
Abstract
Curcumin (diferuloylmethane), an active component of the spice turmeric, induces apoptosis in several types of malignancies. However, little is known about its anticancer activity in small cell lung cancer (SCLC). SCLC represents a highly malignant and particularly aggressive form of cancer, with early and widespread metastases and a poor prognosis. In this study, we found that curcumin does not activate caspase-8 cleavage or alter the expression of apoptotic receptors FAS and TRAIL in NCI-H446 cells, suggesting that curcumin-induced apoptosis is not associated with death receptor-mediated pathways in these cells. Instead, curcumin caused apoptosis by increasing Bax expression while decreasing the expression of Bcl-2 and Bcl-xL. Curcumin induced a rapid decrease in mitochondrial membrane potential and the release of cytochrome c into the cytosol, followed by activation of caspase-9 and caspase-3. In addition, curcumin-induced apoptosis was accompanied by an increase of intracellular reactive oxygen species (ROS) level. These results indicated that a ROS-mediated mitochondrial pathway played an important role in the process of curcumin-induced apoptosis of human SCLC NCI-H446 cells.
Introduction
Targeting the apoptotic machinery is one approach to selectively kill cancer cells while sparing normal ones. Apoptosis is a ubiquitous and highly regulated mechanism by which cells undergo programmed cell death (Boyce et al., 2004). Apoptosis is mediated by a family of caspases, with two pathways leading to caspase activation: the death receptor pathway and the mitochondrial pathway (Johnstone et al., 2002). The death receptor pathway is initiated by the ligation of transmembrane death receptors, such as the Fas, which activate caspase-8 and subsequently effector caspase-3. The mitochondrial pathway requires disruption of the mitochondrial membrane to release cytochrome c, which functions with Apaf-1 to activate caspase-9, thereby promoting the activation of effector caspase-3.
Despite advances in chemotherapy, the prognosis of lung cancer remains poor. Natural products were the main source of healthcare products in ancient times. In modern medicine, natural products remain a major source of new drug development. Curcumin is the primary bioactive component isolated from turmeric, a dietary spice made from the rhizome of Curcuma longa, which has been used in traditional medicine in China and India for thousands of years. Extensive research has recently revealed important functions of curcumin. The medicinal value of curcumin has been well recognized for its anti-inflammatory, antimicrobial, wound healing, and antitumor activities (Manju and Nalini, 2005). Several studies have indicated that people in southeastern Asian countries have a much lower risk of acquiring colon, gastrointestinal, prostate, breast, ovarian, and other cancers than Western populations (Sinha et al., 2003; Dorai and Aggarwal, 2004). It is likely that constituents of their diets, such as curcumin, garlic, and ginger, may play a role in the prevention of such cancers. Curcumin has potential as a chemopreventative and chemotherapeutic agent, but its interactions with clinically relevant SCLC are poorly characterized. The purpose of this study was to identify the mechanism of curcumin-mediated apoptosis in human SCLC NCI-H446 cells.
Materials and Methods
Chemicals and antibodies
Curcumin (1,7-bis[4-hy-droxy-3-methoxyphenyl]-1,6-heptadiene-3,5-dione) was purchased from Sigma-Aldrich, dissolved in DMSO at a stock concentration of 10 mM, and diluted to the indicated concentration with RPMI 1640 medium. The antibodies for Apaf-1, Bcl-2, Bcl-xL,Bax, cytochrome c, and cleaved caspase-3, −8, and −9 were purchased from Cell Signaling Technology. The antibodies for Fas, TRIAL, survivin, β-tubulin, and GAPDH were purchased from Santa Cruz Biotechnology. Other chemicals were purchased from Sigma-Aldrich.
Cell culture
NCI-H446 (human SCLC) cells were obtained from the Chinese Academy of Medical Sciences. The cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 mg/mL). The cells were incubated at 37°C in a humidified atmosphere with 5% CO2.
Cell viability assay
Cell proliferation was determined using the conversion of MTT to formazan via mitochondrial oxidation. NCI-H446 cells were seeded in 96-well plates with an average of 2000 cells/well. Cells were incubated overnight and then treated in quintuplicate with 3.75, 7.5, 15, 30, 60, and 120 μL curcumin for 24, 48, and 72 h. After treatment with curcumin, the medium was replaced with 0.5 mg/mL MTT medium and incubated for 4 h. The MTT solution was removed from the wells, and the formazan crystals were dissolved in DMSO. Finally, the optical density was read by a microplate reader (Molecular Devices) at a wavelength of 490 nm. All experiments were repeated at least three times.
Observation of cellular ultrastructure by transmission electron microscopy
NCI-H446 cells were fixed with 2.5% glutaraldehyde in 0.1 M phosphate-buffered saline (PBS, pH 7.4) overnight at 4°C and postfixed in 1% osmium tetraoxide for 30 min. After washing with PBS, the cells were progressively dehydrated in a 10% graded series of 50%–100% ethanol and propylene oxide and embedded in Epon 812 resin. The blocks were cut into ultrathin sections using a microtome, and the sections were stained with saturated uranyl acetate and lead citrate. The ultrastructure of the cells was then observed under a transmission electron microscope (JEM-1230; Jeol).
Hoechst DNA staining
Nuclear changes that are characteristic of apoptosis were visualized by Hoechst DNA staining. Briefly, NCI-H446 cells were seeded at 2×105 cells/well into six-well flat-bottomed tissue culture plates containing sterile 22×22 mm coverslips. The cells were cultured for 12 h to allow adherence to the coverslips and then exposed to 7.5, 15, and 30 μM curcumin for an additional 48 h. Coverslips were washed with PBS containing 5% heat-inactivated FBS and incubated with 100 μg/mL Hoechst 33342 trihydrochloride dye in PBS for 10 min at room temperature in the dark. Finally, coverslips were washed with PBS containing 5% FBS and then photographed using a fluorescence microscope (Olympus Optical).
DNA laddering assay
NCI-H446 cells (2×106) were harvested and lysed in cold buffer containing 10 mM Tris-HCl (pH 8.0), 150 mM NaCl, 2 mM MgCl2, 1 mM dithiothreitol (DTT), and 0.5% NP-40 on ice for 40 min. Lysates were centrifuged, and the pellets were resuspended in cold buffer containing 10 mM Tris-HCl (pH 8.0), 350 mM NaCl, 1 mM MgCl2, and 1 mM DTT on ice for 20 min. Lysates were then extracted once with a phenol:chloroform:isoamyl alcohol mixture (25:24:1), and DNA was precipitated with 10 mM MgCl2 and 2.5 volumes of 100% ethanol overnight at −20°C. DNA was collected by centrifugation at 14,000 g for 20 min, resuspended in TE buffer (10 mM Tris-HCl [pH 8.0] and 1 mM EDTA) with 0.1 mg/mL RNase A, and incubated at 37°C for 1 h. Proteinase K (1 mg/mL) was added and the mixture was incubated at 37°C for an additional hour. Fragmented DNA was then electrophoresed in 1.5% agarose gels containing 0.5 μg/mL EB.
