
Abstract
FY antigens are candidate minor histocompatibility antigens relevant to renal allograft rejection, but no data have been reported about their role in graft-versus-host disease (GVHD) incidence after human leukocyte antigen (HLA)-identical siblings hematopoietic stem cell transplantation (HSCT). The aim of this study was to examine the effect of donor/recipient disparity at FY antigens on the incidence of GVHD in Tunisian patients receiving an HLA-identical HSCT. This work enrolled 105 Tunisian pairs of recipients and their HLA-identical sibling donors of HSCs. FY genotyping was performed with the polymerase chain reaction–sequence-specific primer method and donor/recipient disparity for these antigens was analyzed at two levels: incompatibility and nonidentity. The case–control analyses showed no significant correlation between FY disparity and the incidence of either acute or chronic GVHD. Sample size calculation showed that 572 cases and 1716 controls would be necessary to be able to detect a significant association with 80% power and two-sided type I error level of 5% (α=0.05). The lack of association in the studied cohort may be explained by the low immunogenicity of FY antigens in HSCT context, compared with other antigens such as HA-1 and CD31.
Introduction
FY antigens belong to the Duffy blood group system that is of major interest in therapeutic medicine (Meny, 2010). This serologically determined system consists of two antithetic antigens, FY1 (Fya) and FY2 (Fyb), encoded by a single FY gene that maps to chromosome 1q22–q23 (Lautenberger et al., 2000). This gene contains four major allelic forms: two codominant alleles FY*1 and FY*2 coding, respectively, for the FY1 and FY2 antigens; FY*0 is the FY silent version and does not express FY antigens only at the erythroid level; and an FY*X allele producing a very weak FY2 antigen. The molecular bases of FY polymorphism have been well described (see, for example, Daniels, 2009). Interethnic analysis for FY alleles shows diversity when comparing Caucasian and sub-Saharan populations. In fact, it has been reported that FY null phenotype (lack of FY1 and FY2 antigens on RBCs) is very frequent in African people (∼90%) and this has been explained by certain epidemiologic factors such as the natural selection with malaria vivax, because Duffy molecule is the only RBCs receptor for Plasmodium vivax (Kwiatkowski, 2005; Daniels, 2009). In contrast, the FY-positive phenotype (expression of FY1 and/or FY2 antigens on RBCs) is common among Caucasians (Daniels, 2009). For Tunisians, FY allele frequencies were found (after having excluded FY*X, which had a frequency of 0.0174): FY*1=0.291 (expressed 0.260; silent 0.031); FY*2=0.709 (expressed 0.427; silent 0.282) (Sellami et al., 2008).
The Duffy molecule (gpD) is a transmembrane glycoprotein of 35–43 kDa expressed in two forms α and β of 338 and 336 amino acids, respectively. This molecule contains an NH2 extracellular domain containing 59 amino acids, which seems to be very important at the histocompatibility level (a candidate MiHAg), especially in transplantation contexts (Tournamille et al., 2003; Lerut et al., 2007). In fact, this peptide sequence contains the FY1/FY2, FY6, and a part of the FY3 antigen, which are linked to certain hemolytic diseases such as that affecting the newborn (HDN) (Tournamille et al., 2003; Cotorruelo et al., 2009). Among others, Duffy molecule is expressed on RBCs, cerebellar neurons, epithelial cells of nonerythroid organs, and endothelial cells lining postcapillary venules throughout the body, which represent the first site of contact with the donor alloreactive T cells after HSCT (Grimberg et al., 2007). Thus, we suspect that Duffy molecule polymorphism can affect the occurrence of the GVHD reaction after HLA-identical HSCT by acting as MiHAg. Therefore, we decided to examine association in a Tunisian cohort of patients receiving an HLA-identical sibling HSCT. For this purpose, we selected the FY*1/FY*2 (rs12075) polymorphism coding for FY1/FY2 antigens for molecular analysis.
Materials and Methods
Informed consent was obtained from all participants and/or their families to participate in this study according to a protocol approved by the ethical committee for scientific and medical research of the National Blood Transfusion Centre of Tunis (Tunisia).
Patient selection
The cohort contains 105 Tunisian pairs of patients and their HLA-identical siblings. The patients have undergone the HLA-matched HSCT in the National Bone Marrow Transplantation Centre of Tunis between January 2000 and March 2010. All patients received cyclosporine A and/or methotrexate for GVHD prophylaxis and had either grades 0–I/grades II–IV acute GVHD or chronic GVHD. For other clinical characteristics, see Figure 1.
