
Abstract
The p53 tumor suppressor gene plays an important role in preventing tumor development. The p53 protein interacts with other p53 signal pathway members to control cell proliferation. In this study, expression of the p53, Human homolog of murine Double Minute 2 (HDM2), p14Alternating Reading Frame (ARF), Zinc Finger and BTB domain containing 7A (ZBTB7A), and B-Cell Lymphoma 6 (BCL6) genes was quantitatively investigated by real-time polymerase chain reaction (PCR) in the peripheral blood of patients with chronic lymphocytic leukemia (CLL) and healthy controls. Plasma fibronectin levels were determined by enzyme-linked immunosorbent assay. Expression of the p53, p14, and HDM2 genes were significantly higher in the patients. However, ZBTB7A and BCL6 gene expression was not detectable in both groups. A positive correlation between p14ARF and HDM2 expression and a negative correlation between p53 and p14ARF expression was observed. Expression of the p14ARF and HDM2 genes were inversely correlated in the control group. Neither HDM2 nor p14ARF gene expression was correlated with p53 expression. The p53 gene was also analyzed for the presence of mutations. A splice-site mutation was found in a single patient. Our findings indicate that expression of the p53, p14ARF, and HDM2 genes are associated with CLL. Elucidation of the mutual interactions at the protein level warrants further studies.
Introduction
CLL is characterized by a highly variable clinical course (Chiorazzi et al., 2005) and treatment is reserved for symptomatic patients or patients with advanced disease. Despite improvements in response rates due to new chemoimmunotherapy modalities, CLL remains incurable, and patients who are refractory to treatment or who experience multiple relapses have a poor prognosis (Keating et al., 2005). Therefore, there is a need for new and reliable prognostic indicators which may enable accurate risk classification, especially for patients with the more aggressive form of the disease.
While staging of the disease is carried out according to the Rai and Binet classifications (Rai et al., 1975; Binet et al., 1981), it is also important to assess the biological properties of the disease. It has been shown that carriers of mutant IgVH survive longer than patients without mutations (Stilgenbauer et al., 2002). CD38 and zeta-chain protein associated tyrosine kinase 70 (ZAP70) are also important biological prognostic factors. CLL patients who do not harbor Ig gene mutations express high amounts of the ZAP70 protein (Rosenwald et al., 2001). Expression of ZAP70 allows IgM signaling in B-cells that may lead to aggressive disease (Hamblin, 2007). High levels of ZAP70 expression lead to chemokine migratory signals in B-cells (Richardson et al., 2006). CD38 expresion in this patient group is associated with poor prognosis (D'Arena et al., 2007).
The p53 gene is activated in a variety of DNA damage or stress situations and decides whether a cell can repair DNA damage at the G1/S transition of the cell cycle or die via apoptosis (Murray-Zmijewski et al., 2008). The activity of the protein product is regulated by the p14ARF and human homolog of murine double minute 2 (HDM2) proteins acting in the p53 pathway (Lane et al., 2004; Mills, 2005). Cellular p53 levels are also regulated by posttranslational modifications like phosphorylation of serine residues and acetylation of the C-terminal lysines (Unger et al., 1999a, 1999b; Chao et al., 2003). Deletions in the p53 gene are observed in 5%–7% of early stage CLL and 25%–40% of advanced disease (Zenz et al., 2008) and are associated with poor prognosis (Xu et al., 2008) as are mutations of the ATM gene (Starostik et al., 1998).
The HDM2 gene codes for an ubiquitin ligase which, besides directly inhibiting p53 gene expression, also suppresses the activity of the p53 protein by binding to its transactivational domain. When p53 levels rise, HDM2 expression is induced and the p53 protein is marked for proteasome-mediated degradation by the covalent attachment of ubiquitin (Haupt et al., 1997). The HDM2 protein is overexpressed in various malignancies (Deb, 2003).
The B-cell lymphoma 6 (BCL6) protein acts as a repressor of p53 gene transcription. It is essential for germinal center development and T-cell-dependent humoral immune response (Ye et al., 1997). The BCL6 protein contains a bric-a-brac tramtrack broad complex (BTB)/Pox virus zinc finger domain and is highly expressed in mature germinal center B-cells (Phan and Dalla-Favera, 2004).
p14ARF is an indirect activator of the p53 protein by antagonizing HDM2 and hence preventing uncontrolled cell proliferation (Kamijo et al., 1998). Expression of p14ARF may increase in response to cellular stress, serum starvation, and action of viral oncogenes. In addition to frequent deletions at the INK4A/ARF locus, methylation of the INK4/ARF gene promoter is also frequent in cancers (Sherr, 2006).
