
Abstract
This study evaluated the immune response elicited by a ubiquitin (Ub)-fused MPT64 DNA vaccine against Mycobacterium tuberculosis. BALB/c mice were vaccinated with plasmid DNA encoding MPT64 protein, Ub-fused MPT64 DNA vaccine (UbGR-MPT64), and negative DNA vaccines, respectively. MPT64 DNA vaccine immunization induced a Thl-polarized immune response. The production of Thl-type cytokine (interferon-gamma [IFN-γ]) and proliferative T cell responses were enhanced significantly in mice immunized with UbGR-MPT64 fusion DNA vaccine, compared with nonfusion DNA vaccine. Moreover, this fusion DNA vaccine also resulted in an increased relative ratio of IgG2a to IgGl and the cytotoxicity of T cells. IFN-γ intracellular staining of splenocytes indicated that UbGR-mpt64 fusion DNA vaccine activated CD4+ and CD8+ T cells, particularly CD8+ T cells. Thus, this study demonstrated that the UbGR-MPT64 fusion DNA vaccine inoculation could improve antigen-specific cellular immune responses, which is helpful for protection against TB.
Introduction
To develop new vaccines, full understanding of the protection mechanism against TB is required. The crucial factor of protective immunity against TB is a T-cell-mediated response characterized by the secretion of interferon-gamma (IFN-γ) and other cytokines (Cardona et al., 1999). Hence, new vaccines that can provoke potent protective cellular immunity are urgently needed. DNA vaccine is promising compared with conventional vaccines and can induce Th1-type response. DNA vaccines have been studied against tuberculosis in animal models (Ulmer et al., 1997; Zhu et al., 1997; Tanghe et al., 1999; Okada, 2006; Palma et al., 2007; Tang, et al., 2007; Zhang et al., 2007). These DNA vaccines encoding Ag85A/B/C, ESAT-6, MPT64, PST1/2/3, HSP65, 38 kDa, or HSP70, when used individually or in combination, have conferred inferior or similar protection against M. tuberculosis challenge as BCG. Hence, the efficacy of DNA vaccines against TB needs improvement.
The protein MPT64, which is restricted to M. tuberculosis, virulent Mycobacterium bovis strains, and a small number of strains of BCG, is recognized by the immune systems of the majority of TB patients and their contacts. Hence, MPT64 is a promising candidate antigen. In experimental mouse models, MPT64 DNA vaccine conferred protective efficacy inferior to that of BCG vaccine (Arun et al., 1999). Thus, the efficacy of MPT64 DNA vaccine requires improvement. The ubiquitin (Ub)-proteasome system plays a key role in antigen presentation through the major histocompat complex (MHC) class I pathway (Varshavsky, 1996). When a protein is fused to Ub, the degradation of the protein in the proteasome and presentation can be enhanced, resulting in an improvement of immune response. In this study, we demonstrated that UbGR-MPT64 fusion DNA vaccine was capable of improving the cellular immune response against MPT64.
Materials and Methods
Mice
BALB/c female mice, 6–8 weeks old, were bred in the animal facilities of the Second Military Medical University (SMMU). All procedures performed on animals were conducted according to the guidelines for the care and use of laboratory animals of SMMU under protocols approved by the institutional Animal Care and Use committee at the SMMU.
Cell transfection
The recombinant plasmid pcDNA3-MPT64 was transfected into P815 (H-2d a lymphoma cell line, from Type Culture Collection of Chinese Academy of Sciences) cells by liposome (Roche Molecular Biochemicals) according to the manufacturer's instruction. After selection in medium supplemented with G418 (800 μg/mL), stable transfectants were subcloned by limiting dilution and then determined by reverse transcription–polymerase chain reaction (RT-PCR) and immunochemistry methods.
Immunocytochemistry
Expression of MPT64 protein was detected by immunocytochemistry. P815 stable transfectants were fixed in 4% paraformaldehyde for 10 min and placed on poly-l-lysine-treated microslides, and were then air-dried for 30 min. Slides were redehydrated and blocked using 1% bovine serum albumin (BSA) in phosphate-buffered saline (PBS) plus 0.1% Triton X-100 (pH 7.2) for 1 h. Then, slides were incubated overnight at 4°C in a humid chamber with appropriate sera diluted at 1:20 in PBS from the patients infected with M. tuberculosis (provided by Dr. Xiao An with the permission of patients). After washing in PBS (three times for 10 min), the bound human immunoglobulin was detected by incubation for 24 h at 4°C with goat anti-human-horseradish peroxidase (HRP)-conjugated second antibody (Southern Biotechnology Associates, SBA) diluted 1:100 in PBS plus 1% goat serum. After washing in PBS (three times for 10 min), the target antigen was colored by Diaminobenzidine (DAB) substrate and the slides were counterstained with hematoxylin.
Plasmid construction and preparation
For the construction of pcDNA3-MPT64, the cDNA of MPT64 was inserted into the HindIII and XbaI restriction sites of the pcDNA3 plasmid (Invitrogen), downstream of the CMV early promoter(Qingmin et al., 2003). For construction of Ub-MPT64 fusion DNA vaccine, the cDNA encoding Ub with HindIII and BamHI restriction sites was obtained from mouse testicle by RT-PCR. An arginine (R) was added to the C-terminal residues of Ub. The cDNA of MPT64 antigen with BamHI and XbaI restriction sites was also obtained by PCR, not including the starting condon. The spacer sequence (GGGGS) was added between the Ub and MPT64 antigen. Plasmids used in this study were prepared with alkaline lysis method followed by TritonX-114 treatment to remove endotoxin (Cotten et al., 1994).