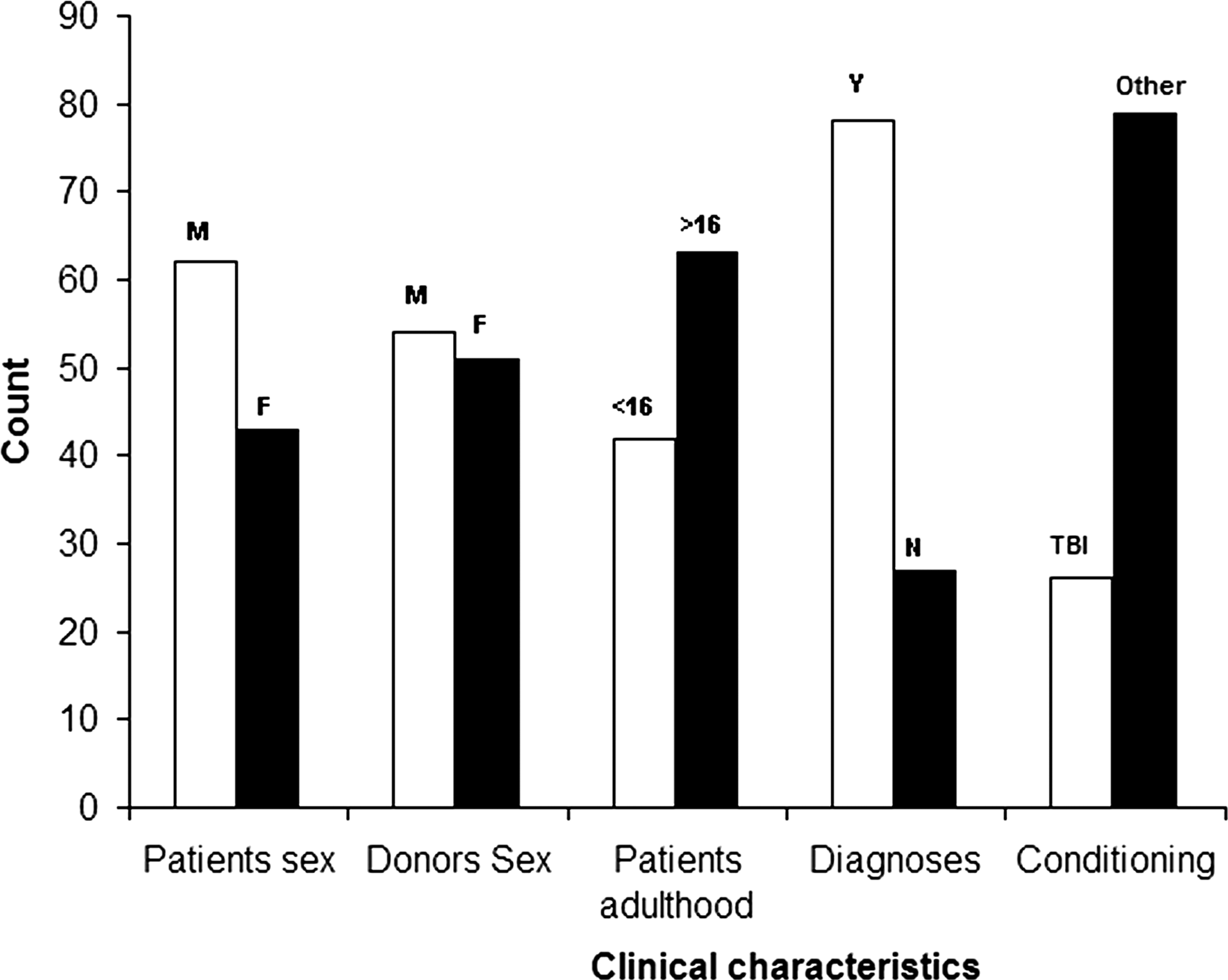
Clinical characteristics of the patient/donor pairs enrolled in the study (M, male; F, female; <or>16,<or>16 years; Y, malignant; N, nonmalignant; TBI, conditioning with TBI; Other, other conditioning regimens.
