
Abstract
Oxidative damage is thought to play a pivotal role in the pathogenesis of nonalcoholic fatty liver disease (NAFLD). Glutathione-S-transferases (GSTs) are involved in cell protection against oxidative stress. We examined whether GSTM1, GSTT1, and GSTP1 polymorphisms are associated with NAFLD in a sample of the Iranian population. The current case–control study included 83 patients with NAFLD and 93 healthy subjects. The GSTM1 and GSTT1 polymorphisms were analyzed by multiplex polymerase chain reaction (PCR). The GSTP1 polymorphism was detected by tetra amplification refractory mutation system-PCR assay. The GSTM1-null genotype was significantly associated with the development of NAFLD (odds ratios [OR]=2.171, 95% confidence intervals [CI]=1.188–3.970, p=0.015). The GSTP1 Val allele was shown to be a risk factor for NAFLD (OR=1.739, 95% CI=1.089–2.777, p=0.024). The GSTT1 polymorphism was not significantly different between control and patient groups (p=0.221). This study showed that GSTM1 and GSTP1, but not GSTT1, genetic polymorphisms are associated with NAFLD in a sample of the Iranian population, and may be used to determine the risk of development of NAFLD.
Introduction
There is a growing line of evidence that polymorphisms in genes encoding enzymes responsible for xenobiotic metabolism are among the key players in determining inter-individual susceptibility to liver-related diseases (Burim et al., 2004; Ghobadloo et al., 2004; Palli et al., 2005; Abd El-Moneim et al., 2008; Kandemir et al., 2008; Kiran et al., 2008; Kordi-Tamandani et al., 2011). Glutathione-S-transferases (GSTs) are a family of enzymes broadly expressed in mammalian tissues, and have been demonstrated to be critical in the conjugation reaction in phase II xenobiotics metabolism and catalyze reactions between glutathione and a variety of potentially toxic and carcinogenic electrophilic compounds (To-Figueras et al., 1997; Singh et al., 2008; Mo et al., 2009). Based on their primary structure, the distinct cytosolic isoenzymes have been assorted to separate classes that are coded into at least eight diverse loci: μ (GSTM), θ (GSTT) π (GSTP), σ (GSTS), α (GSTA), κ (GSTK), o (GSTO), and τ (GSTZ) (To-Figueras et al., 1997; Mo et al., 2009). GSTT1, GSTM1, and GSTP1 are mapped on chromosomes 1p13.3, 22q11.2, and 11q13, respectively, and have gained the most attention (Mo et al., 2009). Homozygous deletion of GSTM1 and GSTT1 genes is the result of homologous recombination of a number of repeats spanning the left and right of these genes. These deletions, referred to as GSTM1 null and GSTT1 null, result in the lack of enzyme activity. The GSTP1 polymorphism is the result of a base substitution (A to G), which leads to an amino acid replacement (codon 105, isoleucine to valine) in the GSTP1 binding site and modifies enzyme catalytic activity (Hur et al., 2005).
A limited number of studies have evaluated the association of GST genotype profile with liver-related diseases. Several have reported a positive association between the GSTM1 and GSTT1 null genotypes and GSTP1-Val (105) polymorphisms and increased risk of alcoholic pancreatitis, alcoholic cirrhosis, hepatocellular carcinoma, hepatis B virus (HBV) infection progression, NAFLD, and cryptogenic cirrhosis development (Frenzer et al., 2002; Burim et al., 2004; Ghobadloo et al., 2004; Mohammadzadeh Ghobadloo et al., 2006; Abd El-Moneim et al., 2008; Hori et al., 2009; Khan et al., 2009; Maruyama et al., 2010). The current study was carried out to determine the impact of GSTM1, GSTT1, and GSTP1 genetic polymorphisms on the susceptibility of NAFLD.
Materials and Methods
Eighty-three patients with NAFLD and 93 healthy individuals were recruited for this study. The local Ethics Committee of Zahedan University of Medical Sciences approved this study, and informed consent was obtained from all participants. The study design and the enrolment procedure have been previously described in detail (Hashemi et al., 2011). Genomic DNA were extracted from peripheral blood as previously described (Hashemi et al., 2010a).