Vaccination protocol
For DNA vaccination, mice were injected with pcDNA3-MPT64 or pcDNA3-ub-MPT64 (UbGR-MPT64) into both quadriceps with 2×50 μg DNA three times at 3-week intervals. Mice inoculated with pcDNA3 plasmid or pcDNA3-ub were used as negative controls. To enhance muscle cells uptake of plasmid DNA (Danko et al., 1994), 25% sucrose was injected into the muscles of both quadriceps 15 min before plasmid inoculation.
Enzyme-linked immunoabsorbent assay
Anti-MPT64 IgG, IgG1, and IgG2a were measured by enzyme-linked immunoabsorbent assay (ELISA) in individual serum samples from vaccinated mice. The method was as described previously (Wang et al., 2003), using recombinant MPT64 protein (1 μg per well) (Wang et al., 2004) and anti-mouse IgG, IgG1, or IgG2a coupled to HRP (Southern Biotechnology Associates, SBA). The antibody titers were determined according to the optical density (OD450 nm). Finally, the relative ratio of IgG2a to IgG1 was calculated.
Lymphocytes proliferation assay
Mice were sacrificed 3 weeks after the last immunization. Spleens from each group were pooled and analyzed. Th-cell proliferation assay was performed as previously described (Wang et al., 2003). Briefly, the isolated spleen cells were resuspended to a concentration of 5×106 cells/mL. A volume of 100 μL of cell suspension was added to 96-well plates and the MPT64 protein (Wang et al., 2004) was added to the wells in triplicate at the final concentration of 5 μg/mL. The plates were incubated at 37°C in an atmosphere of 5% CO2 for 66 h. Then, the proliferation responses were detected by MTT [3-(4, 5-dimethylthiazol-2-yl) 2, 5-diphenyltetrazolium bromide] (5 mg/mL; Sigma) method and the stimulation index (SI) was calculated. The SI was determined from the formula: SI=experimental OD/negative OD. To assure that cells were healthy, 10 μg/mL ConA was used as a polyclonal stimulator for positive control.
Evaluation of cytokines production in vitro
Single splenocyte suspension from immunized mice was diluted in 10% bovine calf serum-supplemented RPMI 1640 to 5×106 cells/mL. About 100 μL of cell suspension was added to 96-well plates (Costar) and the MPT64 protein was added to the wells in triplicate at the final concentration of 5 μg/mL. After 72 h of incubation, cell-free supernatants were harvested and were screened for the presence of IFN-γ and interleukin (IL)-4 with an ELISA detection system (Jingmei; Biotech) according to the manufacturer's instructions.
Intracellular IFN-γ measurement using flow cytometry
Splenocytes from vaccinated mice were cultured at 2.5×106/mL in 24-well tissue culture plates (Nunclon) in the presence of 5 μg of MPT64 protein/mL for 3 days. Brefeldin A (Sigma) was added to the cultures for the last 5 h to prevent secretion of the intracellular cytokine. One million cells from each group were first incubated with fluorescein isothiocyanate-conjugated anti-CD4 Ab (clone RM4; 4 PharMingen) or CD8 Ab for 30 min at 4°C. Cells were then washed, fixed with 4% paraformaldehyde, and permeabilized with phosphate buffered saline containing 0.1% saponin. To label intracellular IFN-γ, cells were incubated with phycoerythrin-conjugated anti-IFN-γ Ab (clone XMG1.2; PharMingen) for 1 h at room temperature, washed, and acquired on a cytofluorometer (FACSCALIBUR; BD). Lymphocytes were gated by their forward and side light scattering properties, and 100,000 cells were acquired in the lymphocyte gate. Analysis was performed by Cell Quest software.
Cytotoxicity assay of T cells
Spleen cells adjusted to a concentration of 107/mL from in vivo-primed mice were cocultured with mitomycin (10 μg/mL)-treated target cells (P815-MPT64, 5×105/mL) in a 10 mL cell suspension in RPMI 1640 for 5 days at 37°C in 5% CO2. Twenty units per milliliter recombinant murine IL-2 (Biosource) was also added to the cell solution for 5 days. The P815 cell was used as a negative control.
To measure the specific lysis of the target cells, we used the lactate dehydrogenase (LDH) release assay, which yields results similar to the standard chromium release assay, but does not require the use of radioisotopes. In 96-well round-bottom plates, effector cells were incubated with target cells at different E/T ratio for 4h in phenol red-free RPMI 1640 containing 2% BSA, 2 mM glutamine, and 1% penicillin and streptomycin. After centrifuging the plates at 250 g for 10 min, 100 μL per well of the supernatant was then transferred to 96-well plates, and lysis was determined by measuring LDH release using a cytotocxicity detection kit (Roche Molecular Biochemicals). The released LDH converted the added substrate (tetrazolium salt) into a red formazan product, and the amount of color is proportional to the number of lysed cells. The absorbance values from supernatants were recorded at OD492 nm on an ELISA microplate reader. The percent lysis was calculated as follows: (sample release − spontaneous release)×100%/(total release − spontaneous release)×100%.
Statistical analysis
The statistical significance of differential findings between experimental groups was determined by Student's test. Data were considered statistically significant at p<0.05.
Results
Construction of pcDNA3-MPT64 and UbGR-MPT64 fusion DNA vaccine
The recombinant pcDNA3-MPT64 plasmid was confirmed by HindIII and XbaI digestion (Qingmin et al., 2003). The UbGR-MPT64 fusion DNA vaccine was confirmed, respectively, by HindIII and XbaI, BamHI and XbaI, HindIII, and XbaI digestion. Finally, the sequences of the two DNA vaccine plasmids were confirmed by sequencing. After large-scale preparation, the plasmids were suspended in endotoxin-free PBS. DNA was quantified by spectrophotometry at 260 nm and the final concentration of the solution was adjusted to 1 μg/μL of DNA in PBS.