DNA samples
EDTA-treated blood samples were collected from all individuals before HSCT. DNA was purified from peripheral blood leukocytes using the DNA blood minikit (Qiagen) according to the manufacturer's instructions.
GVHD grading
The diagnosis and grading of acute and chronic GVHD were defined according to previously published standard clinical criteria (Przepiorka et al., 1995).
DNA amplification using polymerase chain reaction–sequence-specific primer
Molecular amplification was obtained using specific primers generating a 709-bp fragment for the FY alleles and a 434-bp fragment for the internal control, which is a sequence of the human growth hormone gene (HGH). The primers used in this work are those previously published (Mullighan et al., 1998). These primers were synthesized as oligonucleotides from PROLIGO Primers & Probes.
Reaction mixtures and amplification conditions
Amplification was carried out in a final volume of 10 μL containing 200 μM of each dNTP, 0.2 μM of the detection primers (detecting the specific allele), 0.1 μM of the control primers (detecting the HGH sequence), and 1.5 mM MgCl2 in 2 μL of 5X Green Buffer (Promega Corporation). Each polymerase chain reaction (PCR) was performed using 150 ng of genomic DNA and 0.4 units of Taq polymerase (Go Taq™ DNA Polymerase; Promega Corporation).
The amplification protocol was 94°C for 5 min followed by 30 cycles of amplification consisting of denaturation at 94°C for 30 s, annealing at 60°C for 1 min, and elongation at 72°C for 30 s (Gene Amp PCR System 9600; Perkin Elmer). The PCR products were visualized on agarose gels (1.5%) with UV illumination and photodocumented (UViTechTM).
Statistical analysis
Donor/recipient disparity at FY antigens is defined as follows: • Nonidentity: when the donor and the recipient are not carriers of the same FY genotype. • Incompatibility: when the donor is not compatible with the recipient for the FY alleles.
Chi-square and Fisher's exact tests were used to analyze the association between both FY nonidentity/incompatibility and either acute or chronic GVHD occurrence among the whole cohort. The cohort was subdivided into subgroups according to similarities of the peptide binding motif of their HLA class I; and the association between both FY nonidentity/incompatibility and either acute or chronic GVHD was tested with univariate analyses. Subgroups for which p-values were <0.2 were reexamined in multivariate logistic regression models including major risk factors such as adulthood and conditioning.Odds ratios (OR) and p-values for univariate and multivariate analyses were used to analyze the correlation between risk factors and the probability of both acute and chronic GVHD. These analyses, modeled according to the default method and Hosmer-Lemeshow goodness-of-fit, were performed with SPSS software (V 17.0). Positive correlation was considered only when p-value was <0.05 (type I error level or α).
A sample size calculation was done using the Power and Sample Size Program PS V3.0.43 (January 2009) (Dupont and Plummer, 1998) to determine the sample size needed to detect a significant association between the donor/recipient FY disparity and the GVHD incidence in the recipient of HSCs (i.e., to reject the null hypothesis H0). For this purpose, we made the following assumptions: two-sided type I error level (α) of 0.05, a type II error level (β) of 0.2 (power of 80%), and a control/case ratio of 30.
Results
The molecular genotyping approach showed the presence of all possible FY genotypes in the studied cohort. Clinically, the post-HSCT exams showed that all patients were evaluable for acute GVHD versus only 86 cases for the chronic form.
General case–control analysis
In this part of the study, we preformed a general association analysis using donor/recipient FY disparity and incidence of GVHD after HSCT. This analysis showed no significant correlation between either FY nonidentity or incompatibility and the incidence of acute and chronic GVHD among recipients of HSCs (Fig. 2). Thus, we decided to subdivide the cohort into subgroups according to similarities of the peptide binding motif of their HLA class I and reexamine these associations separately in each group.
