
Abstract
The distinguishable morphologic features of nuclei of acute myelogenous leukemia cells with enlarged size and finely distributed nuclear chromatin indicate incomplete chromosome condensation that can be related to elevated gene expression. To confirm this, interphase chromosome structures were studied in exponentially growing rat myelomonocytic leukemia 1 cells isolated at the University of Debrecen (My1/De cells). This cell line was established from primary rat leukemia chemically induced by 7,12-dimethylbenz[a]anthracene treatment. The enlarged nuclei of My1/De cells allowed improved fluorescent visualization of chromosomal structures. Increased resolution revealed major interphase intermediates consisting of (1) veil-like chromatin, (2) chromatin ribbon, (3) chromatin funnel, (4) chromatin bodies, (5) elongated prechromosomes, (6) seal-ring, spiral shaped, and circular chromosomal subunits, (7) elongated, bent, u- and v-shaped prechromosomes, and (8) metaphase chromosomes. Results confirmed the existence of the chromatin funnel, the first visible interphase chromosome generated by the supercoiling of the chromatin ribbon. Other intermediates not seen previously included the spiral subunits that are involved in the chromonemic folding of metaphase chromosomes. The existence of spiral subunits favors the helical coil model of chromosome condensation. Incomplete chromatin condensation in leukemia cells throughout the cell cycle is an indication of euchromatization contributing to enhanced gene expression and is regarded as a leukemic factor.
Introduction
Morphological features are still the most important criteria for diagnosing different types of leukemia, de novo acute erythroid leukemia occurring more frequently than pure erythroid leukemia (Domingo-Claros et al., 2002). To study in vitro chromosome morphology, chemically induced rat erythroleukemia cells have been established (Maeda et al., 1980). Intravenous injections of 7,12-dimethylbenz[a]anthracene (DMBA), a neoplastic agent, induced erythroblastic leukemia with high frequency of #2 trisomy in Long-Evans rat (Osaka et al., 1995). These leukemias revealed a highly specific mutation of the N-ras gene similar to that in human leukemias and provided an important target for the study of chemical carcinogenesis and leukemia (Sugiyama et al., 2002). Granulocytic leukemia induced in Long-Evans rats using the Sugiyama method is an ideal model, which obtains solid tumor growth and myelomonocytic leukemias equivalent to human M4-M5-type leukemia (leukemia genetic rearrangements associated with French-American-British Classification M4-M5) (Kozma et al., 1993). Genetic rearrangements in patients with AML were significantly associated with French-American-British Classification M5, high number of white blood cells, and young age (Cimino et al., 1995).
To analyze interphase chromosomal structures, we used reversibly permeabilized cells (Banfalvi et al., 1984) and visualized them by fluorescent microscopy (Banfalvi et al., 2006; Banfalvi, 2009). To demonstrate the temporal order of chromosome condensation, intermediates were isolated at different stages of the cell cycle from cells synchronized by counterflow centrifugal elutriation (Banfalvi, 2008a). As a result of synchronization, major intermediates of chromosome condensation have been described in four mammalian cell lines (Banfalvi et al., 2006; Banfalvi, 2009), in Drosophila cells (Banfalvi et al., 2007), and under in vivo conditions in rat hepatocarcinoma primary tumor cells (Trencsenyi et al., 2007).
Myelomonocytic leukemias (chronic or juvenile) are hematological disorders, characterized by increased cell size in the bone marrow and peripheral blood. Because of the common dysplastic changes, the enormously enlarged nuclei (average diameter: 12 μm) of the rat myelomonocytes (myelomonocytic leukemia 1 isolated at the University of Debrecen [My1/De]; diameter: 20–25 μm) was chosen to visualize structural chromatin transitions during the interphase. In this study, we used these extremely enlarged nuclei of myelomonocytic leukemia cells to confirm the existence of large-scale chromosomal changes seen before in other mammalian cells. The enlarged size allowed us to visualize two new intermediates of interphase chromosomes, namely, the chromatin funnel appearing in mid-S phase and the small spiral unit of chromosome condensation appearing at the end of the condensation process. The incomplete chromatin condensation could relate to the proliferation and elevated gene expression in myeloid leukemia and may serve as an important diagnostic tool.