The zinc finger and BTB domain containing 7A (ZBTB7A) protein is a member of the Pox virus, Zinc Finger and Krüppel (POK) transcriptional repressors. POK proteins participate in cellular development, differentiation, and oncogenesis (Ye et al., 1997; Barna et al., 2000; Costoya et al., 2004; He et al., 2005). Inactivation of ZBTB7A in mouse results in incomplete embryonal development and lethality. Cotransfection of ZBTB7A into murine embryo fibroblasts (MEF's) with oncogenes results in transformation. ZBTB7A-null MEF's are resistant to the oncogenic effects of the E1A/RasV12, Myc/RasV12 , SV40 large T antigen/RasV12 , and BCL6/RasV12 gene combinations (Maeda et al., 2005a). ZBTB7A and BCL6 proteins are negative regulators of the p53 cell cycle control machinery (Haupt et al., 1997; Maeda et al., 2005b). The ZBTB7A protein binds to the promoter region of the p14ARF gene and represses its transcription (Maeda et al., 2005b). Recently, a role for fibronectin in triggering apoptozis (Ghosh et al., 2010a) and Mdm2-dependent p53 degradation (Ghosh et al., 2010b) has been shown. This interaction during the DNA damage response has been confirmed in a subsequent study (De Wever et al., 2011). Alternatively, binding of fibronectin to β1 integrin receptors in B-cell precursor leukemia cells also stimulates ZBTB7A expression (Astier et al., 2003). In this study, we investigated the transcriptional status of the p53-p14ARF-HDM2 pathway genes and their correlation with the plasma fibronectin levels in patients with CLL.
Materials and Methods
Peripheral blood samples of 40 patients (mean age 60±10.6) with CLL and 40 healthy volunteers without any disease were tested. The study was approved by the Istanbul University Faculty of Medicine Local Ethics Committee and all patients provided their informed consent. Characteristics of the patients are depicted in Table 1. The patients were evaluated according to the Rai and Binet systems which are the most frequently used clinical and prognostic classifications for staging the patients with early, intermediate, and advanced disease based on the involvement of the lymphatic system, anemia, and thrombocytopenia.
Lymphocytes were isolated using Ficoll Histopaque (Sigma-Aldrich). Following RNA isolation using the High Pure Total RNA Isolation Kit (Hoffmann La-Roche), cDNA was synthesized using the RevertAid™ First Strand cDNA Synthesis Kit (Fermantas). Gene expression analysis was performed both on the Lightcycler 1.2 instrument with G6PDH as the housekeeping gene (utilizing standard gene reference hydrolysis probes) and on the LightCycler 480 (using universal library probes [UPL] probes) (Hoffmann La-Roche). In the latter case the TATA-Box binding protein (TBP) gene was used as the reference gene. Primer and probe sequences are shown in Table 2. Expression levels were evaluated using the Basic Relative Quantification Software (Hoffmann La-Roche). Gene expression was analyzed using both systems. Individual expression values were calculated by dividing the expression level of the target gene by the value from the TBP gene. Plasma fibronectin levels were determined using the human fibronectin enzyme-linked immunosorbent assay kit (AssayPro).
ZBTB7A, Zinc Finger and BTB domain containing 7A; BCL6, B-cell lymphoma 6; ARF, Alternating Reading Frame; HDM2, Human homolog of murine Double Minute 2; UPL, universal library probes.
Exons 5–8 of the p53 gene were amplified by polymerase chain reaction and the fragments were analyzed for mutations by direct sequencing as previously reported (Buyru et al., 2003) with the exception that exon 5 was amplified in a single fragment. The sequences of the primers and the reaction conditions are available at request. The polymerase chain reaction (PCR) reaction contained 200 ng DNA, 0.2 mM dNTP, 15 pmol of each primer, and 1.5 U DreamTaq polymerase (Fermentas) in a total volume of 50 μL DreamTaq reaction buffer. The PCR products were subjected to direct sequencing (ABI) in both the forward and reverse directions.