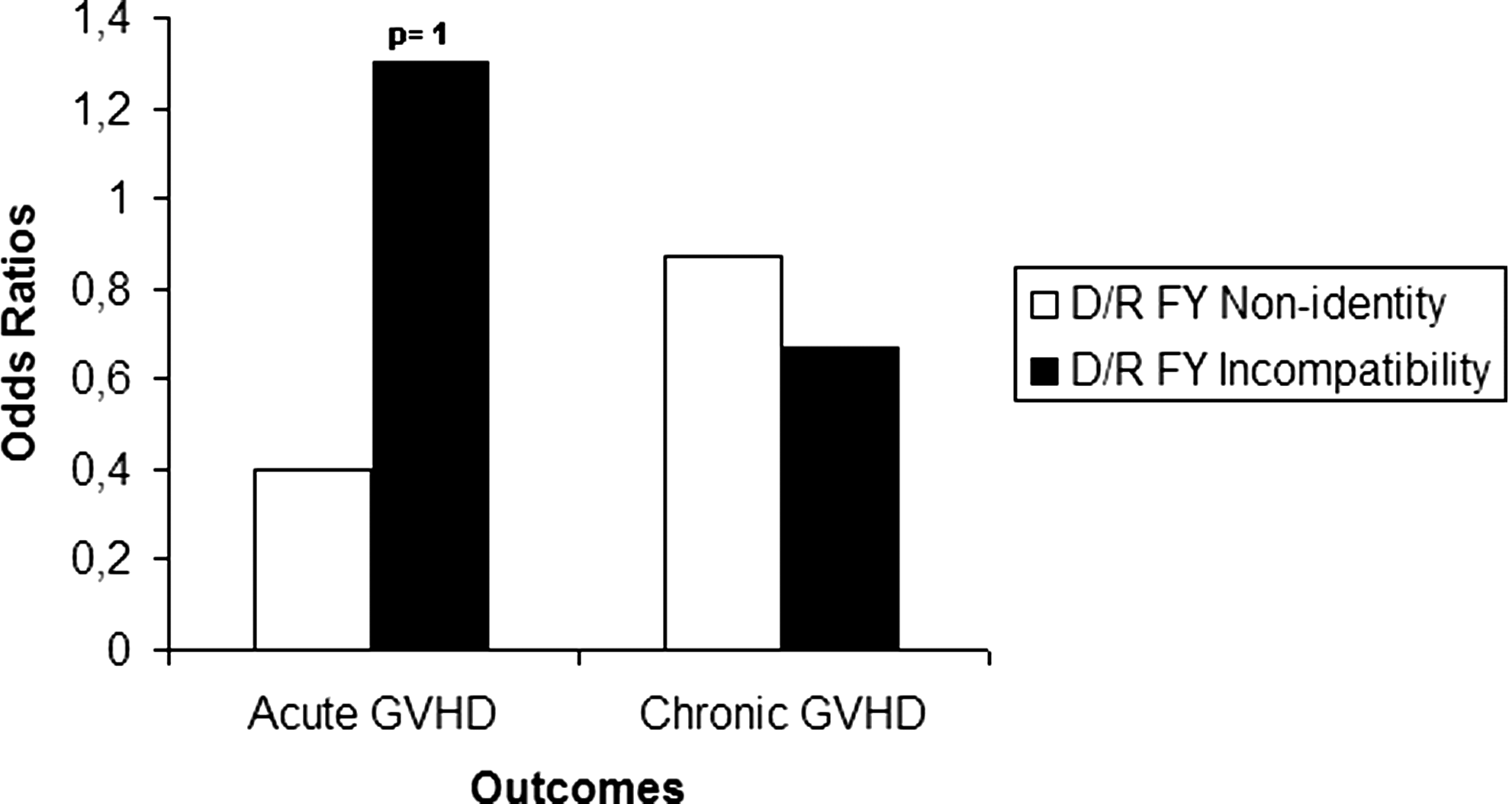
Analysis of association between both FY nonidentity and incompatibility and incidence of either acute or chronic GVHD among patients enrolled in this study. GVHD, graft-versus-host disease.
Subgroup case–control analyses
As there is a lack of a significant association in the general analysis, we suspected that subdividing the cohort into HLA class I subgroups may be a useful way to reinforce the statistical analysis. Subgroups are as follows: A2 (donor/recipient pairs carrier of the HLA-A*02 except A*0207, A*6802, and A*6901 alleles), other than A2 (pairs carrier of HLA-A alleles other than of the first group), B7-like (pairs carrier of HLA-B7, -B35, -B51, -B53, -B54, -B55, -B56, -B67, -B78 alleles), B44-like (pairs carrier of HLA-B37, -B41, -B44, -B45, -B47, -B49, -B50, -B60, -B61 alleles), and other than B7-like/B44-like (donor/recipient pairs carrier of HLA-B alleles other than of B7-like/B44-like groups). In this model, we did not find any significant association (Figs. 3 and 4). Consequently, we decided to reanalyze only groups in which we observed a p-value of <0.2 in a multivariate context including major risk factors such as adulthood and conditioning.