Tetra primer amplification refractory mutation system–polymerase chain reaction (T-ARMS-PCR) is a simple and rapid method for detection of single-nucleotide polymorphism (Ye et al., 2001; Hashemi et al., 2010b, 2011). We designed T-ARMS-PCR for detecting the GSTP1 exon 5, 105 Ile/Val (rs1695) polymorphisms. We used two external primers (Forward outer: 5′ CAGGTGTCAGGTGAGCTCTGAGCACC 3′, Reverse outer: 5′ ATAAGGGTGCAGGTTGTGTCTTGTCCCA 3′), and the two internal primers were (Forward inner (A allele or Ile allele): 5′ CGTGGAGGACCTCCGCTGCAAATCCA 3′, Reverse inner (G allele or Val allele): 5′-GCTCACATAGTTGGTGTAGATGAGGGATAC-3′). Product sizes were 233 bp for A allele and 290 bp for G allele, whereas the product size of the two outer primers was 467 bp, as schematically shown in Figure 1.
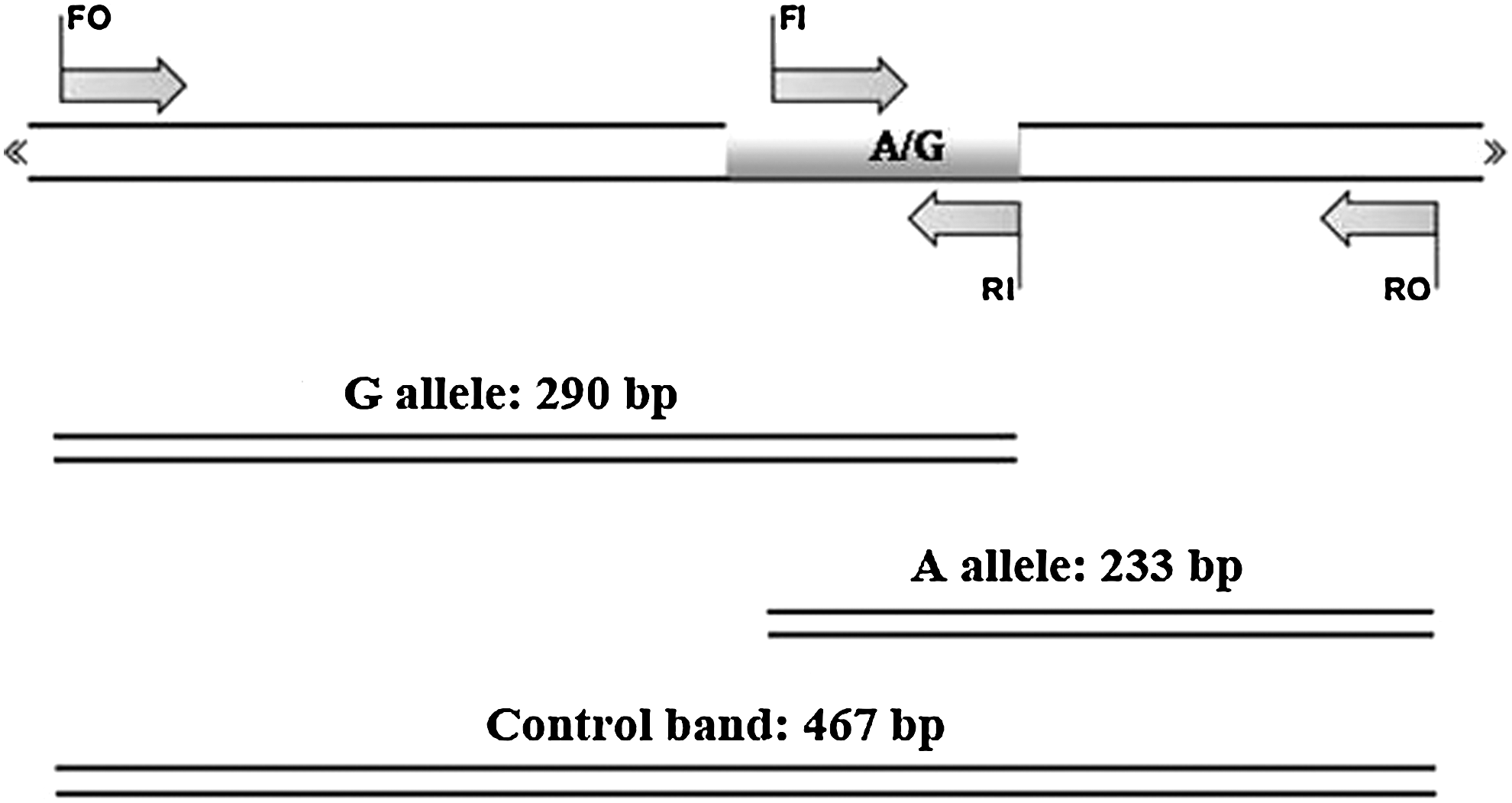
Schematic illustration of tetra amplification refractory mutation system–polymerase chain reaction (PCR) assay for detection of glutathione-S-transferase P1 (GSTP1) Ile105Val (rs1695 A/G). Two forward and two reverse specific primers are used to produce three potential products. Product sizes were 233 bp for A allele, 290 bp for G allele, and 467 bp for two outer primers (control band).
