
Abstract
Keratins and keratin-associated proteins (KAPs) are large heterogeneous groups of proteins that constitute about 90% of the wool fiber. The genes encoding the high glycine-tyrosine (HGT) KAPs are the first sub-group of KAP genes expressed in the wool follicle and just after expression of the keratin genes. Little is known about variation in these genes, which led us to investigate two HGT-KAP genes, KRTAP7-1 and KRTAP8-1. Polymerase chain reaction–single-stranded conformational polymorphism analysis was used to investigate these genes in 250 Romney-cross sheep. For KRTAP7-1, two unique banding patterns were detected for amplicons that spanned the entire coding region. Sequencing confirmed the presence of two sequences with only one nucleotide difference (c.173G/A) putatively resulting in p.Ser58Asn. One was identical to the published ovine KRTAP7-1 sequence. For KRTAP8-1, five unique banding patterns were detected in an amplicon that spanned the entire coding region. Sequencing revealed five different DNA sequences, all of which were highly homologous to the previously reported ovine KRTAP8-1 sequence. Among these five sequences, four single-nucleotide substitutions were identified and three of them were located in the coding region. One of these was nonsynonymous and would putatively result in p.Tyr34Asn. The variation detected in KRTAP7-1 and KRTAP8-1 may influence their expression or protein structure.
Introduction
The HGT-KAPs are predominantly present in the orthocortex and are thought to have some associations with the crimp of wool fiber (Powell and Rogers, 1990). In the wool follicle, the expression of the HGT-KAPs follows shortly after expression of the keratins, but before the expression of the HS and UHS KAPs (Rogers, 2006). The content of HGT-KAPs in hair/wool fiber varies considerably within and between species, ranging from <3% in human hair and Lincoln sheep wool, through to 4%–12% in Merino wool, 18% in mouse hair, and 30%–40% in echidna quill (Gillespie, 1990). The HGT-KAP content appears to be influenced by dietary, physiological, and genetic factors (Gillespie, 1990).
In sheep, there are three known HGT-KAPs families, KAP6, KAP7, and KAP8. KAP6 appears to be a multi-gene family (Fratini et al., 1993; Gong et al., 2011), but only one member has been identified for each of the KAP7 and KAP8 families and these have been named KAP7-1 and KAP8-1, respectively (Kuczek and Rogers, 1987). While allelic variation in the KAP6 genes has been reported recently (Gong et al., 2011), variation in KAP7-1 was previously described using Southern hybridization–restriction fragment length polymorphism (RFLP), and no sequence information was given (McLaren et al., 1997), and variation in KAP8-1 was only described in the promoter region (Wood et al., 1992) and has not been investigated in the coding region.
Given that the KAP7-1 and KAP8-1 genes are clustered with the KAP6 genes on a chromosome region that appears to be associated with variation in wool fiber diameter in a Merino half-sib family (Parsons et al., 1994), it appears to be worthwhile investigating genetic variation in the ovine KAP7-1 and KAP8-1 genes, as fiber diameter is a key determinant of wool value. In this study, variation in the ovine KAP7-1 and KAP8-1 genes was investigated using polymerase chain reaction–single-stranded conformational polymorphism (PCR–SSCP) analysis.
Materials and Methods
Sheep investigated and DNA isolation
Two hundred fifty Romney-cross sheep from New Zealand were investigated. These are the most common ewe-breed in the New Zealand sheep industry, and encompass the NZ Romney, Perendale (Romney×Cheviot) and Coopworth (Romney×Border Leicester) breeds. Blood samples were collected onto FTA cards (Whatman BioScience, Middlesex, United kingdom) and genomic DNA was purified using a two-step washing procedure as described in Zhou et al. (2006).