Annexin V–FITC/PI double staining assay
The cells were exposed to 15 μM curcumin for 24, 48, and 72 h and then harvested and resuspended in Annexin V binding buffer. The suspension was incubated with 5 μL of Annexin V-FITC and 10 μL of PI for 10 min at room temperature in the dark. The cells were immediately analyzed after staining using a FACScan flow cytometer (Becton Dickinson) and ModFit LT software (Verity Software). For each measurement, at least 20,000 cells were counted.
Measurement of mitochondrial membrane potential (Δψm)
The mitochondrial probe JC-1 (Molecular Probes) was used to detect mitochondrial membrane potential changes. Briefly, after treatment with curcumin, NCI-H446 cells were harvested, washed in PBS, and incubated with JC-1 dye (2.5 μg/mL) at 37°C in a 5% CO2 incubator for 20 min. The cells were then washed, resuspended in PBS, and analyzed by flow cytometry.
The Δψm was also measured by laser confocal scanning microscopy (LSM510/ConfoCor2; Zeiss). The cells were washed with PBS three times and stained with the JC-1 at 2.5 μg/mL for 20 min in the dark at 37°C. Subsequently, the cells were washed with PBS three times and examined using a laser confocal microscope. JC-1 was excited at 490 nm and the emission fluorescence was recorded through a 454–600-nm filter.
Cytochrome c release assay
The release of cytochrome c from mitochondria was assayed as previously described (Hussain et al., 2006). Briefly, the cells were treated with and without curcumin, harvested, and resuspended in hypotonic buffer. The cells were homogenized and cytosolic fractions were isolated by differential centrifugation. Protein from the cytosolic fractions of each sample was analyzed by immunoblotting using an anti-cytochrome c antibody.
Measurement of reactive oxygen species
2′,7′-Dichlorofluorescin diacetate (DCF-DA) is a fluorogenic freely permeable tracer specific for reactive oxygen species (ROS) assessment. It can be deacetylated by intracellular esterase to the nonfluorescent 2′,7′-dichlorofluorescin, which is oxidized by ROS to the fluorescent compound 2′,7′-dichloroflorescein. Thus, the fluorescence intensity of 2′,7′-dichloroflorescein is proportional to the amount of ROS produced by the cells. Approximately 1×106 cells/well of NCI-H446 cells were plated in a six-well plate. After treatment, the cells were incubated with 10 μM 2′,7′-dichlorofluorescein diacetate (Invitrogen) at 37°C for 30 min in the dark. After incubation, the cells were harvested and washed with PBS three times. ROS generation was expressed as mean fluorescence intensity, which was analyzed by flow cytometry. The experiment was done at least in triplicate.
Caspase-3, -8, and -9 activity assay
NCI-H446 cells were treated with 15 μM curcumin for 12, 24, and 48 h. After treatment, the cells were extracted with extraction buffer at 4°C for 15 min. The soluble extracts were collected by centrifugation at 14,000 g and 4°C for 15 min and stored at −80°C until assayed. Ten microliters of cell extract (20 μg of protein) was added to a 100 μL reaction mixture containing either 12 μM Ac-DEVD-pNA, Ac-IETD-pNA, or Ac-LEHD-Pna (Biomol Research Laboratories) as the substrate for caspase-3, caspase-8, and caspase-9, respectively, in a 96-well plate. After incubation at room temperature for 2 h, the amount of caspase-mediated p-nitroaniline–derived substrate cleavage was determined using a microplate reader at 405 nm.
Reverse transcription–polymerase chain reaction assay
Total RNA of each group (n=4) was extracted from the basolateral amygdala using Trizol according to the manufacturer's instructions (Invitrogen), and 1 μg of total RNA was reverse transcribed into cDNA, which was amplified using Manju Van RNA PCR kit (AM Ver. 3.0; Takara Bio). The primers were designed and synthesized by Shenggong Biotech Company according to the serial number from GenBank as shown in Table 1. The reaction was started at 94°C for 2 min, amplified for 35 cycles of 30 s at 94°C, 60 s at 54°C (for Bax), 45 s at 59°C (for Bcl-2), or 30 s at 72°C and ended with a 5 min extension at 72°C. GAPDH mRNA was used as an internal control and coamplified with Bax or Bcl-2 mRNA. The products were visualized by electrophoresis on a 1.5% agarose gel, and the density of each band was analyzed using the Gel Image Analysis System (Tanon 2500R).
Western blot
Total protein from cells was extracted in lysis buffer (Pierce) and quantified using the BCA method. A total of 40 μg of protein was separated using 10%–15% SDS-PAGE and then electrophoretically transferred to a PVDF membrane (Millipore). The membrane was blocked with 5% nonfat milk in TBST solution and incubated at 4°C overnight with primary antibody in blocking solution. After washing three times with TBST, the membrane was incubated at room temperature for 1 h with horseradish peroxidase–conjugated secondary antibody diluted in TBST. Protein bands were visualized using enhanced chemiluminescence (Pierce) and detected using BioImaging Systems (UVP). The relative protein levels were calculated based on GAPDH protein as a loading control.
Statistical analysis
SPSS 13.0 software was used for statistical analysis. Data were summarized as mean±SD. Student's t-tests were used to determine the significant differences between groups. Results were considered to be significant for p-values of <0.05.
Results
Curcumin inhibits the proliferation of NCI-H446 cells in a time- and dose-dependent manner
To establish the optimal conditions for curcumin-induced apoptosis of human small cell lung carcinoma cells in vitro, NCI-H446 cells were cultured in the presence or absence of curcumin (3.75–120 μM) for 24, 48, or 72 h. Curcumin-induced cytotoxicity was both time and dose dependent in NCI-H446 cells. Maximum curcumin-induced cytotoxicity was evident after 72 h exposure to 120 μM curcumin. The IC50 value of curcumin was 15 μM in NCI-H446 cells (Fig. 1).