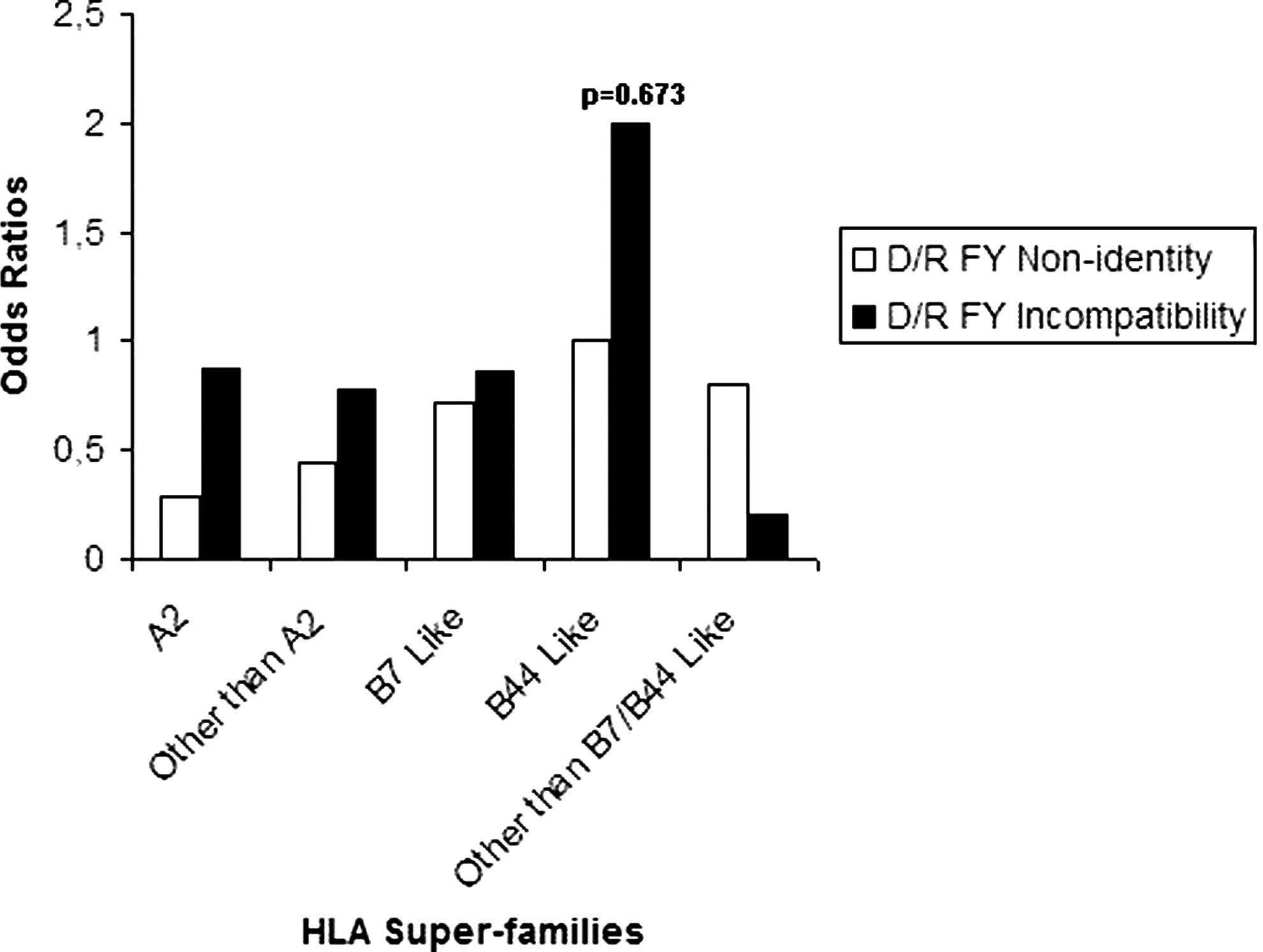
Univariate analysis of correlation between FY nonidentity and incompatibility and incidence of acute GVHD among each HLA class I subgroup. HLA, human leukocyte antigen.