PCR was performed by using commercially available PCR premix (AccuPower PCR PreMix; BIONEER, Daejeon, Korea) according to the manufacturer's instructions. Briefly, 1 μL template DNA (∼100 ng/μL), 1 μL of each primer (10 pmol/μL), and 15 μL DNase-free water were added to AccuPower PCR PreMix. Amplification was performed with an initial denaturation step at 95°C for 5 min, followed by 25 cycles at 95°C for 30 s, 65°C for 20 s, and 72°C for 15 s with a final extension at 72°C for 10 min. The PCR products were separated by electrophoresis in 2% agarose gels, and visualized by ethidium bromide staining. The T-ARMS-PCR methods were effectively applied to the genotyping polymorphism of GSTP1 105 Ile/Val (rs1695). The genotypes determined by this method (Fig. 2) were concordant with those determined by sequencing (Supplementary Fig. S1; Supplementary Data are available online at
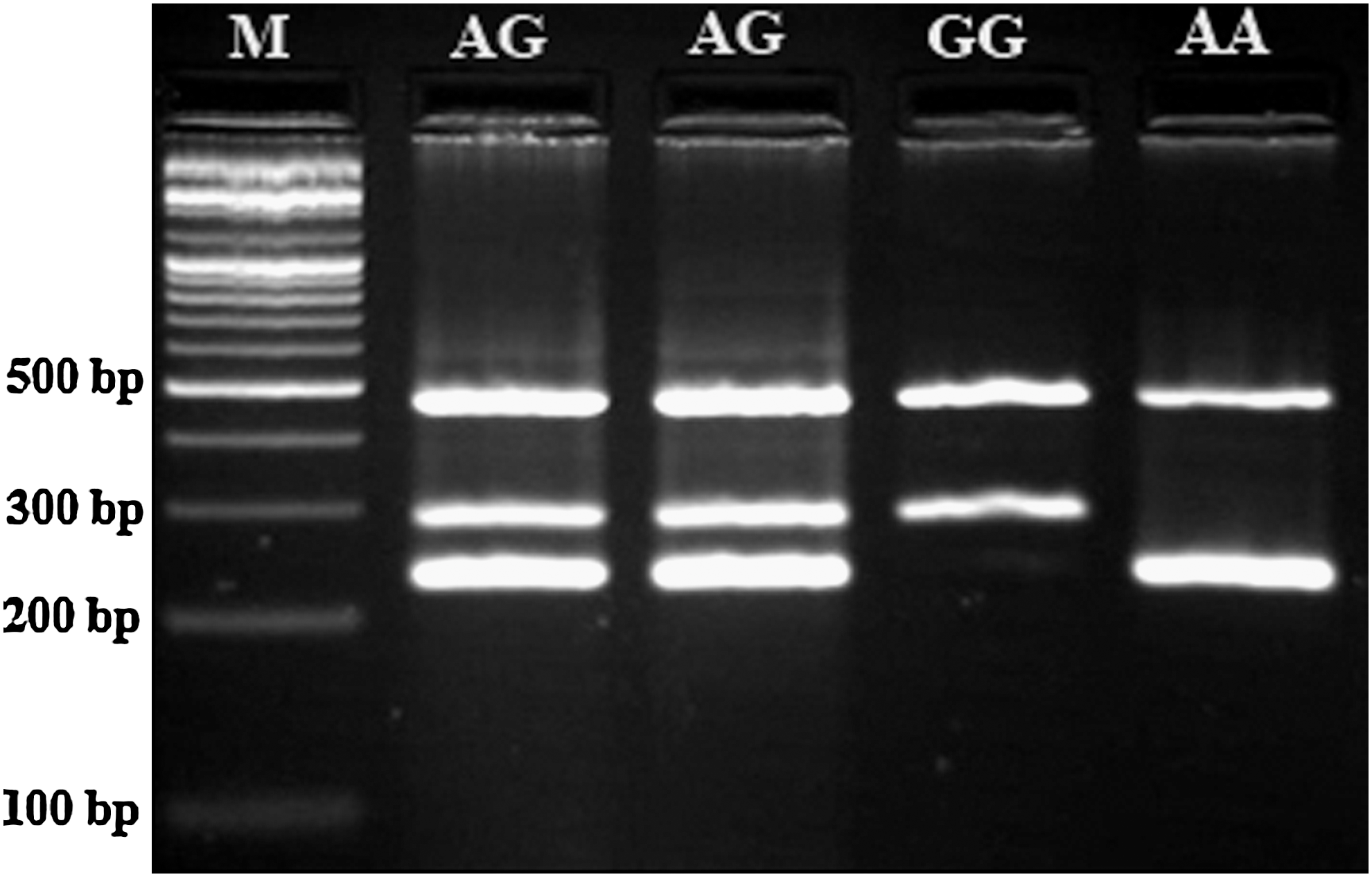
Photograph of the PCR products of the (GSTP1) Ile105Val (rs1695 A/G) gene polymorphism. M, DNA marker.