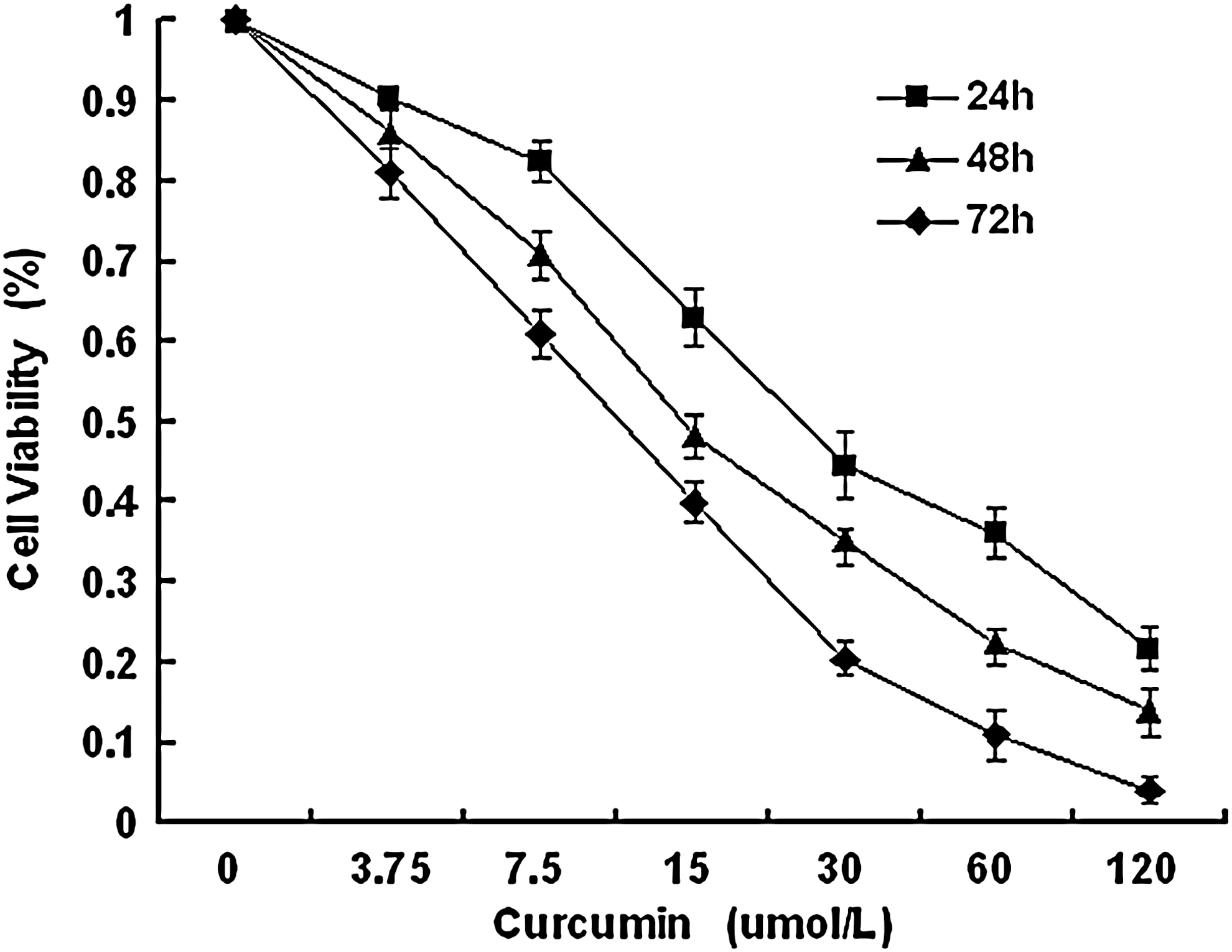
Effect of curcumin on cell viability. NCI-H446 cells were treated with different concentrations of curcumin for different time periods. Proliferation rates were calculated as [(ODtreated/ODcontrol)×100%]. Experiments were performed in triplicate. Values represent mean (±SD) cell viability as a percentage of untreated control samples.
Morphological features of apoptosis
We next examined the cell morphologic characteristics by electron microscopy. Curcumin-treated NCI-H446 cells showed morphologic characteristics of apoptosis, including the disappearance of microvilli, cell shrinkage, a half-moon of condensed chromatin, and membrane blebbing (Fig. 2A).
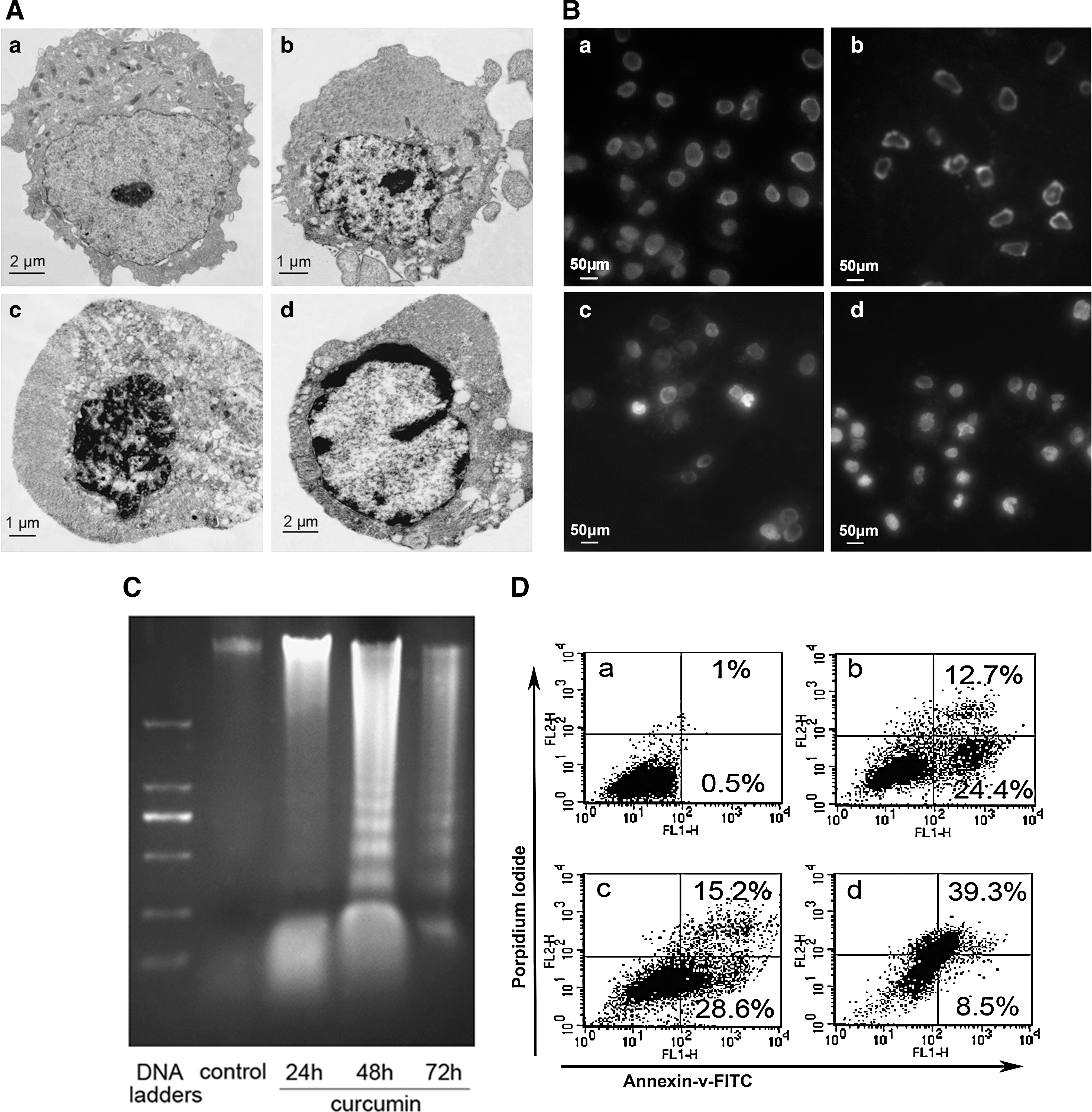
To further observe the morphologic characteristics of apoptosis, NCI-H446 cells were exposed to 7.5, 15, and 30 μM curcumin for 48 h, stained with Hoechst 33342, and analyzed by fluorescence microscopy. Control cells showed an even distribution of the stain and round homogeneous nuclei. Apoptotic cells increased gradually in a dose-dependent manner and displayed typical changes, including reduction of cellular volume, bright staining, and condensed or fragmented nuclei (Fig. 2B).