The stable expression of MPT64 protein in P815 cells
The purpose of transfection experiment was to obtain the specific target cells for cytotoxicity assay. After selection by G418 (800 μg/mL), 15 clones were obtained and 5 clones of transfected cells were randomly chosen and screened for MPT64-mRNA by RT-PCR. After electrophoresis, a specific single band about 0.7 kb in length was observed in clones I, II, III, IV, and V. Expression of MPT64 was further examined in clone I by immunocytochemistry. The immunostaining was restricted to the cytoplasm of the cells transfected with pcDNA3-MPT64 plasmid. However, no staining was detected in P815 cells. No signals were detected with the sera from healthy people, which indicated that the staining is specific. Those results demonstrated that MPT64 antigen could be expressed stably in P815 cells, and that the clone I could be used as target cells in the cytotoxicity assay.
Production of antibodies induced by different vaccines
To determine the level of MPT64-specific IgG elicited by different vaccines, mice of different groups were immunized three times at 3 weeks intervals. Three weeks after the last immunization, the sera from mice were collected by retro-orbital bleeding and antigen-specific antibodies were detected by ELISA. As shown in Figure 1, compared with the pcDNA3 vector group or pcDNA3-ub group, the MPT64 DNA vaccine elicited a significantly higher level of IgG (p<0.01). However, the IgG level in the UbGR-MPT64 fusion DNA vaccine group was lower than that in MPT64 DNA vaccine group (p<0.05).

The MPT64-specific IgG titer in mice immunized by different DNA vaccines. Each group of mice (n=8) was immunized, respectively, by the blank vector, pcDNA3-ub, pcDNA3-MPT64 DNA vaccine, or UbGR-MPT64 DNA vaccine at 0, 3, and 6 weeks. Mice were bled at 3 weeks after the last immunization, and MPT64-specific IgG titer was detected by enzyme-linked immunoabsorbent assay. Optical density was measured at 450 nm. Data shown represented geometric mean titers (GMT) and standard errors for each group of animals. **p<0.01 pcDNA3-MPT64 versus pcDNA3 group or pcDNA3-ub group; *p<0.05 pcDNA3-MPT64 versus UbGR-MPT64 group. The experiment was repeated three times.
The IgG subclasses give an indication of the Th1 versus Th2 nature of the immune response. We also detected the relative ratio of IgG2a/IgG1. As shown in Figure 2, although the IgG level decreased in the Ub fusion DNA vaccine group, the relative ratio of IgG2a/IgG1 increased significantly in the fusion DNA vaccine (p<0.05), compared with the MPT64 DNA vaccine group.
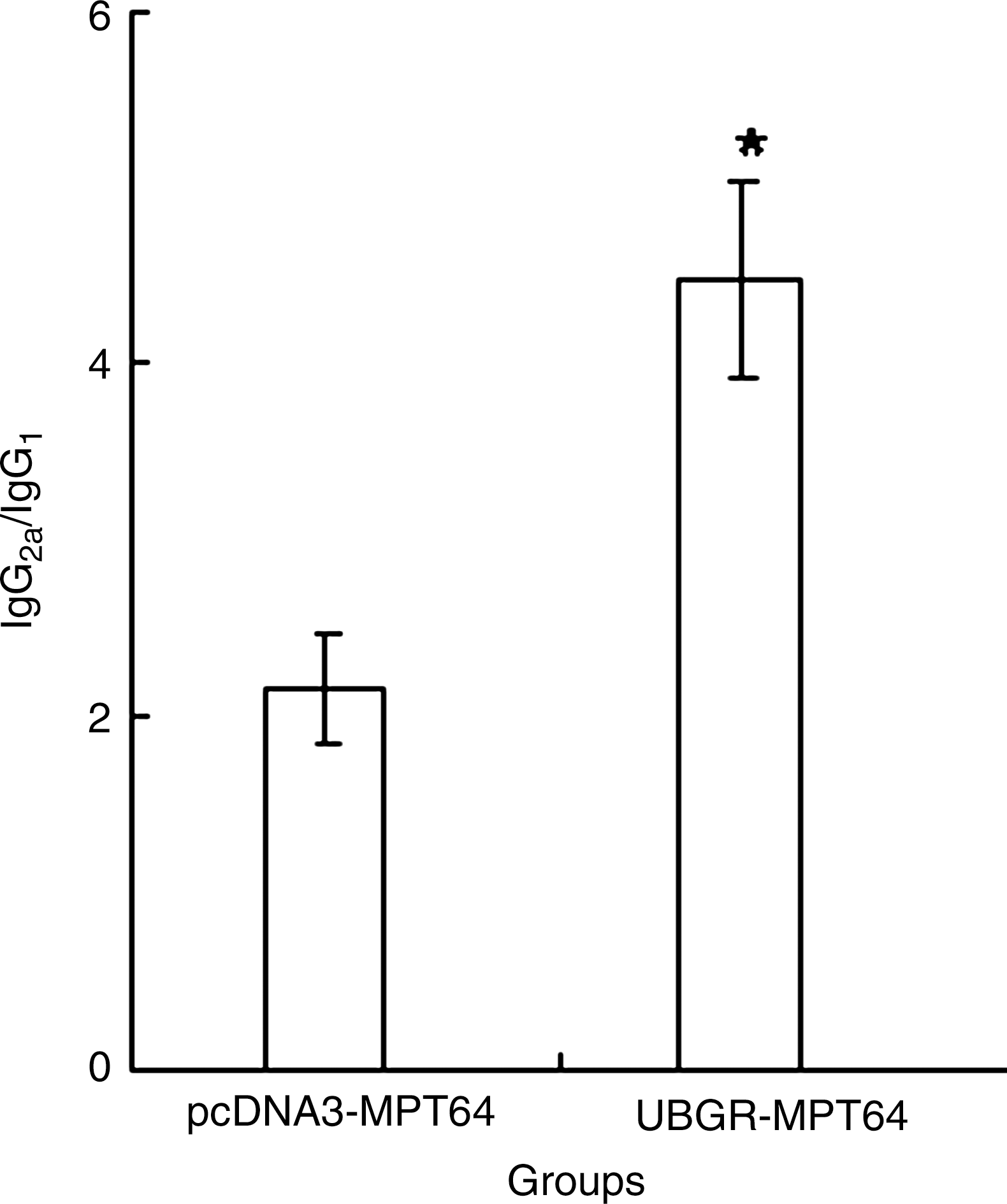
The ratio of IgG2a titer to IgG1 titer in different groups. The mice were immunized as described in Figure 1. The titers of IgG2a and IgG1 were detected, and the ratio of IgG2a/IgG1 was obtained by dividing the titer of IgG2a by that of IgG1. *p<0.05 UbGR-MPT64 group versus pcDNA3-MPT64 group. The experiment was repeated three times.