PCR primers and amplification
KRTAP7-1 PCR primers (KRTAP7-up: 5′ ACTTGCTCTTC ACATTCTATC 3′ and KRTAP7-dn: 5′ GTAGTCATCTGGA GCCATG 3′) were designed based on the published ovine KRTAP7-1 sequence (Genbank X05638; Kuczek and Roger, 1987), and the published ovine KRTAP8-1 sequence (GenBank X05639; Kuczek and Roger, 1987) was used to design the KRTAP8-1 PCR primers (KRTAP8-up: 5′ CATTCCCTG CTCTCCAAGC 3′ and KRTAP8-dn: 5′ GAGAAGATTCC ATGCCTCTG 3′). The primers spanned the entire coding region of the gene. Primers were synthesized by Integrated DNA Technologies (Coralville, IA).
Amplification was performed in a 20-μL reaction containing the genomic DNA on one 1.2-mm punch of FTA paper, 0.25 μM of each primer, 150 μM of each dNTP (Eppendorf, Hamburg, Germany), 2.5 mM of MgCl2, 0.5 U of Taq DNA polymerase (Qiagen, Hilden, Germany), and 1X the reaction buffer supplied. The thermal profile consisted of 2 min at 94°C, followed by 35 cycles of 30 s at 94°C, 30 s at 60°C, and 30 s at 72°C, with a final extension of 5 min at 72°C. Amplification was carried out in an iCycler (Bio-Rad, Hercules, CA).
Amplicons were observed by electrophoresis in 1% agarose (Quantum Scientific, Queensland, Australia) gels, using 1X TBE buffer (89 mM Tris, 89 mM boric acid, 2 mM Na2EDTA) containing 200 ng/mL of ethidium bromide.
SSCP analysis
A 0.7-μL aliquot of each amplicon was mixed with 7 μL of loading dye (98% formamide, 10 mM ethylenediaminetetraacetic acid [EDTA], 0.025% bromophenol blue, and 0.025% xylene-cyanol). After denaturation at 95 °C for 5 min, samples were rapidly cooled on wet ice and then loaded on 16 cm×18 cm, 14% acrylamide:bisacrylamide (37.5:1; Bio-Rad) gels. Electrophoresis was performed using Protean II xi cells (Bio-Rad), at 250 V for 18 h at 25°C in 0.5X TBE buffer. Gels were silver-stained according to the method of Byun et al. (2009).
Sequencing of allelic variants and sequence analysis
For each gene, PCR amplicons representative of different SSCP patterns from sheep that appeared to be homozygous were directly sequenced at the Lincoln University DNA Sequencing Facility. For those alleles that were only found in heterozygous sheep, they were sequenced using a rapid approach described previously (Gong et al., 2011). Briefly, an individual band corresponding to the allele was excised as a gel slice from the polyacrylamide gel, macerated, and then used as a template for reamplification with the original primers. This second amplicon was then directly sequenced.
Sequence alignments, translations, and comparisons were carried out using DNAMAN (version 5.2.10, Lynnon BioSoft, Vaudreuil, Canada). The BLAST algorithm was used to search the NCBI GenBank (
Results and Discussion
Variation in ovine KRTAP7-1
The PCR primers designed for ovine KRTAP7-1 produced PCR amplicons of the expected size (approximately 327 bp) with all the sheep DNA samples. These amplicons exhibited different banding patterns upon SSCP analysis and two unique banding patterns could be detected (Fig. 1a). Sequencing of the amplicons revealed that these two unique patterns represented DNA sequences that had a single-nucleotide difference (c.173G/A). One of the sequences was identical to the published ovine KRTAP7-1 sequence (GenBankX05638; Kuczek and Roger, 1987) and named KRTAP7-1*A, whereas the other was novel and named KRTAP7-1*B. This suggests that these two sequences represent allelic variants of ovine KRTAP7-1. The nucleotide substitution detected in ovine KRTAP7-1 was nonsynonymous and would result in an amino acid change (p.Ser58Asn) if the gene is expressed. These allelic sequences have been deposited into the NCBI GenBank with accession nos. JN091630 and JN091631. The frequencies of KRTAP7-1*A and KRTAP7-1*B in the sheep investigated were 77% and 23%, respectively.