Multiplex PCR was used to simultaneously detect the GSTT1 and GSTM1 genes polymorphism in the presence of the internal control. Primers for the GSTM1 polymorphism were as follows: forward 5′ GCTGCCCTACTTGATTGATG 3′ and reverse 5′ CCCCAAATCCAAACTCTGTC 3′. Primers for GSTT1 were as follows: forward: 5′ TTCTGCTTTATGGTGGGGTC 3′ and reverse 5′ GTGATGTTCCCTGTTTTCCT 3′. Toll-like receptor 2 (TLR2) primers were as follows: forward 5′ GATGCATTTGTTTCTTACAGTGAGCG 3′ and reverse 5′ TCTCATCAAAAAGACGGAAATGGG 3′, and were used as an internal control.
PCR was performed as described above with amplification by denaturation at 95°C (5 min), followed by 25 cycles of 95°C for 30 s, 60°C for 25 s, and 72°C for 30 s, with a final extension step of 72°C for 10 min. The PCR products were electrophoresed in 2% agarose gels, and visualized by ethidium bromide staining (Fig. 3).
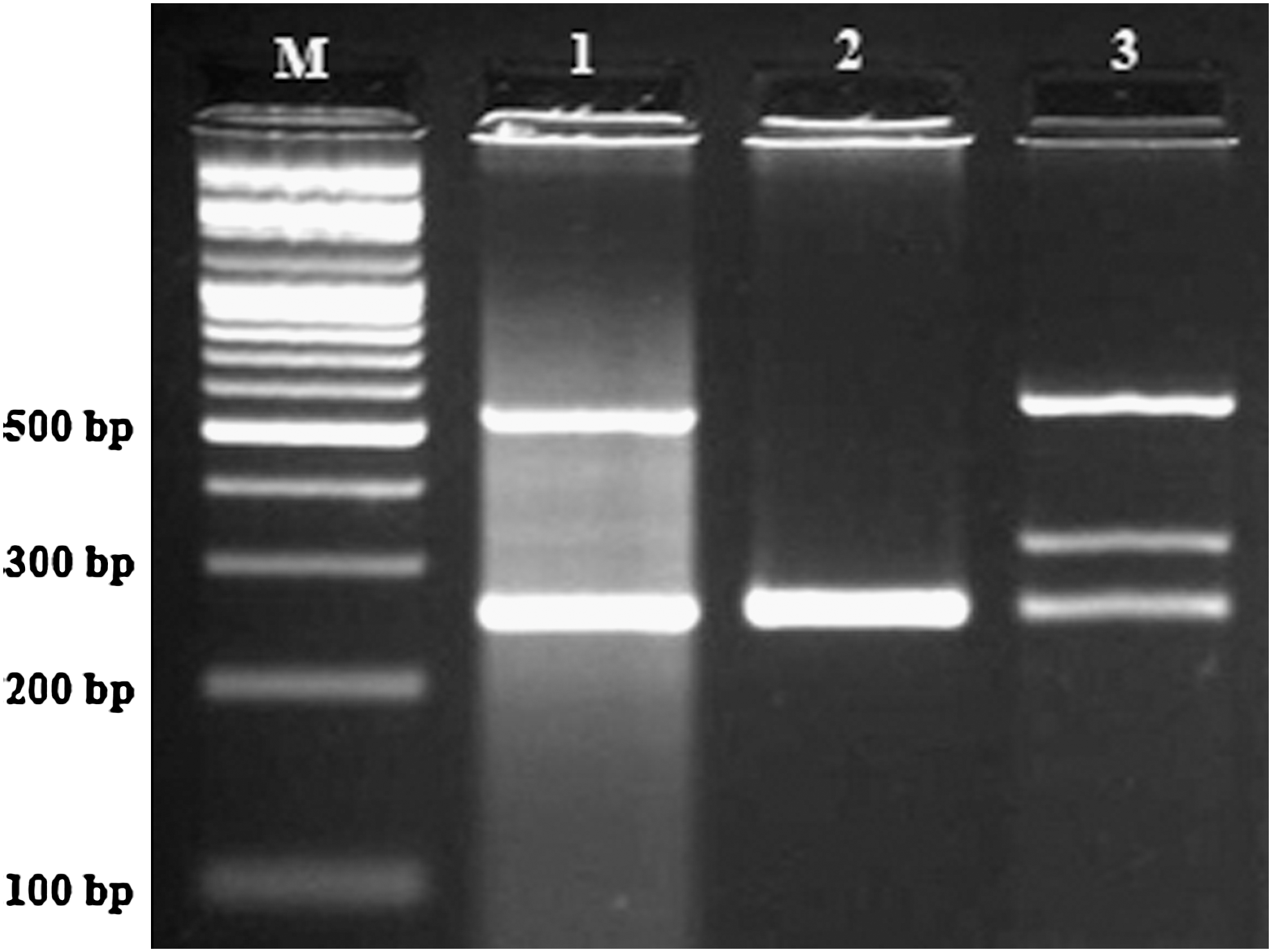
Representative multiplex PCR products resolved by agarose gel electrophoresis to detect the presence or absence of the GSTT1 and GSTM1 genes simultaneously in the same tube in the presence of an internal control. GSTT1; 542 bp, GSTM1; 325 bp, Toll-like receptor 2; 259 bp (control). M, DNA marker. Lane 1: GSTT1 present GSTM1 null genotype; lane 2: both GSTT1 and GSTM1 present null genotype; lane 3: both GSTT1 and GSTM1 present genotype.
DNA from samples positive for GSTM1 and GSTT1 genotypes yielded bands of 325 and 542 bp, respectively, whereas the internal positive control (TLR2) PCR product corresponded to 259 bp. Absence of 325 and 542 bp bands indicated homozygous null genotypes of GSTM1 and GSTT1, respectively. To ensure genotyping quality, we re-genotyped all polymorphisms in random samples and found no genotyping mistake.