Lymphocyte proliferation and cytokine production by splenocytes from vaccinated mice
T helper cells play an important role in eliciting both humoral and cellular immune responses via expansion of antigen-stimulated B cells and expansion of CD8+ T cells. Hence, it is important to measure proliferation of T cells after immunization with vaccines when stimulated in vitro with a specific antigen. In our experiment, MPT64 (5 μg/mL) and ConA (10 μg/mL) were used as a specific stimulator and a polyclonal stimulator of T cells, respectively. As shown in Figure 3, a low background level of T-cell proliferation was observed in vector control group and pcDNA3-ub group. A significant increase of T cells proliferation (p<0.01) was observed in pcDNA3-MPT64 group compared with vector group or pcDNA3-ub group. The ubiquitinated MPT64 DNA vaccine significantly enhanced Th-cell proliferation responses compared with nonubiquitinated MPT64 DNA vaccine (p<0.05).
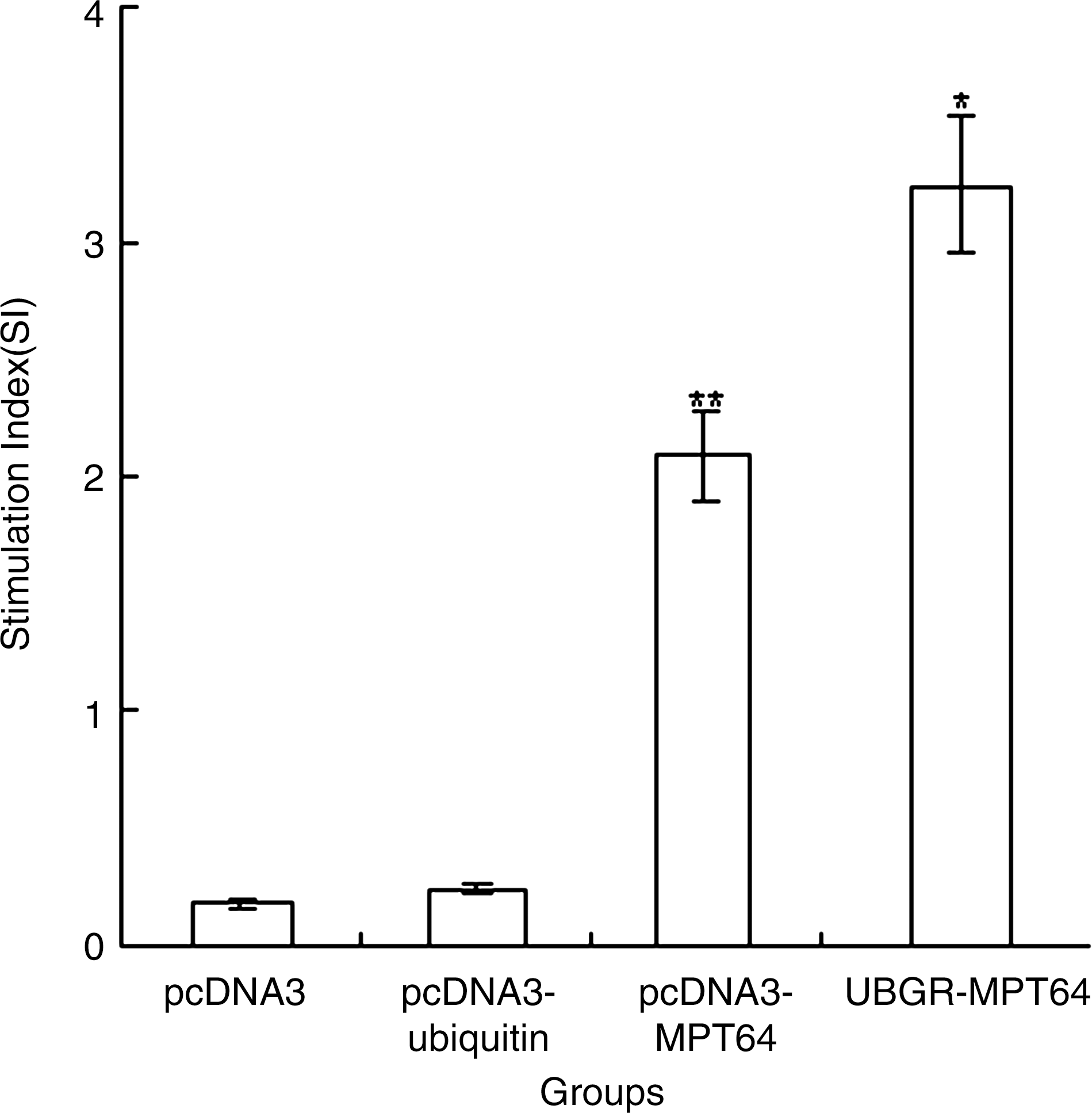
The proliferation of splenocytes after in vitro stimulation with MPT64 protein. Different group of mice (n=8) were, respectively, immunized by pcDNA3 vector, pcDNA3-ub, pcDNA3-MPT64, or UbGR-MPT64 DNA vaccines three times at 3 weeks interval. Three weeks after the last immunization, spleen cells from sacrificed mice were pooled and stimulated with 5 μg/mL MPT64 protein, or 10 μg/mL ConA as a positive control. After 66 h stimulation, samples were assayed in triplicate. The ConA control sample showed a stimulation index of 7.02±1.6. Values and bars represent the mean and S.D. This experiment was repeated, with the expected results. **p<0.01 pcDNA3-MPT64 group versus pcDNA3 group or pcDNA3-ub group; *p<0.05 UbGR-MPT64 group versus pcDNA3-MPT64 group. Eight spleens were pooled and the experiment was repeated three times.