Materials and Methods
Chemicals and solutions
1,4-Diazobicyclo-(2,2,2)-octane (DABCO) was obtained from Sigma (St. Louis, MO). 2,6-Diamino-2-phenylindole (DAPI) was the product of Braunschweig Chemie (Braunschweig, Germany). Tween 20 was purchased from Pierce (Rockford, IL). Dextran T-150 was purchased from Pharmacia-Biochemicals (Uppsala, Sweden). Colcemid (N-methyl-N-deacetyl-colchicine) was the product of Boehringer (Mannheim, Germany). Fetal bovine serum was obtained from GIBCO BRL, Life Technologies (Gaithersburg, MD). 7,12-Dimethylbenz[a]anthracene (DMBA; C20H16; M w: 256.36) and other chemicals were obtained from Sigma-Aldrich (Budapest, Hungary).
Antifade medium consisted of 90% glycerol, 2% (w/w) DABCO, 20 mM Tris-Cl, pH 8.0, 0.02% sodium azide, and 25 ng/mL DAPI for blue fluorescent total staining of DNA. Hypotonic buffer for reversible permeabilization contained 9 mM HEPES (pH 7.8), 5.8 mM dithiothreitol, 4.5% dextran T-150, 1 mM EGTA, and 4.5 mM MgCl2. Swelling buffer consisted of 50 mM KCl, 10 mM MgSO4, 3 mM dithiothreitol, and 5 mM NaPO4 (pH 8.0). Fixative solution contained methanol:glacial acetic acid (3:1).
Animals
Female Long-Evans (LE) rats were kept in a conventional laboratory environment and fed on a semisynthetic diet (Charles River Mo, Kft, Godollo, Hungary) and tap water ad libitum. Animals received human care according to the criteria outlined in the Guide for the Care and Use of Laboratory Animals authorized by the Ethical Committee for Animal Research, University of Debrecen, Hungary (permission number: 22/2007). The NIH Guidelines for the Laboratory Use of Chemical Carcinogens were implemented during the operations and manipulations involving DMBA.
Isolation of myelomonocytic cells
Myelomonocytic leukemia was originally induced in rats by DMBA (Kozma et al., 1993). Cancer development in rats by intravenous administration of an emulsion of DMBA (Huggins and Sugiyama, 1966; Huggins et al., 1970) has been widely used to elucidate mammary carcinogenesis (Huggins and Yang, 1962; Jordan, 1987; Russo and Russo, 1996) because of the simplicity of the procedure. My1/De cells originated from the recipient transformed cells obtained after several passages of cells primarily isolated from DMBA-treated rats.
Establishment of the My1/De leukemia cell culture
After serial passage, the cells of granulocytic leukemia isolated from rats became transformed. The newly transformed cells could be transplanted to LE rats. A quantity of 108 cells/100 g body weight was intravenously injected into LE rats, and after 2–3 weeks, myelomonocytic leukemia developed (Kozma et al., 1993). Primary cell culture was started from the bone marrow of myelomonocytic leukemia-bearing rats ex vivo as a suspension culture. The cells were resuspended in RPMI 1640 medium (culture medium developed in Roswell Park Memorial Institute) supplemented with 10% fetal bovine serum and antibiotics. The primary cell culture of myelomonocytes isolated from leukemic rats induced by DMBA was grown further, and after 20 subculturings the new cell culture referred to as My1/De was established. My1/De cell cultures originating from the bone marrow of leukemic Long-Evans rats can be maintained for 2–2.5 months under in vitro conditions. After 60 days, the cells differentiated to macrophages.