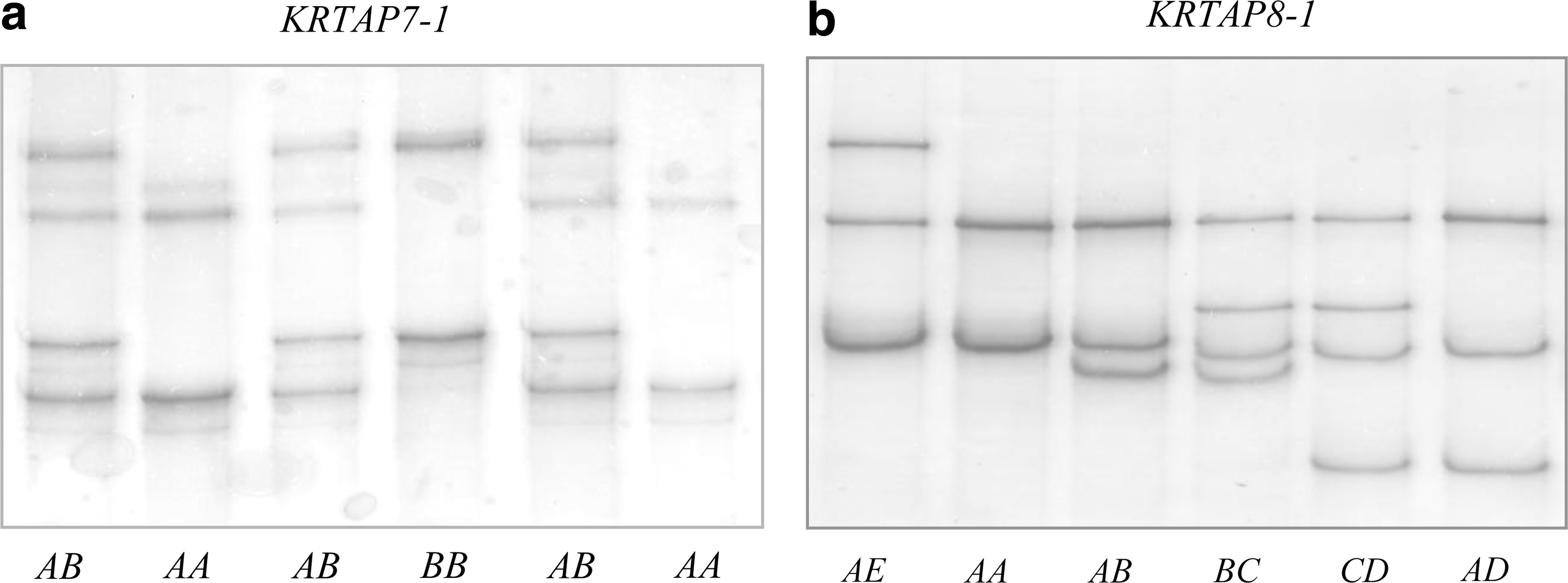
Polymerase chain reaction–single-stranded conformational polymorphism (PCR–SSCP) of ovine KRTAP7-1 and KRTAP8-1. Sheep representative of the unique PCR–SSCP patterns corresponding to
Only two alleles were detected in ovine KRTAP7-1, and SSCP with different electrophoresis conditions did not reveal any further variation. The extent of variation detected here was somewhat lower than that reported by McLaren et al. (1997) using Southern hybridization–RFLP. They reported two alleles using the restriction enzyme BglII and four alleles using MspI in analyzing three full-sib pedigrees of the AgResearch International Mapping Flock (Crawford et al., 1995). However, the sequence variation revealed here was not in either a nominal BglII or MspI recognition site. What is more, there are no BglII recognition sequences present or that can be created by a single-nucleotide substitution, in the coding region of ovine KRTAP7-1. This suggests that these recognition sites were located outside of the coding region of the gene, and hence the variation revealed by Southern hybridization–RFLP was in regions flanking the gene. If this was the case, then one would expect only simple RFLP patterns when using a gene-specific probe, although this was not discussed by the authors. It is also possible that the discrepancy between our study and McLaren et al.'s (1997) may be due to different samples being investigated. Nevertheless, this study shows that PCR–SSCP detected sequence variation in the ovine KRTAP7-1 coding region that cannot be detected by Southern hybridization–RFLP in a previous study, as the latter is incapable of detecting sequence substitutions located outside of the restriction endonuclease recognition sites.