As shown in Figure 2C, the classic laddering pattern of internucleosomal DNA fragmentation was observed in curcumin-treated cells, indicating that irreversible apoptotic death had been induced.
Curcumin induces NCI-H446 cell apoptosis
We next determined whether curcumin-induced growth inhibition and cytotoxicity of NCI-H446 cells was mediated through the induction of apoptosis by flow cytometry. The percentage of apoptotic cells were analyzed by Annexin V/PI staining. As shown in Figure 2D, curcumin increased the number of apoptotic NCI-H446 cells in a time-dependent manner.
Curcumin stimulated the caspase-3 and caspase-9 activity of NCI-H446 cells
After treatment with 15 μM curcumin for different durations, NCI-H446 cells expressed increased caspase-3 and caspase-9 activity. Caspase-8 activity was not affected by treatment (Fig. 3A). Caspase-9 activity reached its maximum at 12 h, and caspase-3 activity reached its maximum at 24 h. Next, we assessed the effect of curcumin on cleavage of caspase-3, caspase-8, and caspase-9 by western blotting. As shown in Figure 3B and C, the expression levels of cleaved caspase-9 and cleaved caspase-3 were also enhanced with increased curcumin exposure time.
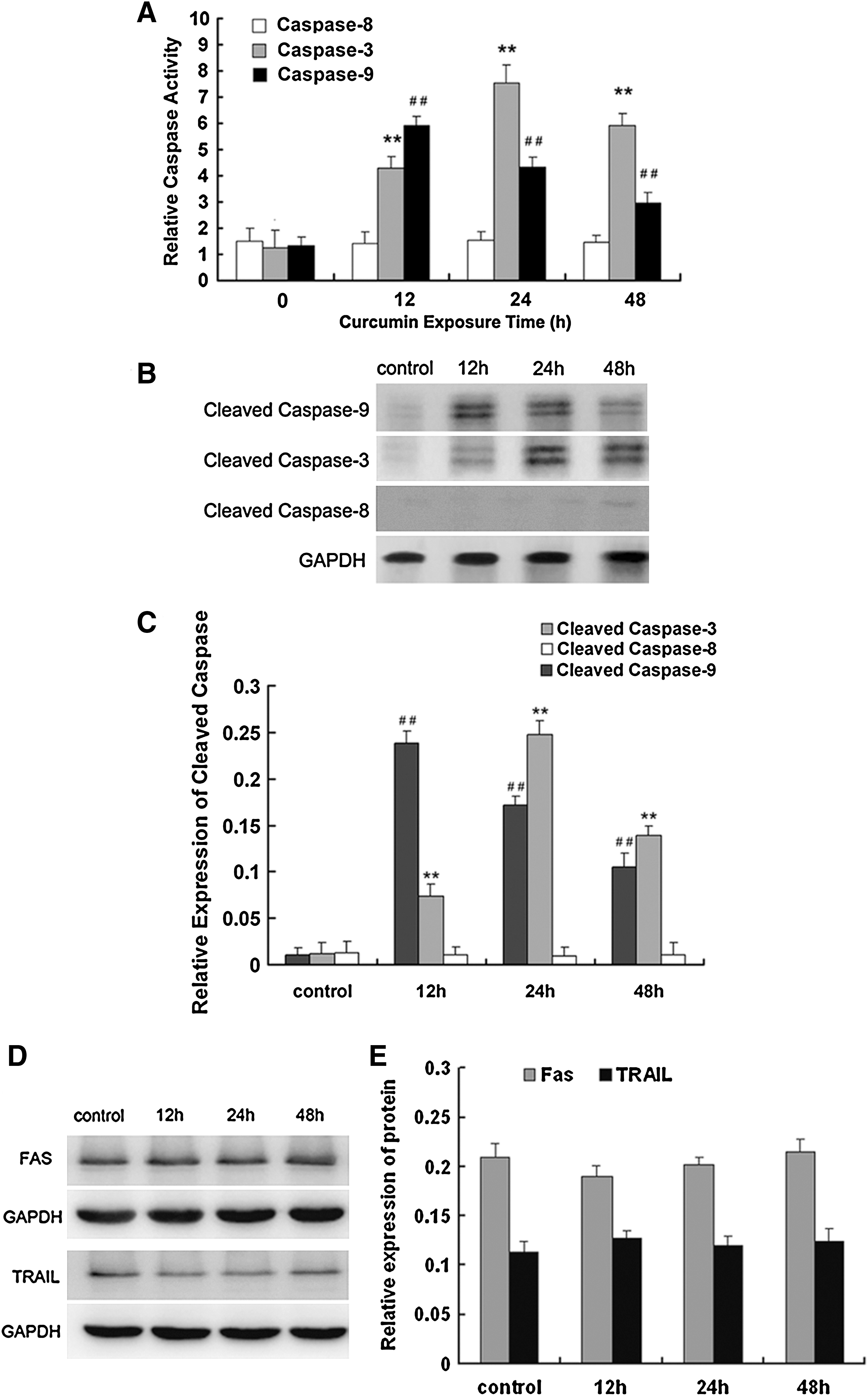
Effect of curcumin on extrinsic apoptotic pathways
Because curcumin treatment did not cause activation of caspase-8, which is an initiator caspase in the extrinsic apoptosis pathway (Walczak and Krammer, 2000), we sought to further determine whether curcumin-induced apoptosis is associated with the death receptor pathway. Curcumin (15 μM) did not cause caspase-8 cleavage (Fig. 3C, D). Under the same experimental conditions, curcumin did not markedly affect the expression of the death receptors FAS and TRAIL (Fig. 3E). These results suggest that curcumin-induced apoptosis may not be mediated by the FAS and TRIAL apoptotic pathway.
