AT allele in the HSIL group (50.0%; OR=2.39 [1.13–5.04]; p=0.022) relatively to the ICC group (32.6%; OR=1.05 [0.65–1.71]; p=0.829). This was more evident when we analyzed both groups of cervical lesions in women with high parity. In these women, the AT allele was present in 85.7% of HSIL (OR=12.53 [1.44–28.21]; p=0.007) and in 31.3% of ICC (OR=0.95 [0.50–1.82]; p=0.876). The power of the study was 55.65%.
Logistic regression with age adjustment.
High parity women were defined as those with three or more full-term pregnancies.
Fisher exact test.
Discussion
The mechanism of cervical carcinogenesis and progression is not well understood. After infection of basal cells of cervical squamous epithelium with high risk HPV, the differentiation program is affected and a progressive proliferation and dysplasia are induced in epithelial cells (Chu et al., 1999). Cervical cancer can then develop through a multistep process with cervical lesion progression.
Protein products of high-risk HPV types 16 and 18 interact with growth-regulatory proteins of the cervical cell, p53 and pRb. When p53 is inactivated, its functions of cell cycle control and apoptosis induction are impaired. p73 is a gene with similar functions to p53 and it may play a role in a p53-independent manner. HPV E6 and E7 can inactivate p73 as p53, however, with controversial results (Marin et al., 1998; Prabhu et al., 1998; Park et al., 2001; Brooks et al., 2002; Liu et al., 2004). p73, unlike p53, is functionally inactivated by both high- and low-risk-HPV E6 proteins, by a different mechanism that does not involve degradation through an ubiquitin pathway, which could play an important role in the malignant transformation and benign condyloma formation of the cervix, respectively. E6 binds to p73, inhibiting the p73-mediated activation of p21WAF1 (CDKN1A), a p53 target primarily responsible for p53-dependent G1 arrest, and hence providing an advantage for cervix carcinogenesis (Park et al., 2001). On the other hand, p73 is transactivated by HPV E7 in cell lines and primary human keratinocytes. Oncogenic HPV E7 is able to associate with and functionally inactivate the protein (pRb) encoded by the Rb gene, which causes the release of E2F1 and the subsequent expression of a number of genes with a critical function in the control of cell proliferation and the stimulation of S-phase cell cycle entry (Brooks et al., 2002). Inactivation of pRb also disrupts the normal differentiation program of the stratified squamous epithelium, demonstrating that pRb plays an important role in the differentiation of this tissue in vivo (Balsitis et al., 2003).
Gene polymorphisms have been suggested to be important in cancer susceptibility and tumor behavior (Costa et al., 2002; Catarino et al., 2005, 2006; Coelho et al., 2005; Duarte et al., 2005; Ferreira et al., 2006). Previously published reports suggest the involvement of p73 polymorphisms in cancer susceptibility (Ryan et al., 2001; Li et al., 2004a, 2004b; Liu et al., 2004; Niwa et al., 2004, 2005; Pfeifer et al., 2005).
p73 function is regulated by post-translational modifications and protein–protein interactions (Merlo et al., 2005) and could be affected by the presence of polymorphisms, namely, in cervical cancer. Moreover, E6 and E7 are not DNA binding transcription factors, as they interact by protein–protein interactions altering the function of distinct targets regulating cell cycle, apoptosis, and differentiation, which is required for carcinogenic progression (Riley et al., 2003).
When we compared our results with the reported genotype frequencies of p73 G4C14-to-A4T14 polymorphism in normal control subjects (Table 3) from different populations, we observed that our population has a similar pattern to other European (De Feo et al., 2009) or American Caucasian populations (Li et al., 2004b; Chen et al., 2008).
RFLP, restriction fragment length polymorphism.
These results do not confirm an effect of p73 in ICC. However, we have to consider that it may be due to bias because our cases were under radiotherapy treatment and this sample may be not representative of all ICC cases. Further studies must take this into account. Our results demonstrate a twofold increased susceptibility to the development of HSIL in women carrying the p73 AT allele (OR=2.39; p=0.022). Further, this association seems to be more evident in women with high parity (OR=12.53; p=0.007). One weakness of our study is the sample size and the power of the study (55.65%). The sample size of future studies must be enlarged. However, our results are consistent with previous published reports concerning the influence of this C/T polymorphism in cervical cancer (Niwa et al., 2004). Further, our results are consistent with a role of p73 in carcinogenesis (Brooks et al., 2002; Nenutil et al., 2003; Liu et al., 2004; Niwa et al., 2005) and its association with estrogen activity, as suggested before (Padma et al., 2000).