The association among the GSTM1, GSTT1, and GSTP1 genotypes and the risk of NAFLD was estimated by using odds ratios (OR) with 95% confidence intervals (CI). All computational analyses were executed by using statistical software package SPSS 18.
Results
Eighty-three NAFLD (50 men and 33 women; age 40.45±12.12) and 93 normal subjects (42 men and 51 women; age 42.33±16.25) were in the current study. Age and sex were not significantly different between the groups (p>0.05). GSTM1, GSTT1, and GSTP1 genotype distribution were compared between NAFLD and healthy individuals in Table 1. The frequency of the GSTM1 null genotype was significantly higher in NAFLD (57.8%) than in the control group (38.7%) and is a risk factor for susceptibility to NAFLD (OR=2.171, 95% CI=1.188–3.970, p=0.015). No significant differences were detected between the NAFLD group (2.4%) and the control group (0%) for the GSTT1 null mutation, but a significant difference was observed between controls and patients with regard to the GSTP1 105Ile/Val polymorphism (p=0.012). We found that the Val allele is a risk factor for susceptibility to NAFLD (OR=1.739, 95% CI=1.089–2.777, and p=0.024). Based on our findings, sample power was calculated for GSTT1, GSTM1, and GSTP1 by comparison of each genotype with the sum of other related genotypes at each polymorphic region by using STATA 10 and are shown in Table 1.
NAFLD, nonalcoholic fatty liver disease; GST, glutathione-S-transferase; OR, odds ratios; CI, confidence intervals.
In addition, the combination of GSTM1, GSTT1, and GSTP1 genotypes was analyzed (Table 2). A synergistic gene–gene interaction was found in the GSTM1 Null/GSTP1 Ile/Val or Val/Val genotypes (OR=5.21, 95% CI=2.14–12.69, p<0.001).