Curcumin strongly induces loss of mitochondrial membrane potential and release of cytochrome c
The intrinsic apoptotic pathway involves mitochondrial dysfunction caused by mitochondrial membrane permeabilization due to a loss of membrane potential. The mitochondrial probe JC-1 was used to detect Δψm changes. This fluorescent compound exists as a green fluorescent monomer at low concentrations and as red fluorescent aggregates at high concentrations. Mitochondria with an intact membrane potential concentrate JC-1 into aggregates, whereas deenergized mitochondria cannot concentrate JC-1 and fluoresce green. Curcumin induced loss of Δψm in a concentration- and time-dependent manner. The percentage of cells with a loss of mitochondrial membrane potential increased from 6.2% in control cells to 42.4%–68.8% after treatment with various concentrations for 24 h (Fig. 4A, C). After treatment with 15 μM curcumin for 6 h, the percentage of cells with loss of mitochondrial membrane potential had increased to 5.2% and continually increased to reach 67.3% by 24 h (Fig. 4B, D)
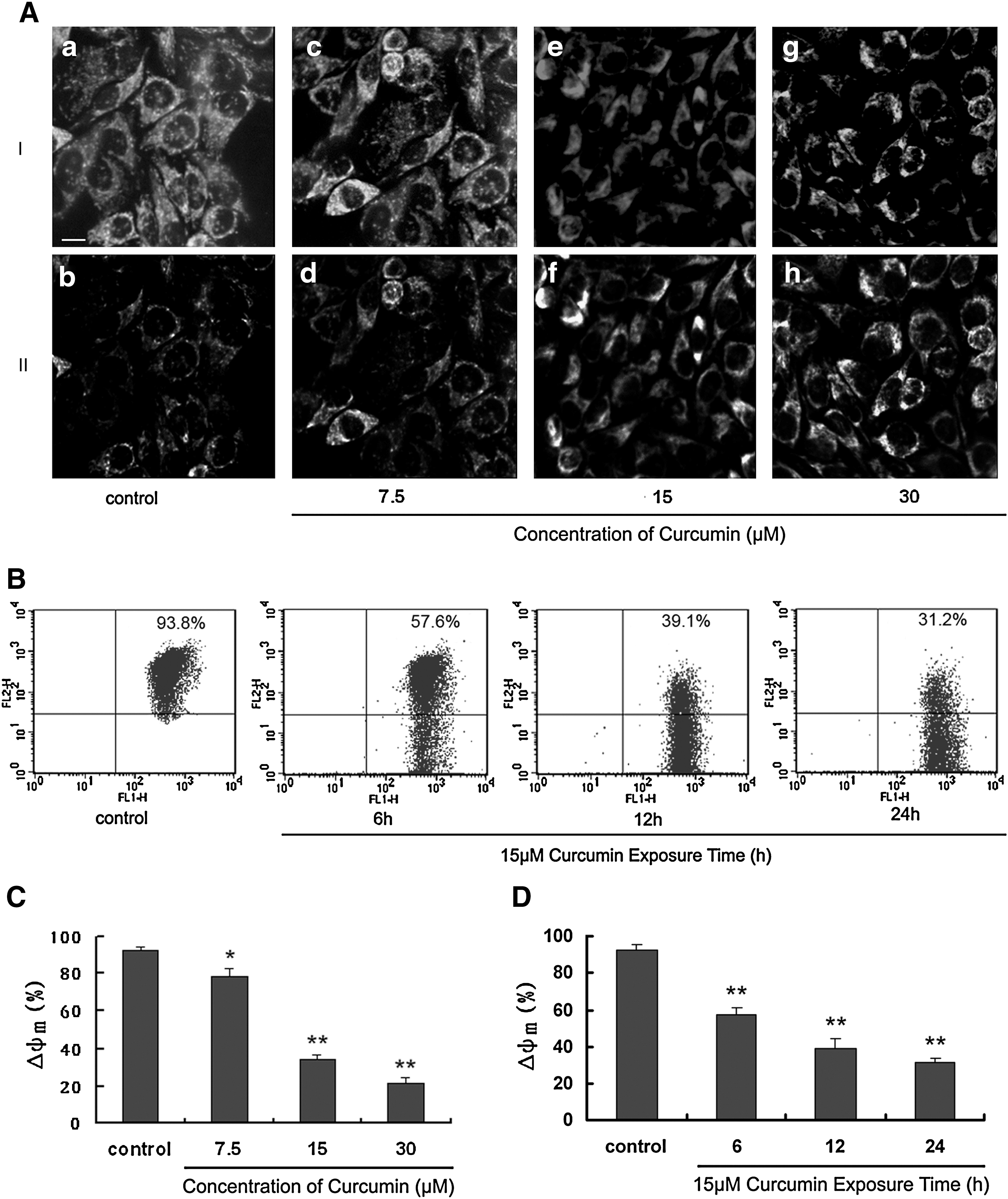
Time- and dose-dependent effect of curcumin on mitochondrial membrane potential.
Cytochrome c release is a key event during mitochondria-dependent apoptosis. Therefore, we examined whether treatment with curcumin enhanced the release of cytochrome c from mitochondria into the cytosol by western blotting. After treatment with 15 μM curcumin, the induced release of cytochrome c was evident at 6 h and further increased at 12–48 h (Fig. 5). Taken together, these results suggest that the mitochondrial pathway plays an important role in curcumin-induced apoptosis.
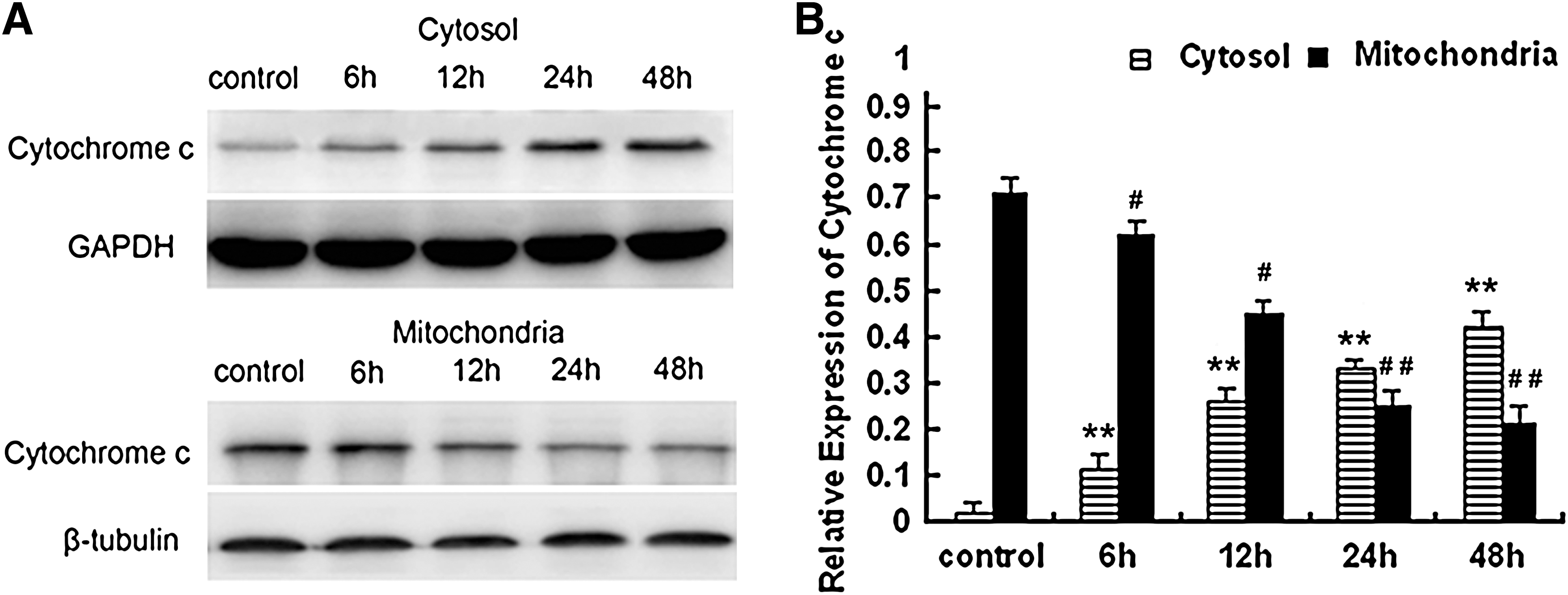
Time response of curcumin on cytochrome c release. NCI-H446 cells were treated with 15 μM curcumin for different time periods as indicated. After treatment, cytosolic and mitochondrial fractions were prepared as described above.