Statistical comparison of gene expressions and fibronectin levels were performed with the Mann-Whitney U test, correlations between gene expressions were evaluated using Spearman correlation analysis and the associations between disease stage and gene expression by Kruskal-Wallis analysis.
Results
Expression of the p14ARF [median values for the patients and the controls 0.159 and 0.011 respectively, p<0.001, sample power%100 (alpha error level 5%, CI: 95%)], p53 [median values 2.713 and 0.155, p<0.001 sample power99.8% (alpha error level 5%, CI: 95%)] and HDM2 genes [median values 3.761 and 1.064, p<0.001 sample power 23.6% (alpha error level 5%, CI: 95%)] were significantly higher in the patients. Gene expression levels in the patients are shown in Figure 1.
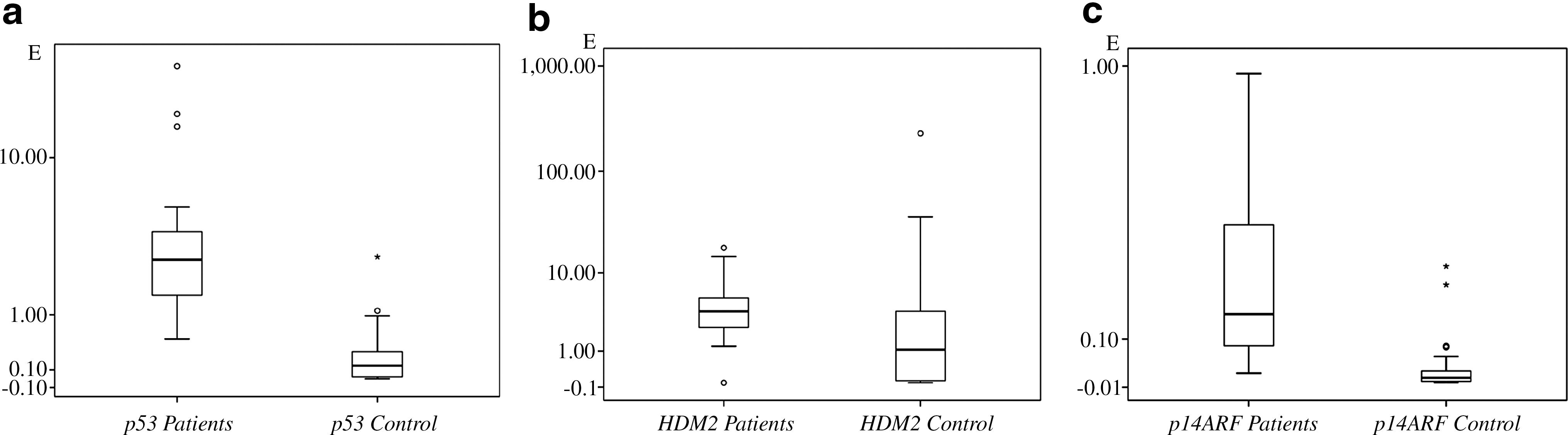
Expression levels of the
p14ARF expression was inversely correlated with p53 expression in the patients [p=0.013 sample power 99.8% (alpha error level 5%, CI: 95%)]. Expression of the p14ARF and HDM2 genes were closely correlated in the patients [p=0.018 sample power 100% (alpha error level 5%, CI: 95%)]. However, an inverse correlation between the same genes was noted in the controls (p=0.032). No association was observed between p53 gene expression and expression levels of the p14ARF and HDM2 genes [p=0.62, p=0.26 sample power 99.8% and 16.3% (alpha error level 5%, CI: 95%)] in the control group. We did not observe a significant difference between the patients and healthy controls in plasma fibronectin levels [p=0.2, sample power 54% (alpha error level 5%, CI: 95%)]. Plasma fibronectin levels were not associated with the expression of any gene both in the patients and the healthy controls. Plasma fibronectin levels in the groups are shown in Figure 2. We also did not observe an association between the disease stage and gene expression levels or plasma fibronectin levels. A notable decrease in p53 expression and an increase in HDM2 expression was observed in Stage IV patients which however, failed to reach statistical significance. p53 gene expression was inversely correlated with Binet staging (p=0.035) and no correlation was observed between Binet staging and p14ARF or HDM2 expression and plasma fibronectin levels. The expression levels are summarized in Table 3.