Cytokines were also detected as a specific indicator of CD4+ T-cell activation. Th1 cytokines (IL-2, IFN-γ) and Th2 cytokines (IL-4, IL-5, and IL-10) are major parameters in our understanding of the polarization of immune responses. Th1 immune responses are thought to drive induction of cellular immunity, whereas Th2 immune responses preferentially drive humoral immunity. In this study, the levels of IFN-γ and IL-4 were examined.
As demonstrated in Figure 4, the level of IFN-γ was significantly higher in the MPT64 DNA vaccine group than that in the pcDNA3 group or in pcDNA3-ub group. The secretion of IFN-γ significantly increased in UbGR-MPT64 fusion DNA vaccine group (p<0.01) compared with MPT64 DNA vaccine group. However, the level of IL-4 was lower in the fusion DNA vaccine group than that in the nonfusion vaccine group (p<0.01). In the MPT64 DNA vaccine group, the level of IFN-γ was higher than that of IL-4, which indicated the MPT64 DNA vaccine elicited a Th1-profile immune response. The Ub fusion DNA vaccine increased the secretion of IFN-γ and decreased the level of IL-4, which demonstrated that the Ub fusion enhanced the Th1-type immune response.
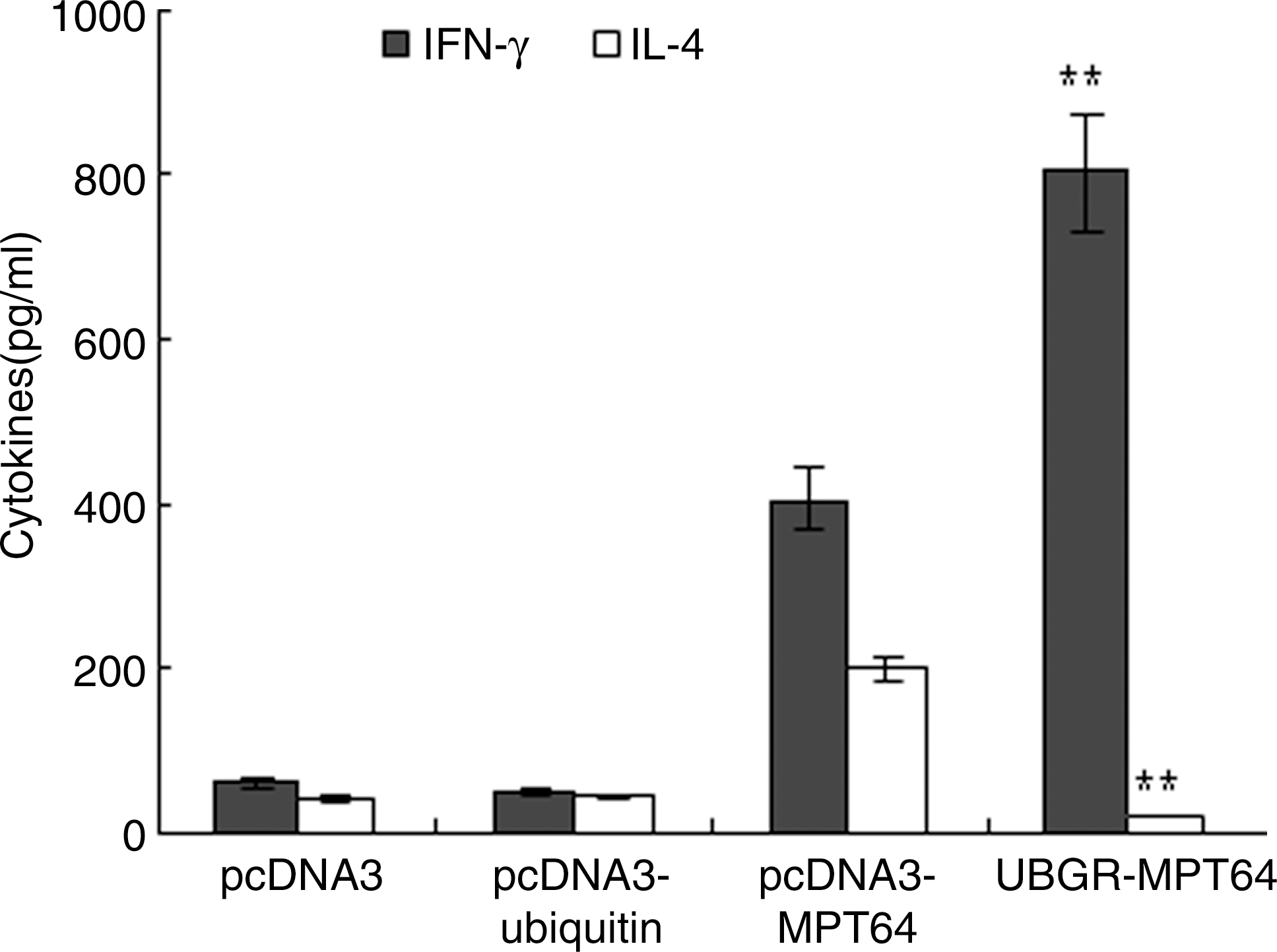
Production of IFN-γ and interleukin-4 from splenocytes of different groups. Different group of mice (n=8) were immunized by the blank vector, pcDNA3-ub, pcDNA3-MPT64 DNA vaccine, or UbGR-MPT64 DNA vaccine at a 3 week interval. Three weeks after the last immunization, spleen cells from sacrificed mice were pooled and stimulated with 5 μg/mL MPT64 protein for 72 h. Samples were assayed in triplicate. Values and bars represent the mean of released cytokine concentrations and S.D. **p<0.01 UbGR-MPT64 group versus pcDNA3-MPT64 group. Eight spleens were pooled and the experiment was repeated three times. IFN-γ, interferon-gamma.