Curcumin induced the production of ROS in NCI-H446 cells
Several reports have demonstrated that curcumin could induce production of ROS in cancer cells (Bhaumik et al., 1996; Atsumi et al., 2006; Hail, 2008; Sánchez et al., 2010). We sought to investigate whether curcumin-induced NCI-H446 cell apoptosis could be tied to ROS generation. We examined the intracellular ROS level by flow cytometric analysis after incubation with DCF-DA in cells treated with curcumin. Compared with the control group, the cells being stained for ROS were significantly increased after treatment with 15 μM curcumin (p<0.05); up to 120 min the levels of ROS were highest and then at 240 min ROS levels were decreased (Fig. 6). These results demonstrated that curcumin increases intracellular ROS levels in human SCLC cells.
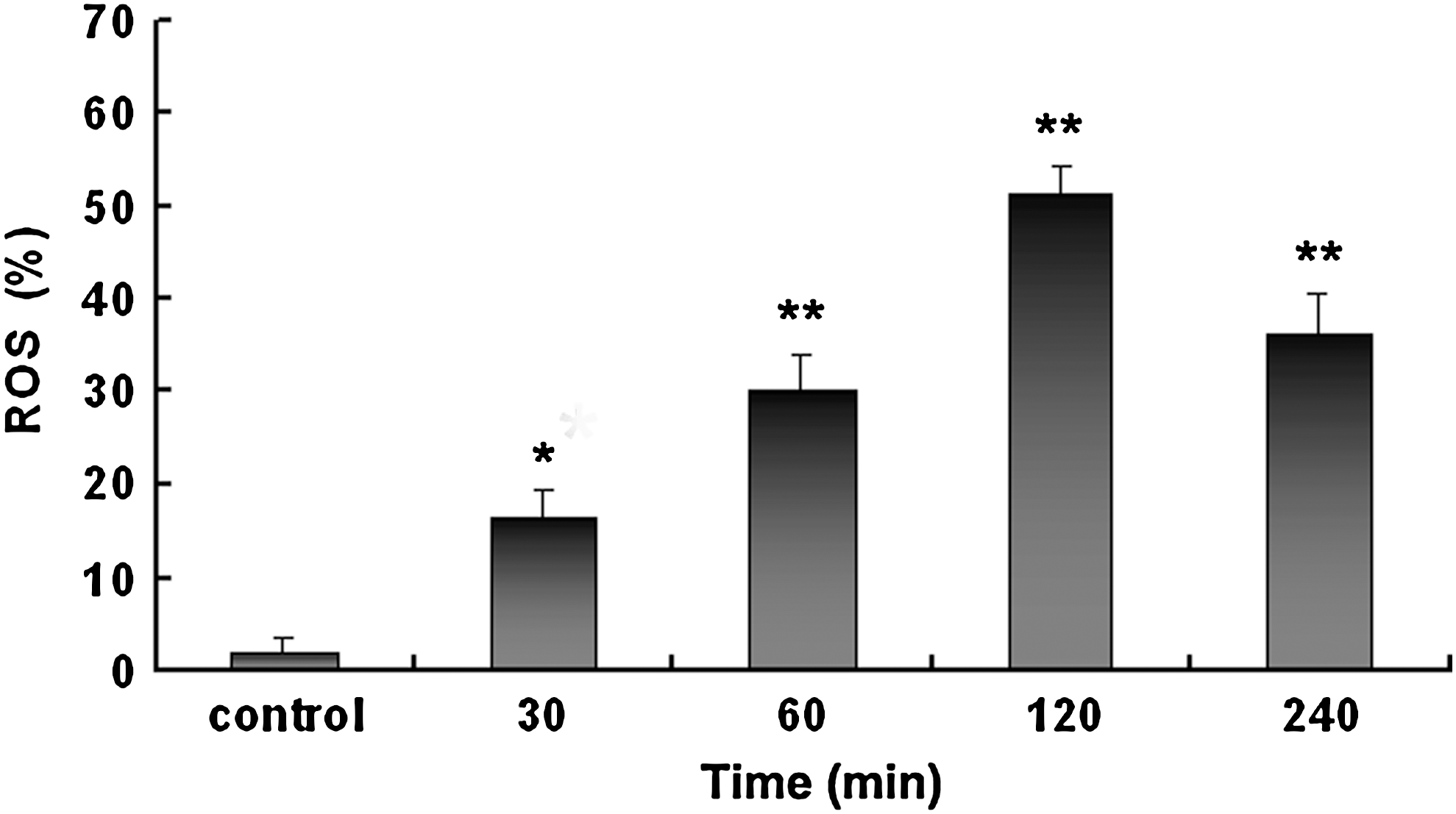
Flow cytometric analysis of reactive oxygen species (ROS) levels in small cell lung cancer NCI-H446 cells with or without curcumin treatment. The NCI-H446 cells were treated with 15 μM curcumin for 30, 60, 120, and 240 min to detect the changes of ROS. *p<0.05 and **p<0.01 versus control.
Effect of curcumin on the mRNA expression of Bcl-2 and Bax
To study the effects of curcumin on Bcl-2 and Bax production at the transcriptional level, NCI-H446 cells were treated with 15 μM curcumin for different times, total RNA was isolated, and reverse transcription–polymerase chain reaction (PCR) was performed. PCR products were visualized by ultraviolet transillumination and showed an increase in the expression of Bax and significant decrease in the expression of Bcl-2 (Fig. 7). This suggests that curcumin reduced the ratio of Bcl-2/Bax at the transcriptional level.