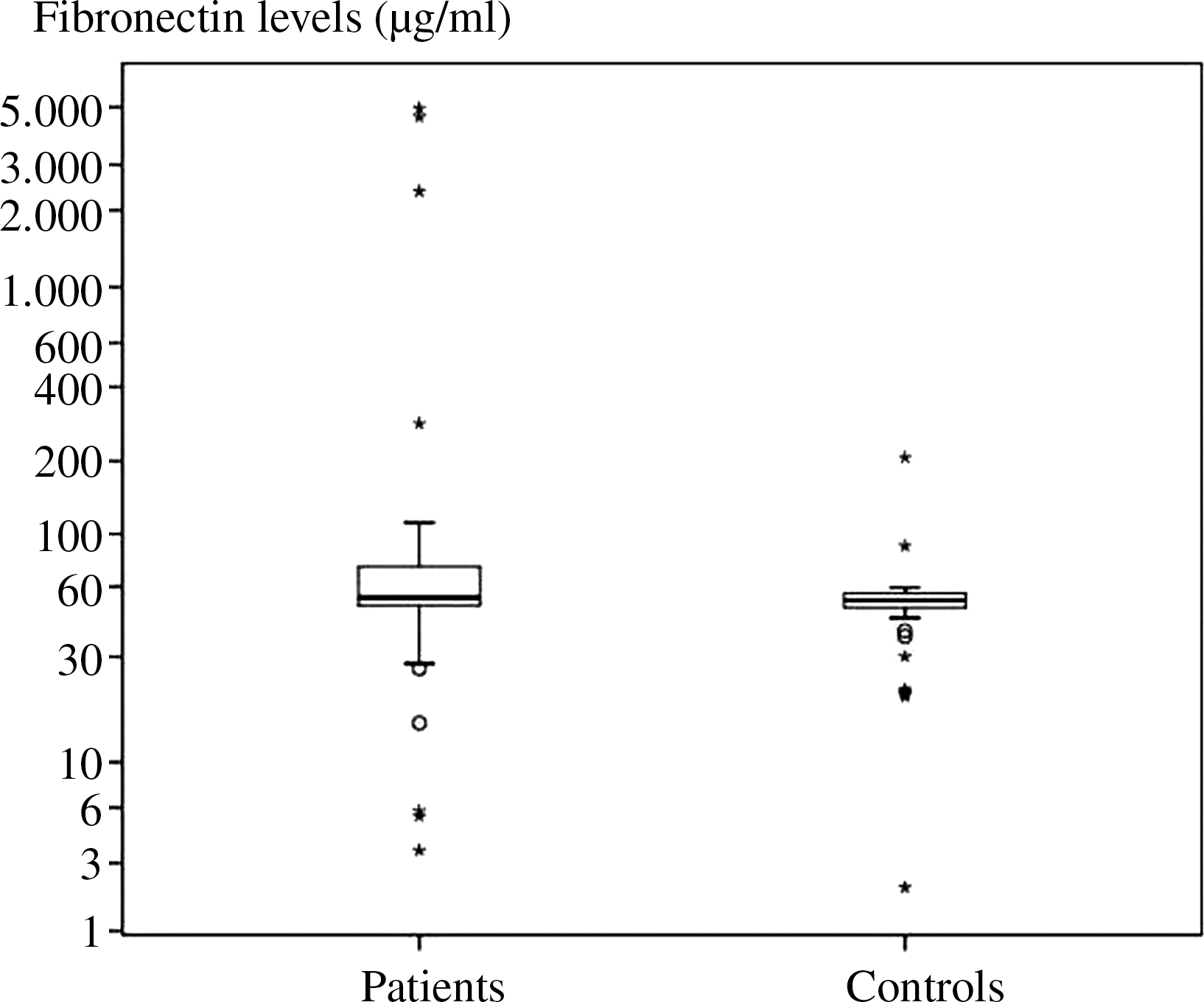
Plasma fibronectin levels in the groups. The asterisks indicate patients with values exceeding the 75% CI interval.
Expression of ZBTB7A and BCL6 genes were not detectable in the patients and the healthy controls. To further evaluate this finding we used a different platform and UPL probes to investigate expression of the ZBTB7A and BCL6 genes. However, we observed only expression of the reference gene but not the target gene using either the hybridization probes or the UPL system. Thus ZBTB7A and BCL6 gene expression was not observed in any sample.