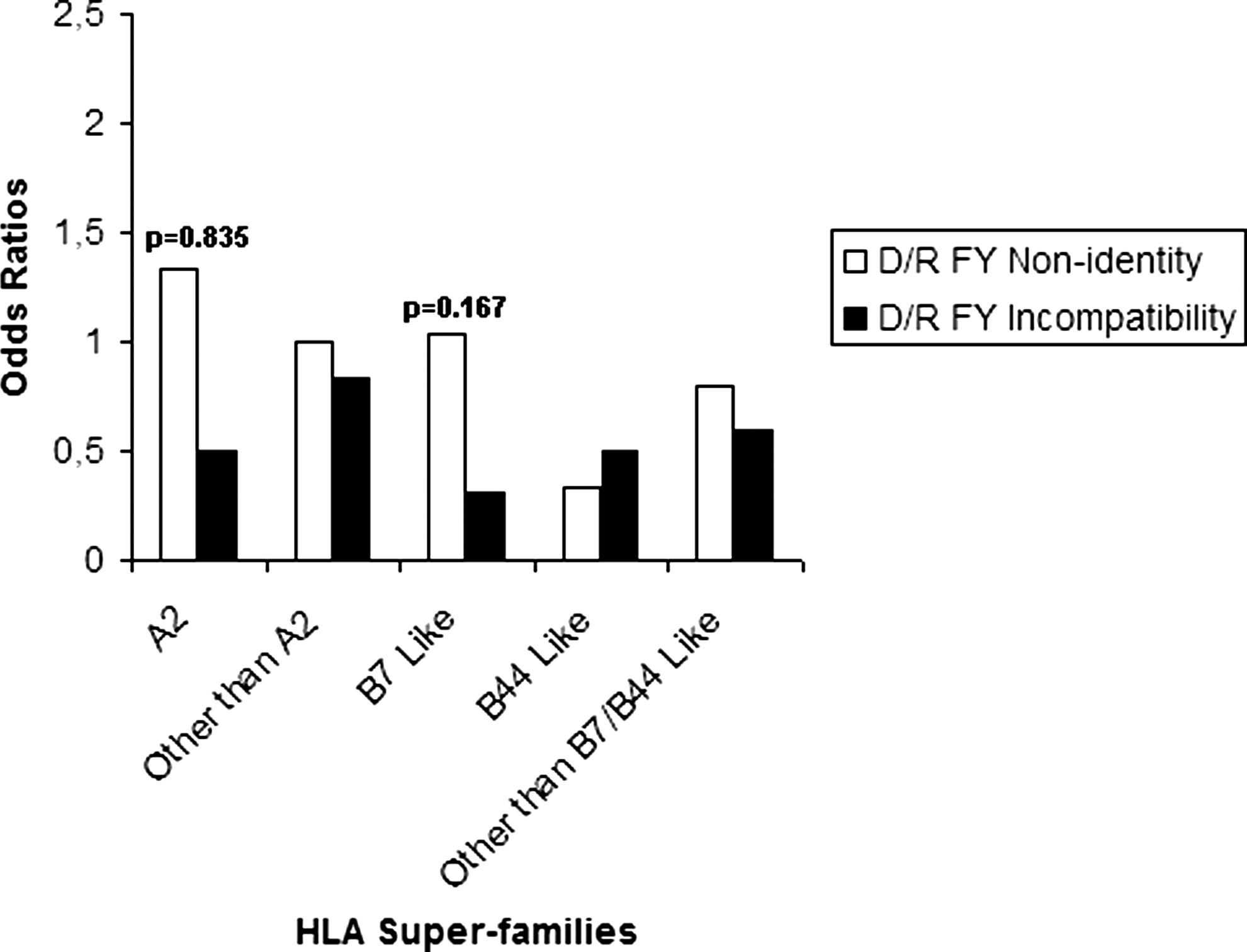
Univariate analysis of correlation between FY nonidentity and incompatibility and incidence of chronic GVHD among each HLA class I subgroup.
Multivariate analysis
This analysis considered only the relationship between FY1/FY2 identity and chronic GVHD incidence in the HLA-B7-like group (p=0.167, OR=1.031). In this model, we analyzed the combined effects of FY antigens identity, disease, donor/recipient sex mismatch, adulthood, and conditioning, which were described as potential risk factors (Fig. 5). As for the other analyses, interactions between these factors were not found to be significant in the GVHD paradigm.