Variation in ovine KRTAP8-1
PCR amplification using KRTAP8-1 primers generated amplicons of the expected size (approximately 258 bp) with all the sheep DNA samples. These amplicons displayed different banding patterns on SSCP gels and five unique banding patterns were detected (Fig. 1b). One or a combination of two banding patterns was observed for each individual sheep, which is consistent with them being either homozygous or heterozygous at this locus. Sequencing of the amplicons revealed five different DNA sequences. All of the sequences were highly homologous to, but none of them was identical to, the published ovine KRTAP8-1 sequence (GenBank X05639; Kuczek and Rogers, 1987), suggesting that these sequences represent allelic variants of ovine KRTAP8-1 (Fig. 2). These alleles were named KRTAP8-1*A to KRTAP8-1*E, and the sequences were deposited into the NCBI GenBank with accession numbers JN091632–JN091636. Alleles A to E were detected at frequencies of 68.3%, 13.3%, 3.3%, 10.0%, and 5.0%, respectively, in the sheep investigated.

Sequence alignment of the ovine KRTAP8-1 alleles (A to E) identified in this study with the published KRTAP8-1 sequence (GenBank X05639). Nucleotides in the coding region are shown in upper-case and amino acids are represented in one-letter code and shown in bold. Nucleotides identical to the published sequence X05639 are represented by hyphens, and amino acid sequences are shown under the nucleotide sequences if different to the transcript of X05536, which is shown. The stop codon is indicated by *.
The variation reported here in the KRTAP8-1 coding region appears to be less than that reported by Wood et al. (1992), who reported 11 KRTAP8-1 alleles in 33 unrelated sheep from various breeds (including the NZ Romney, Coopworth, and Perendale breeds, and various hybrids of these breeds) based on dinucleotide length variation in PCR amplicons of the 5′ flanking region of the gene. In that study, the PCR primers were designed based on the published ovine KRTAP8-1 sequence (GenBank X05639; Kuczek and Rogers, 1987) and were expected to amplify an amplicon of 124 bp. However, the PCR amplicons reported by Wood et al. (1992) were 152–175 bp long, and did not match the expected size (124 bp). There was no sequencing data reported for the amplicons by these authors and accordingly the sequence of the nominal 5′ flanking region variation awaits validation.
In this study, four single-nucleotide substitutions were identified in ovine KRTAP8-1 and three of them were located in the coding region. Of these, only one substitution was nonsynonymous and it would result in an amino acid change (p. Tyr34Asn). It should be noted, however, that silent nucleotide substitutions may also influence gene expression (Duan et al., 2003) and accordingly all the variation detected here may affect the structure and/or expression of KAP8-1, as may that variation reported by Wood et al. (1992). In both cases this genetic variation may affect wool traits.
It is notable that all five KRTAP8 sequences have different nucleotides to the GenBank sequence (X05639) at positions c.48 (T >C) and c.152 (C >T), the later causing a Ser/Phe amino acid substitution although this is unlikely to affect the potential N-glycosylation site that is nearby. This may reflect the different breeds of sheep studied in this research.
Footnotes
Acknowledgments
This work was financially supported by FRST (C10X0710: Keeping New Zealand Wool Products at the Cutting Edge Through Enhanced Wool Quality) and the Lincoln University Gene-Marker Laboratory. The Wool Research Organisation of New Zealand Inc and New Zealand Wool Industry Charitable Trust Postgraduate Scholarship to H.G. is acknowledged.
Disclosure Statement
No competing financial interests exist.