Since the p53 gene is frequently mutated in cancers, the hot spot region of the p53 gene comprising exons 5–8 was analyzed by sequencing to investigate a possible role of p53 mutations. 85% of the mutations in the p53 gene occur in this region. In a single patient we observed a splice-site mutation (G→T) at nucleotide 13708 in the intron-exon boundary of exon 8 which resulted in an aberrant transcript with the deletion of a 24 bp region. Sequencing analysis also revealed a single nucleotide substitution at codon 213 in two patients which did not lead to an amino acid change. No further changes were observed in the remaining patients.
Discussion
CLL is a disease of the elderly and survival may reach 20 years after diagnosis. However, the duration of the disease is variable and outcomes differ (Moreno and Montserrat, 2008). There is a need for new genetic markers to help monitor the progression of the disease and to predict prognosis. The main factors affecting the p53 pathway are supression of p14ARF and overexpression of HDM2 (Efeyan and Serrano, 2007). Expression of the ZBTB7A and BCL6 genes has not been analyzed in CLL. The aim of this study was to evaluate the changes in the expression of these four genes in patients with CLL.
In our study, p53 gene expression was higher in patients. This is in accordance with a previous study in which p53 protein expression has been immunohistochemically shown in CLL (Haidar et al., 1997). We also noted a decrease in the p53 expression levels in patients with advanced disease which warrants further study.
The HDM2 protein is a ligase involved in p53 degradation (Haupt et al., 1997). HDM2 gene expression was higher in patients with leukemia than in the normal controls. Transcription of the HDM2 gene is regulated by p53 and high intercellular p53 protein levels stimulate HDM2 transcription (Yap et al., 1999), which is in line with our findings and with earlier reports (Watanabee et al., 1994; Ramos et al., 1995).
The p14ARF tumor suppressor protein binds to HDM2 and prevents degradation of p53. We observed higher p14ARF expression levels in the patients. High p14ARF expression has been reported as a consequence of the malfunction of common tumor suppressor pathways in aggressive lymphomas (Aguilera et al., 2002).
BCL6 gene expression was not observed in the patients and healthy controls. Our findings are consistent with the view that a feedback loop exists between p53 and BCL6 where the BCL6 protein can suppress p53 gene transcription (Margalit et al., 2006).
The p53 gene is the most frequently mutated gene in human cancer and overexpression of mutated p53 is observed in solid tumors. Expression of the mutant p53 protein has also been reported in CLL patients (Konikova and Kusenda, 2003). Further, possible mutations in the p53 gene that prevent the p53 protein from binding to the p53 response element in the BCL6 promoter might also result in lack of BCL6 expression (Faber and Chiles, 2006). Therefore, we also analyzed whether the p53 gene harbored any mutations in our patient cohort. A splice-site mutation was observed in a single patient indicating that p53 mutations do not play a causative role in CLL. This finding is in accordance with a recent report indicating that p53 mutations are not frequent in CLL (Zainuddin et al., 2011)
Considering the mutual interaction between HDM2, p53, and fibronectin plasma fibronectin levels were also analyzed in the patients. We did not detect differences between the plasma fibronectin levels of the patients and the control group, suggesting that plasma fibronectin levels are not associated with CLL development.
In our study, we could not detect ZBTB7A expression both in the CLL patients and in the healthy controls. To our knowledge, expression of these genes has not been analyzed in blood. There are only a few reports in solid tumors and cell lines. Since the ZBTB7A protein suppresses the p14ARF promoter, high levels of p14ARF expression are consistent with lack of ZBTB7A protein (Maeda et al., 2005a). In a recent study, miR-20a has been shown to inhibit translation of the ZBTB7A mRNA in MEF's (Poliseno et al., 2008). This interaction is considered to represent an alternative mechanism of tumor suppression. Whether this may explain the absence of ZBTB7A expression in our samples warrants further studies.
Conclusion
Our data indicate that aberrant expression of the p53 pathway genes p14ARF and HDM2 may be associated with the development of CLL. Further studies analyzing the interactions between these proteins in the leukemic cells are needed to elucidate the issue.
Footnotes
Acknowledgments
The present work was supported by the Research Fund of Istanbul University. Project No. T-985/06102006 and UDP-8444.
Disclosure Statement
No competing financial interests exist.