Discussion
In the current study, we examined the association between NAFLD and polymorphisms of GSTM1, GSTT1, and GSTP1 in a sample of the Iranian population. We found no significant difference between NAFLD and healthy individuals with regard to the GSTT1 null genotype. However, the GSTM1 null genotype was significantly more frequent in NAFLD than control subjects, and the GSTP1 Ile 105 allele was shown to be a risk factor for susceptibility to NAFLD in this sample of the Iranian population. This corresponds with the findings of Hori et al. (2009), who also described increased frequency of the GSTM1 null genotype in patients with NAFLD compared with healthy individuals. However, they did not observe a statistically significant difference in GSTP1 105 Ile/Val and GSTT1 null polymorphisms (Hori et al., 2009). Recently, it has been reported that there was no association between promoter methylation and expression of GSTT1 and GSTP1 genes and risk of NAFLD (Kordi-Tamandani et al., 2011).
Given the functional significance of GSTs in cellular protection from environmental and oxidative stress, genetic variations among individuals that alter enzyme activity are one of the main factors leading to failure in the critical functions of the liver. The oxidative metabolism of xenobiotics is carried out largely by cytochrome P450 and GST enzymes, in which the latter is present in the cytosol, mitochondria, and microsomal membrane of almost all tissues throughout the body (Wang et al., 2000; Hardwick et al., 2010). GSTs play an important role in the detoxification of exogenous as well as endogenous compounds such as toxic products of lipid oxidation (Laborde, 2010). The liver plays a vital role in regulating fatty-acid and triglyceride metabolism by synthesizing, storing, releasing, and oxidizing free fatty acids (FFA). Disruption of triacylglycerol release, synthesis, or oxidation pathways could contribute to its accumulation in the liver (Utzschneider and Kahn, 2006). Nonalcoholic fatty NAFLD is the most common reason for abnormal liver function, and may occur in 10%–30% of the population (Hori et al., 2009). Complications associated with NAFLD include hepatocyte accumulation of triglycerides, chronic oxidative stress levels, insulin resistance (IR), inflammation, and fibrosis (Hardwick et al., 2010). NAFLD can progress from simple steatosis to nonalcoholic steatohepatitis, hepatocyte necrosis, fibrosis, and cirrhosis of the liver (Song et al., 2005). This progression is thought to be the consequence of two physiologic incidents: IR and oxidative stress (Charlton, 2004). In IR states, the amount of FFA accessible for oxidation surpasses the mitochondrial capacity, which results in increased lipid peroxidation. Excess acetyl CoA enters the citric acid cycle, thereby augmenting NADH (nicotinamide dinucleotide phosphate) production and delivery of electrons to the respiratory chain. This process could lead to mitochondrial failure leading to increased production of reactive oxygen species (ROS) (Schreuder et al., 2008; Hori et al., 2009). Elevated ROS results in hepatocellular injury by several mechanisms, including inactivation of membrane sodium channels, oxidative protein modifications, induction of lipid peroxidation, and cytokine production (Eaton and Bammler, 1999; Petlevski et al., 2003).
Lipid peroxides formed due to oxidative stress serve as endogenous substrates for GSTs. The GSTs are phase II metabolic enzymes and play a critical role in the defense against oxidative stress products and a variety of electrophilic compounds. In the human liver, hepatocytes contain high levels of GSTM and GSTT, whereas GSTP is predominantly expressed in the bile ducts (Eaton and Bammler, 1999; Lakehal et al., 1999).
The GSTM1 and GSTT1 null genotypes are associated with deficiencies in GSTM1 and GSTT1 enzyme activity (Seidegard et al., 1988; Bolt and Thier, 2006), and the GSTP1 Ile105Val polymorphism is associated with altered catalytic function (Watson et al., 1998; Cote et al., 2009). Individuals with GSTM1 and GSTT1 null genotypes or GSTP1 105Val allele would be expected to have decreased GST detoxification and, thus, potential increases in the levels of toxic metabolites.
In conclusion, our findings suggest that a deletion of the GSTM1 gene and functional polymorphism le105Val of the GSTP1 gene may contribute to vulnerability to NAFLD in a sample of Iranian subjects. One limitation of this study is the low sample size. Further research in larger samples is needed to confirm these findings.
Footnotes
Acknowledgments
This work was supported by a research grant from Zahedan University of Medical Sciences. The authors thank all subjects who willingly participated in the study. S.G. was supported by a Parker B. Francis Fellowship in respiratory disease.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.