IFN-γ intracellular staining for splenocytes in vaccinated mice
The role of CD4+ and CD8+ T cell in secreting IFN-γ was investigated by intracellular staining since IFN-γ is a key molecule in the anti-tuberculosis response. As shown in Figure 5, the frequency of IFN-γ+ CD4 T cells and IFN-γ+ CD8 T cells was higher in the MPT64 DNA vaccine group than that in the pcDNA3 vector group or in the pcDNA3-ub group. The frequency of IFN-γ+ CD8 T cells was much higher in the spleens of the UbGR-MPT64 fusion DNA vaccine group than those in the MPT64 DNA vaccine group (p<0.01). The frequency of IFN-γ+ CD4 T cells was also higher in the UbGR-MPT64 fusion DNA vaccine group than that in the MPT64 DNA vaccine group although to a lesser extent (p<0.05). Overall, UbGR-MPT64 fusion DNA vaccine induced more antigen-specific CD8+ T cells than CD4+ T cells. These results indicated that UbGR-MPT64 fusion DNA vaccine activated CD4+ and CD8+ T cells, particularly CD8+ T cells.
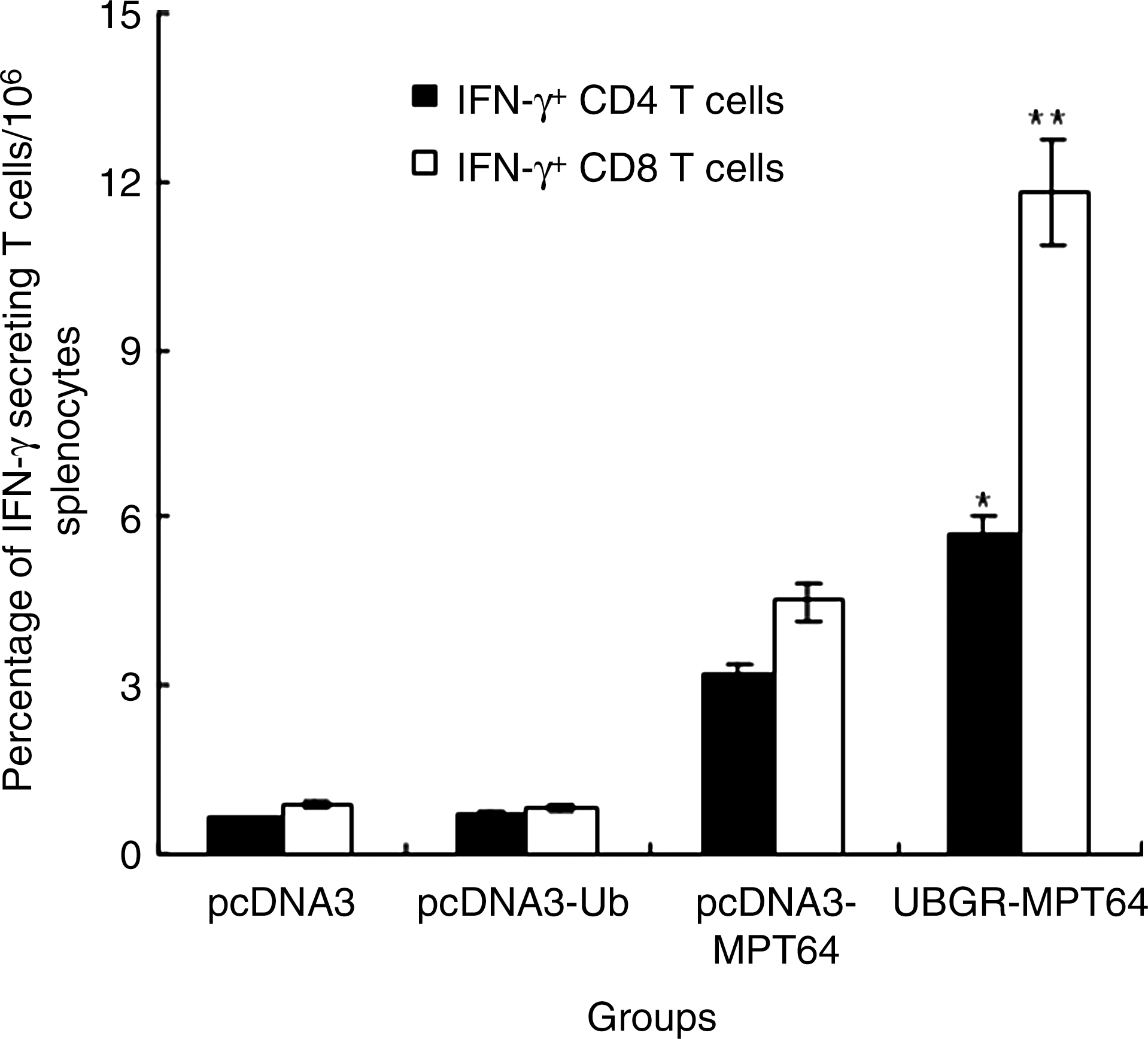
Flow cytometry analysis of MPT64-specific IFN-γ+ CD4 or CD8+ T cells. Mice were inoculated as before (n=8). Three weeks after the last immunization, spleen cells from sacrificed mice were pooled and stimulated with 5 μg/mL MPT64 protein for 72 h. The frequency of IFN-γ+ CD4 or CD8 T cells was detected by flow cytometry. Samples were assayed in triplicate. Values and bars represent the mean of frequency and S.D. **p<0.01 UbGR-MPT64 group versus pcDNA3-MPT64 group; *p<0.05 UbGR-MPT64 group versus pcDNA3-MPT64 group. Eight spleens were pooled and the experiment was repeated three times.