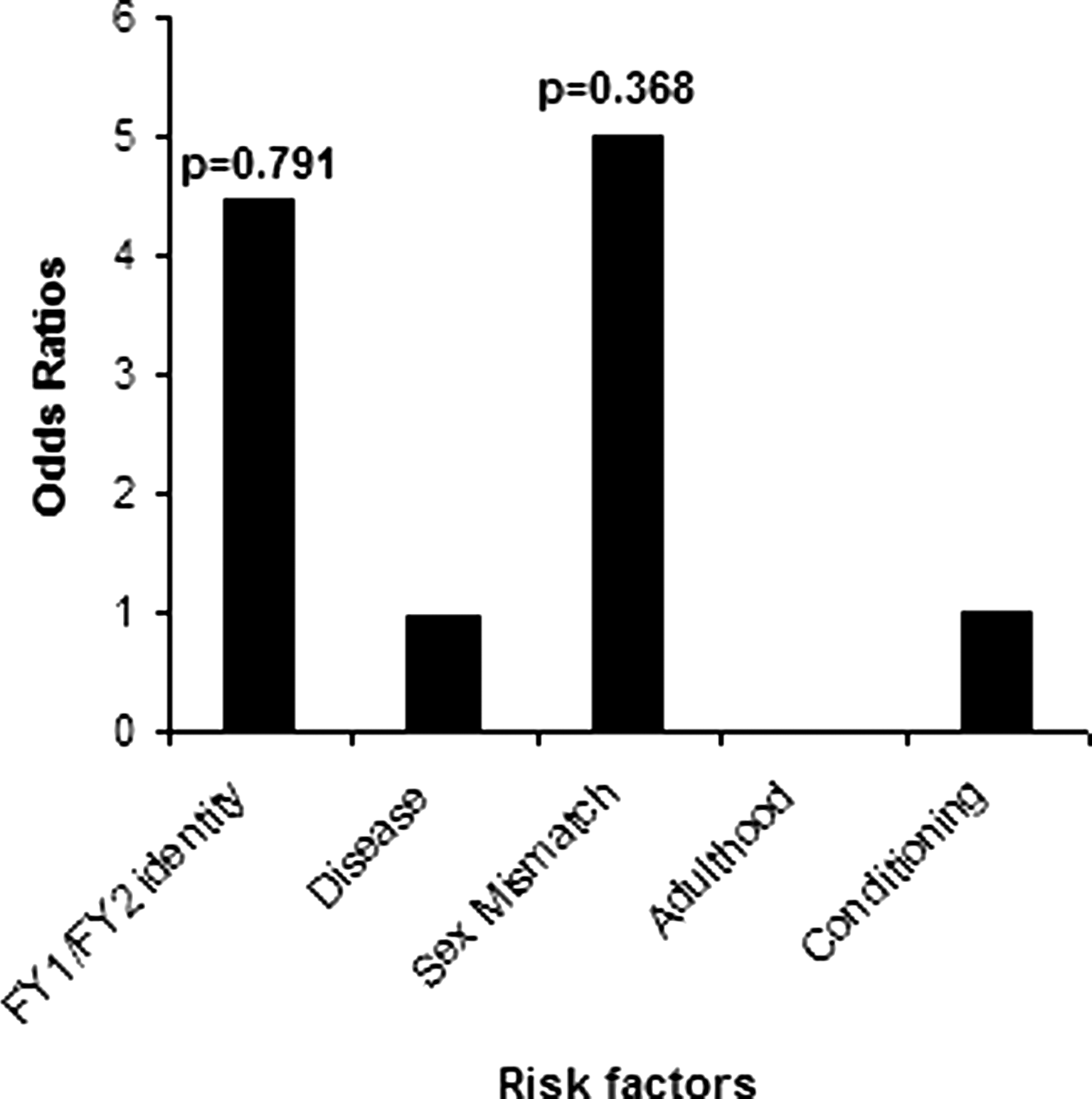
Multivariate logistic regression model of risk factors for chronic GVHD among HLA-B7-like positive patients.