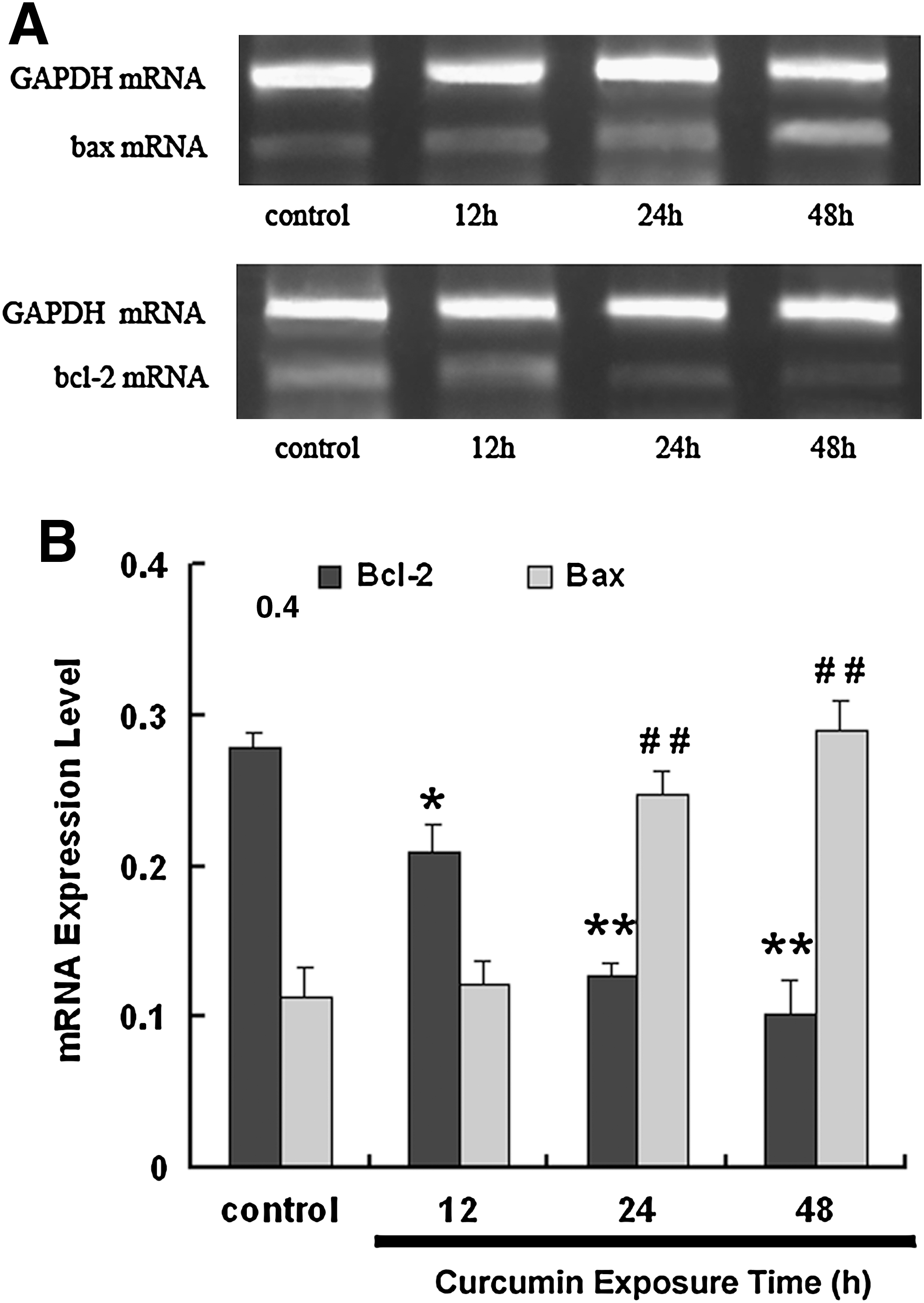
Effect of curcumin treatment on mRNA expression of Bcl-2 and Bax in NCI-H446 cells.
Effects of curcumin on apoptosis-associated proteins in NCI-H446 cells
To characterize the molecular mechanism of curcumin-induced apoptosis in NCI-H446 cells, we examined the expression of apoptosis-associated proteins during treatment with curcumin by western blotting. In the mitochondrial pathway of apoptosis, a number of signals can cause changes in mitochondrial membrane potential and mitochondrial permeability, resulting in the release of cytochrome c. Therefore, we studied the role of curcumin on the expression of proteins involved in the mitochondrial pathway. After treatment with 15 μM curcumin, the antiapoptotic proteins Bcl-2 and Bcl-xL were downregulated in a time-dependent manner, and the proapoptotic protein Bax was upregulated. Therefore, the ratio of Bcl-2/Bax was significantly decreased. Survivin, a member of the inhibitors of apoptosis protein family, is a recently discovered protein that plays a key role in regulation of apoptosis and cell division. Survivin expression is significantly increased in colorectal cancer (Tan et al., 2005). Curcumin treatment dramatically reduced the level of survivin in NCI-H446 cells (Fig. 8).
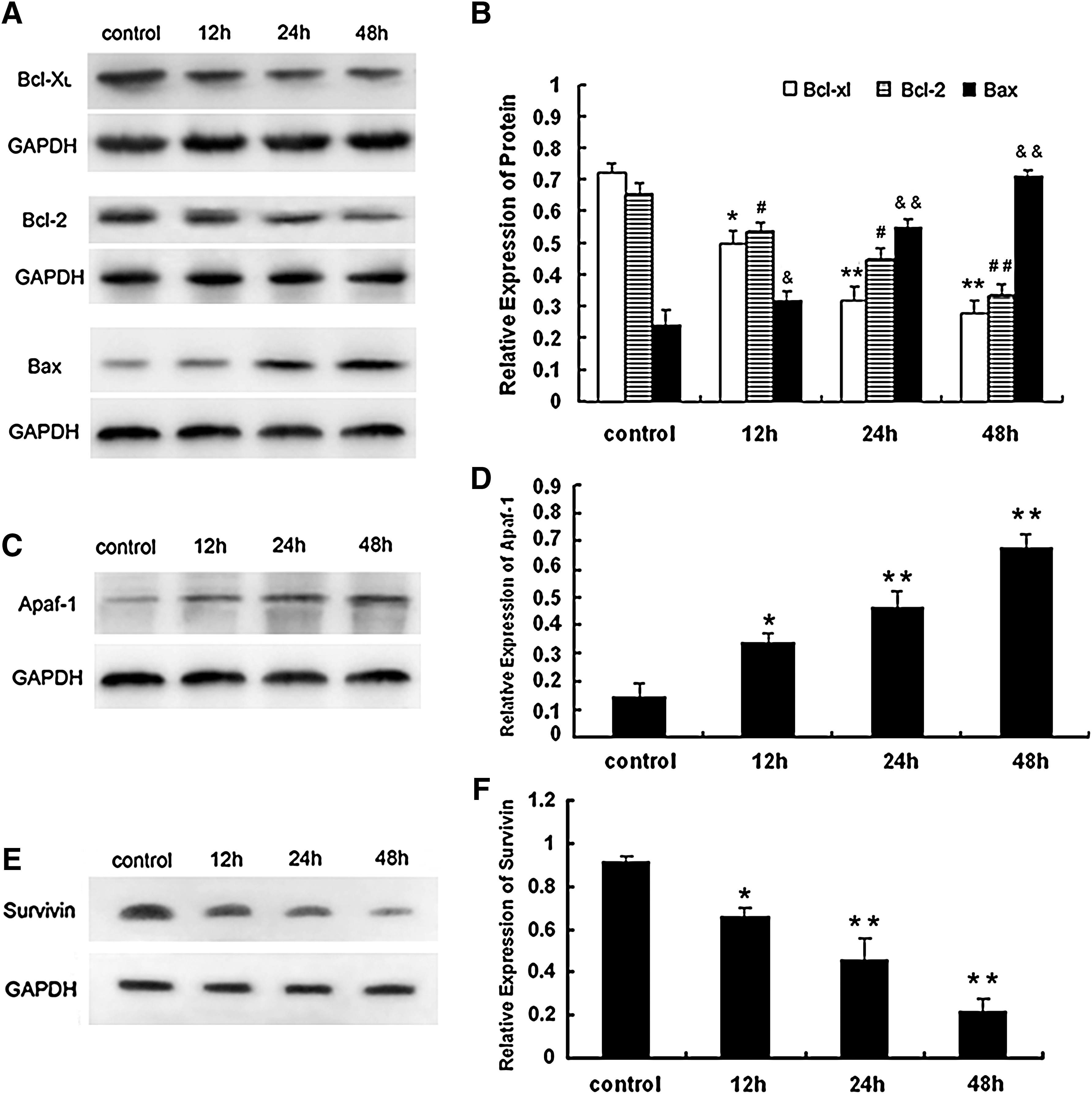
Effect of curcumin treatment on protein expression of
Discussion
SCLC represents only 12%–18% of lung neoplasms and has a slightly declining incidence rate; however, it remains one of the most frequently diagnosed cancers in the United States (Govindan et al., 2006). Compared with NSCLC, it typically presents as a more aggressive and earlier-metastasizing disease that is highly refractory to surgical and radiotherapeutic approaches. This leaves chemotherapy as the only therapeutic option. Therefore, the search for new drugs that can evade drug resistance and adverse effects is very important clinically.