Improved CTL response by UbGR-MPT64 fusion DNA vaccine
Cytotoxic T-cell responses were determined with an LDH release assay, after in vitro restimulation against the target cell line P815-MPT64, which stably expressed the MPT64 protein. P815 cells were used as a negative control. As shown in Figure 6, DNA immunization with MPT64 DNA vaccine resulted in a strong CTL response, which was significantly above the blank vector or pcDNA3-ub immunization (p<0.01). Ub fusion DNA vaccine enhanced the cytotoxic T-cell response, compared with MPT64 DNA inoculation (p<0.05). The blank vector or pcDNA3-ub immunization did not induce CTL response. The spontaneous release was below 10%.
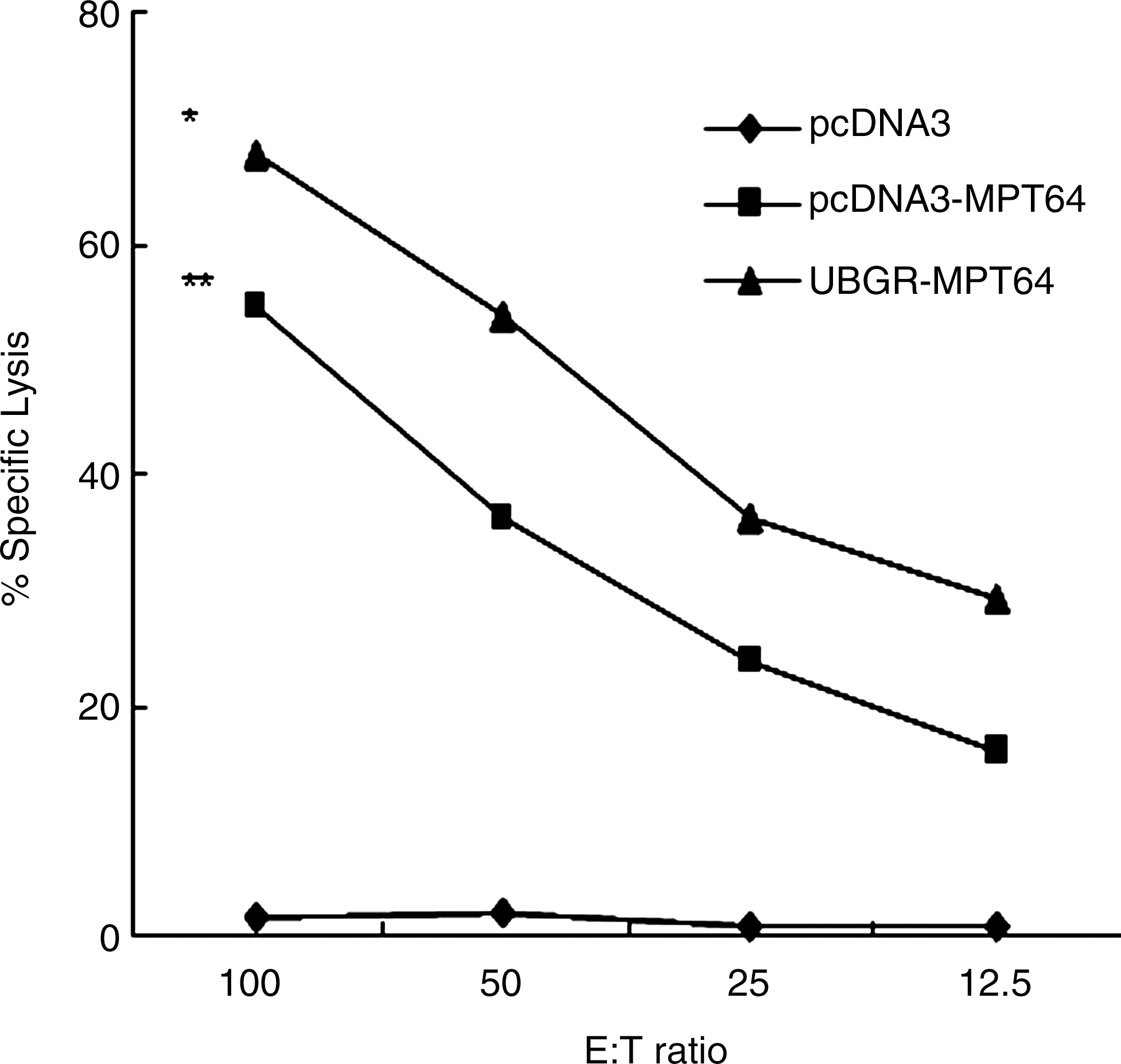
Induction of CD8+ CTL following immunization with different vaccines. Mice were inoculated as before. The percentage of specific lysis was measured after in vitro restimulation of the in vivo-primed cells with mitomycin-treated P815-MPT64 cells. Effector cells [E] were incubated with P815-MPT64 target cells [T] in a lactate dehydrogenase release assay. Samples were assayed in triplicate. Results are representative of three independent experiments. **p<0.01 pcDNA3-MPT64 group versus pcDNA3 group or pcDNA3-ub group; *p<0.05 UbGR-MPT64 group versus pcDNA3-MPT64 group. Eight spleens were pooled and the experiment was repeated three times.