Sample size calculation
We are planning a study of independent cases and controls with a control/case ratio of 30. Statistical data indicate that the probability of exposure among controls is 0.27 and the OR for disease in incompatible donor/recipient pairs relative to compatible donor/recipient pairs is 1.3 (incompatibility level) (Fig. 2). The sample size calculation found that a minimum of 572 cases and 1716 controls would be necessary to be able to detect a significant association with 80% power and two-sided type I error level of 5% (α=0.05).
Discussion
MiHAgs are allogeneic targets of T-cell–mediated GVHD effects following HLA-identical siblings HSCT (Sun et al., 2007; Ferrara et al., 2009; Paczesny et al., 2010). Several immunodominant MiHAgs have been identified, but defining a standard antigen panel remains a grand challenge for researchers. This difficulty is due to the high particularity of these antigens that are a subset of endogenously synthesized polymorphic products encoded by autosomal and sex-linked genes and classified as non-HLA antigens (Goulmy, 1996). Thus, performing GVHD association studies using ubiquitous antigens such as those of some blood group systems may be a practical way to define more immunodominants antigens. For these reasons, we decided to examine the effect of donor/recipient disparity in Duffy antigens (FY1/FY2) on the incidence of GVHD after HLA-identical siblings HSCT, because these antigens are candidate MiHAgs relevant to renal allograft rejection (Lerut et al., 2007).
The general case–control analysis showed no significant correlation between donor/recipient disparity at FY antigens and the incidence of either acute or chronic GVHD among recipients of HSCs. Thus, we decided to subdivide the whole cohort into subgroups according to similarities of the peptide binding motif of HLA class I to reinforce the statistical analysis but we did not find any significant results. The lack of association between FY disparity and GVHD incidence was also observed when we performed a multivariate analysis for the HLA-B7-like group that showed a p-value of <0.2. In this model, we included more risk factors such as donor/recipient sex mismatch and conditioning with TBI, but it appeared that the combined effects of these factors have no influence on the pathogenesis of GVHD. Thus, it appears from these analyses that donor/recipient disparity at FY antigens is not critical in the GVHD paradigm in the studied cohort. This finding may be explained by several lines of reasoning. First, we believe that the absence of relevant correlation is due to the low immunogenicity of the FY antigens compared with other antigens such as HA-1 and CD31. It has been reported that there is a difference at the immunodominance levels between all described MiHAgs, because each one is a complex between an HLA ligand and an immunogen peptide and it is the combinatorial association that is recognized by the alloreactive T cells (Simpson et al., 2002). Further, there are no data suggesting the involvement of FY antigens in T-cell–mediated immune responses such as those in GVHD reaction. In fact, immunogenicity of FY antigens has been usually described as potential cause of developing allo-antibodies, especially anti-FY1, and, therefore, humoral immunity (Meny, 2010). The contribution of humoral immunity in the GVHD paradigm is poorly defined, although that Miklos et al. reported a correlation between antibody responses to H-Y MiHAg and chronic GVHD (2005). Thus, examining the anti-FY level in sera of recipients who were mismatched with their donors of HSCs at Duffy antigens may be useful.
Likewise, the use of a CSA/MTX-based prophylaxis protocol by patients enrolled in this study may explain the lack of a significant association. This prophylaxis is used to prevent GVHD incidence after HSCT, and subsequently, the real number of patients who developed this outcome remains relatively ambiguous. Thus, we believe that the real effect of FY disparity cannot be evaluated in subjects who receive this protocol. We need to investigate this effect in recipients who had not received a GVHD prophylaxis. This procedure may be feasible only in murine models, because the medical and ethical duties require the physician to provide a prophylaxis protocol to the patient.
To the best of our knowledge, this is the first work examining the effect of donor/recipient FY disparity on GVHD incidence after HLA-identical siblings HSCT. The study revealed a lack of significant correlation, but further experiments are needed to confirm this finding. Further, we believe that it could be very important to investigate the effect of FY disparity on other endpoints such as leukemia relapse, disease-free survival, and overall survival.
Footnotes
Acknowledgments
The authors thank the patients and their donors who participated in this study. The authors are also very grateful to the research and medical staff of the “National Blood Transfusion Center and National Bone Marrow Transplantation Centre of Tunis” for their valuable technical assistance. This work was supported by the Tunisian Ministry for Higher Education and Scientific Research (UR06SP04) and the Tunisian Ministry of Public Health.
Disclosure Statement
No competing financial interests exist.