Curcumin possesses anticancer effects on several different forms of human cancer cells. However, the mechanism by which curcumin exerts its anticancer effect needs further research. Despite evidence of curcumin's potential as an anticancer cytotoxic agent, we have little understanding of the mechanism, which involves different effects on different cell types. The effects of curcumin are biphasic; it can promote wound healing, but also be cytotoxic. Either it has multiple targets that are expressed in a cell-specific manner or a target that affects cell-specific pathways. We found that curcumin caused growth inhibition and cell death of NCI-H446 cells in a time- and dose-dependent manner. Our main objective was to determine whether apoptosis was involved in curcumin-induced cell death. We quantified cell apoptosis by flow cytometry, transmission electron microscopy, Hoechst DNA staining, and DNA laddering. As the concentration of curcumin and incubation time increased, the proportion of apoptotic cells also increased. Apoptosis is characterized by a series of morphological changes involving cell shrinkage, chromatin condensation, and the formation of apoptotic bodies (Borner, 2003), which were observed in curcumin-treated NCI-H446 cells. Therefore, curcumin inhibits SCLC cell proliferation mainly by inducing apoptosis. Our results are consistent with other reports of curcumin-mediated cytotoxic activity via the induction of apoptosis (Pongrakhananon et al., 2010; Yallapu et al., 2010). Further, we found that curcumin did not activate caspase-8 or affect the expression of the apoptotic receptors FAS and TRAIL. Thus, apoptotic receptor-related pathways do not mediate curcumin-induced apoptosis. SCLC cells frequently lose both the Fas receptor and caspase-8 because of promoter DNA methylation and are highly resistant to the death receptor pathway (Shivapurkar et al., 2002; Hopkins-Donaldson et al., 2003). In addition, the translocation of apoptosis-inducing factor into the nucleus, which represents a caspase-independent pathway, is less pronounced in SCLC cells than in NSCLC cells after treatment with staurosporine (Joseph et al., 2002). Thus, the apoptotic pathway induced by curcumin may vary according to the type of malignancy. Our data suggest that curcumin-induced growth inhibition and cytotoxicity in NCI-H446 cells is mediated through the mitochondrial apoptotic pathway.
Mitochondria play a crucial role in the apoptotic process by integrating numerous apoptotic signals emanating from the extracellular environment (e.g., toxins, death receptors ligands, anoxia, radiation, and growth factors) and from the intracellular milieu and other organelles (e.g., calcium, ROS, Bax, p53, and caspases). In addition, mitochondria coordinate the caspase-dependent and -independent degradation steps of apoptosis (Brenner and Kroemer, 2000). Mitochondrial membrane potential (MMP) is a point of no return in the apoptosis signaling cascade. As a consequence, MMP modulation has been proposed as a therapeutic goal, and the development of robust cellular and subcellular assays to identify biomarkers, therapeutic targets, and novel therapeutic drugs is currently underway (Bouchier-Hayes et al., 2005; Armstrong, 2006). Mitochondrial dysfunction, including the loss of mitochondrial membrane potential (Δψm), permeability transition, and release of cytochrome c into the cytosol, is associated with apoptosis (Blattner et al., 2001). Opening of permeability transition pores in the mitochondrial inner membrane leads to mitochondrial swelling, membrane depolarization, and release of intramitochondrial solutes (Zamzami et al., 1995). Intracellular ROS function as trigger or signaling molecules to initiate downstream events in regulating cell cycle, cell differentiation and apoptosis. Dysfunction of the mitochondria, which is a hallmark of programmed cell death, likely produces ROS, which influence numerous cell processes (Fleury et al., 2002). Our results showed that curcumin induced ROS production, decreased the levels of Δψm, and promoted the cytochrome c release from mitochondria followed by activation of caspase-3 prior to apoptosis. Cytochrome c combines with an apoptotic protease-activating factor-1 (Apaf-1) and induces the formation of a complex known as the apoptosome. The apoptosome possesses a caspase recruitment domain, which allows it to bind and process the crucial initiator, caspase-9. Activated caspase-9 then cleaves and activates the downstream effector, caspase-3. Our data suggest that treatment with curcumin promotes the release of cytochrome c, which activates caspase-9 and caspase-3, inducing apoptosis through the mitochondrial apoptotic pathway.
Mitochondrial membrane permeabilization is a central process in programmed cell death pathways and is regulated by Bcl-2 family members via multiple molecular mechanisms (Wei et al., 2001). Bcl-2 family members are grouped into the following three classes: one class that inhibits apoptosis, such as Bcl-2; a second class that promotes apoptosis, such as Bax; and a third divergent class of BH3-only proteins, such as Bad (Cory and Adams, 2002). Although Bcl-2 can prevent cytochrome c release and apoptosis, Bax triggers cytochrome c release in cells. Bcl-2 and Bcl-xL are frequently overexpressed in SCLC tumors (Jiang et al., 1996; Reeve et al., 1996; Leech et al., 2000). A meta-analysis of lung cancer found that 76% and 35% of patients' samples are positive for Bcl-2 in SCLC and NSCLC, respectively (Martin et al., 2003). Overexpression of antiapoptotic Bcl-2 family members is known to cause resistance to apoptosis and therapy in a wide range of tumors, and the development of therapies that target these apoptosis modulators appears to be a promising approach (Pellecchia and Reed, 2004; Chonghaile and Letai, 2008). The transcription factor nuclear factor-kappa B (NF-κB) has been reported to be constitutively active in many types of cancer. Many studies have shown that curcumin can inhibit the activity of NF-κB and thereby enhance apoptosis (Aggarwal et al., 2004; Li et al., 2004; Marín et al., 2007). Curcumin treatment could greatly inhibit the expression of Bcl-2 and survivin (as shown in Fig. 8), which are two key target genes in the NF-κB mediated apoptosis pathway. These changes may be associated with NF-kB, which is constitutively active in NCI-H446 cells and may be suppressed by curcumin treatment. Here, we showed that curcumin induced caspase-dependent apoptosis through the mitochondrial pathway, accompanied by decreased Bcl-2 and Bcl-xL expression in SCLC cell lines.
In conclusion, our results demonstrate that curcumin-induced apoptosis in NCI-H446 cells is associated with activation of caspases at initiative and executive stages. Curcumin-induced apoptosis is dependent on the ROS-mediated mitochondrial pathway and is independent of the cell death receptor pathway. These findings provide a better mechanistic understanding of curcumin-induced apoptosis and provide rationale for the use of curcumin in clinical studies of SCLC.
Footnotes
Acknowledgment
The authors thank Chinese Academy of Medical Sciences for providing NCI-H446 cells.
Disclosure Statement
No competing financial interests exist.