Discussion
DNA vaccines preferentially induce Th1-dominant immune response. The exact mechanism driving Th1 or Th2 type response is not well known, but it has been suggested that CpG motifs from a bacterial plasmid might be responsible for driving immune responses toward Th1-type (Bode et al., 2007). Th1-type response correlates with protective immunity in certain tumor, bacterial, and viral infections, as well as some parasitic diseases. Protective immunity against tuberculosis mainly depends on cellular immune responses and some Th1-type cytokines, such as IFN-γ. Hence, to improve the DNA vaccines against M. tuberculosis, strategies must be explored to enhance the protective immune response. In our study, we chose Ub to modulate the immune response elicited by MPT64 DNA vaccine.
The Ub–proteasome pathway is the main source for intracellular protein turnover. MHC class I most often presents peptides derived from endogenously synthesized proteins, which are degraded by the proteasome. Hence, higher rates of intracellular antigen turnover should increase the number and variety of fragments and peptides available for MHCI binding, which may result in an increase of cell-mediated response to the expressed antigens. To this point, conjugation of the antigen with Ub should target the endogenously synthesized antigens to the proteasome pathway and result in an enhanced cellular immune response. Some researchers have optimized the efficacy of DNA vaccines by increasing antigen degradation (Brandsma et al., 2007; Dobano et al., 2007; Sharma et al., 2009; Chou et al., 2010). There are two methods of fusing Ub to the protein of interest. One is to mutate the C-terminal residue of Ub from glycine (G) to alanine (A), resulting in a stable Ub-protein (UbAAg). This stable Ub-protein can be polyubiquitinated and degraded quickly by the proteasome. The other method is to add an arginine (R) to the C-terminus of Ub, resulting in an unstable Ub-protein (UbGR-Ag). This fusion protein can be quickly recognized and degraded by the Ub system according to the N-rule, also resulting in promoted protein degradation.
Based on the Ub paradigm, we fused UbGR with MPT64 antigen from M. tuberculosis. The change of the immune response elicited by UbGR-MPT64 fusion DNA vaccine indirectly showed the change of MPT64 degradation. Compared with the MPT64 DNA immunization, UbGR-MPT64 fusion DNA vaccine resulted in an lower antibody IgG, enhanced lymphocytes proliferation, a stronger Th1 type immune response, and enhanced cytotoxicity of CTLs. The IFN-γ level in UbGR-MPT64 was higher than that in the MPT64 DNA vaccine group. The increased level of IFN-γ resulted from both CD4+ T and CD8+ T cells, particularly from CD8+ T cells. Interestingly, the ubiquitination strategy designed to improve MHC I-mediated cellular responses also resulted in improved cytokine and proliferative responses mediated by CD4+ T cells. It could be that increasing protein degradation by the proteasome also yields peptides that could be taken up by MHC II molecules. Our result is consistent with Dobaño's report (2007), which showed that immunization with DNA vaccine encoding PyHEP17 fused to Ub induced higher IFN-γ, cytotoxic, and proliferative T cell responses than those of unmodified vaccines. However, no effect was seen for another antigen (PyCSP) using the same targeting strategies. Rodriguez's report (1997) demonstrated that an ubiquitinated DNA vaccine targeted to the protein degradation pathway enhanced cytotoxic T-lymphocyte induction and abrogated antibody induction. However, in Vadlin's study (1999), when Ub was fused with hepatitis C virus (HCV) core antigen, an undetectable antibody response and no increase in CTL activity were observed compared with the nonfusion vaccine. In our study, the humoral immune responses were not completely abrogated. Those different results may correlate with the different antigenicities of the proteins and different dependence of the antigens on Ub.
In conclusion, the data presented above suggested that the fusion of UbGR to our DNA vaccine significantly increased the antigen-specific cellular immune response. Infection with M. tuberculosis is largely intracellular; thus, the protective immune response against M. tuberculosis infection involves a cell-mediated response rather than a humoral response. Both CD4+ and CD8+ T cells and the ability to respond with Th1-type cytokines, particularly IFN-γ, are involved. Taken together, our results demonstrated that the fusion of UbGR to MPT64 DNA vaccine could be a new strategy to improve the efficacy of TB DNA vaccines.
Footnotes
Acknowledgments
We thank Dr. Xiao An for providing us the sera from patients infected with M. tuberculosis. This research was funded by the Natural Science Fund of Shanghai (Number: 08ZR1405600) and the National Natural Science Foundation (Number 31070121).
Disclosure Statement
No competing financial interests exist.