The demonstration of an increase in 16(-hydroxylation of estradiol to 16(-hydroxyestrone, which covalently binds to the estrogen receptor in the estrogen highly sensitive transformation zone of cervix in the presence of high-risk HPV infection, suggested the synergistic activation of proliferation by viral oncoprotein cell cycle deregulation and estrogen receptor signaling, prolonging the estrogen effect (Newfield et al., 1998; Jin et al., 1999; Shew et al., 2002; De Villiers, 2003; Miller, 2003; Riley et al., 2003). In addition, p73 has a demonstrated role in the regulation of nuclear entry of goat uterine estrogen receptors and the consequent stimulation of transcription of estrogen-responsive genes (Padma et al., 2000). On the other hand, there is evidence that demonstrates the potent cocarcinogenic activity of the HPV16 E7 oncoprotein with estrogen in the mouse cervix, HPV16 E6 being only a modulator of E7 estrogen cervical carcinogenesis (Riley et al., 2003). E7 expression, in conjunction with estrogen, is sufficient to induce the requisite components of carcinogenic progression, including hyperproliferation, inhibition of epithelial differentiation leading to high-grade dysplasia, insufficient compensatory induction of apoptosis, and centrosome copy number elevations. Estrogen itself increases reproductive tract squamous epithelial proliferation and can induce direct DNA damage via its catechol metabolites. This could be the potential initiating step in this model of cervix carcinogenesis complemented and amplified by E7 (Riley et al., 2003).
An altered expression of p73 protein has been reported during the development of cervical lesions, suggesting the involvement of this protein. On the initiation of HPV infection, the degree of keratinization (differentiation) is a pathological feature of cervical tumors in a model of a local field effect of genomic instability that chronologically affects the clonal evolution. SIL can be classified as low grade (LSIL) or high grade (HSIL), depending on the extent to which epithelial differentiation is altered. Low-grade disease is characterized by abnormal differentiation in the lower third of the epithelium and HSIL may start to invade below the basal layer of the epithelium leading to metastatic disease. In the majority of the cases, if not all, the transition from HSIL to cancer coincides with integration of high risk HPV DNA, especially HPV16 and HPV18 into the host genome (Tommasino et al., 2003). Integration provides a selective advantage to cervical epithelial precursors of cervical carcinoma, causes the increased expression of two viral transforming genes, and is highly correlative with increased levels of E7 protein synthesis (Jeon et al., 1995). The intensity of E7 expression was inversely correlated with differentiation grade of the tumor area, so that the undifferentiated areas contained higher E7 levels. These data suggest that an increase in the E7 expression level could be correlated with a decrease in the differentiation level and that the E7 level reached in HSIL is sufficient for malignant progression (Fiedler et al., 2004). E7 has the ability to disrupt squamous epithelial differentiation and cell cycle regulation (Riley et al., 2003) and delays keratinocyte differentiation in vivo (Balsitis et al., 2003).
The association between p73 G4C14-to-A4T14 polymorphism and HPV16 status in squamous cell carcinoma of the head and neck was already stated and it was suggested that the p73 variant genotypes may be a positivity marker in patients with these tumors, particularly oropharyngeal cancer (Li et al., 2004a; Chen et al., 2008; Ji et al., 2009). This study indicates that the p73 polymorphism could act as a cofactor of HPV in the initiation of cervical carcinogenesis, particularly in the transformation zone of HPV-infected women. This supports evidence for a genetic factor in the development of cervical lesions and help to explain the observation that not all high-risk, HPV-infected HSIL lesions will progress to invasive carcinomas. Larger-scale molecular studies are needed to confirm the role of p73 polymorphism in the development of cervical lesions.
Footnotes
Acknowledgment
This work was supported by the Portuguese Ministry of Health, Projects no. 38/2007 and 32/2007 (Comissão de Fomento da Investigação em Cuidados de Saúde).
Disclosure Statement
No competing financial interests exist.