Reversible permeabilization
This method was originally developed for the study of nascent DNA synthesis in reversibly permeabilized lymphocytes isolated from the murine thymus (Banfalvi et al., 1984). It was found that reversible permeabilization allowed to open the nucleus and the visualize chromatin structures any time during the cell cycle (Banfalvi, 1993). This procedure was adapted to My1/De cells. Briefly, 1 mL of hypotonic buffer was added to 106 cells in the presence of Dextran T-150 as a molecular coat to prevent cells from disruption. Permeabilization lasted for 2 min at 0°C. For reversal of permeabilization, the hypotonic solution was replaced by RPMI 1640 medium containing 10% fetal bovine serum, and the cells were incubated in a CO2 incubator at 37°C in the presence of 5% CO2 for 3 h.
Isolation of nuclei
On account of the cyclic character of chromatin unfolding and chromosome condensation, after reversal of permeabilization, the cells were treated with colcemid to prevent them from entering a new cell cycle (Banfalvi et al., 1984). After reversal of permeabilization, the cells (106) were resuspended in growth medium and treated with 0.1 μg/mL colcemid for 2 h at 37°C under 5% CO2. Monolayer cells were washed twice with phosphate-buffered saline (PBS).
Trypsinization of adherent cells
After the removal of PBS, a minimal volume (i.e., 0.9 mL for a T-75 flask) of 0.25% (w/v) trypsin solution (prewarmed to 37°C) was added. The cells were incubated for ∼3 min, and then the flask was checked under inverted microscope to make sure that the monolayer of cells was lifting off of the flask. The cell suspension was transferred to a sterile 15-mL conical centrifuge tube. The cells were pelleted at 500 g for 5 min in a tabletop centrifuge. The supernatant was carefully discarded so as to not disturb the cell pellet.
Determination of viable cell number
Cells were resuspended in 5 mL PBS, and 50 μL cell suspension was mixed with 50 μL of 0.5% trypan blue dissolved in PBS. Viable cells that did not take up trypan blue were counted in a Bürker chamber. Once the number of cells per milliliter has been determined, 106 cells were taken for osmotic swelling.
Osmotic cell swelling
Cells (106) were washed twice with PBS and incubated at 37°C for 10 min in swelling buffer. Swelling was followed by centrifugation at 500 g for 5 min.
Isolation of nuclei
After the swelling, nuclei were isolated from the pellet of cells by slow addition under constant stirring of 14 mL fixative solution. Nuclei were then centrifuged at 500 g for 5 min and washed twice in fixative.
Spreads of nuclear structures
Preparation of nuclei for spreads of chromatin structures was done using the method developed for metaphase chromosomes. Nuclei were centrifuged at 500 g for 5 min and resuspended in 1 mL fixative. Nuclei were spread over glass slides dropwise from a height of approximately 30 cm. Slides were air dried, stored at room temperature overnight, rinsed with PBS, and dehydrated using increasing concentrations of ethanol (70%, 90%, 95%, and 100%).
Visualization of chromatin structures
Dehydrated slides containing chromatin structures were mounted in 35 μL antifade medium under 24×50 mm coverslips. Blue fluorescence of DAPI was monitored by fluorescence microscopy (Nikon Eclipse E800 Nikon Corporation, Tokyo, Japan).
Results
Intermediates of chromosome condensation in nuclei of My1/De leukemia cells
To ensure that the overwhelming majority of cells are in interphase, logarithmic growth was maintained by daily subculturing 3 days before and during the experiments. My1/De cells (106) were grown for 15 h in X-VIVO™ 20 medium containing 10% FBS in a CO2 incubator. Reversible permeabilization was used to open the nucleus any time during the cell cycle (Banfalvi et al., 1984, 2006).
As the field of chromosome condensation has a long history of artifacts, one could argue that reversible permeabilization may suffer from same sort of handicap. The best evidence that we obtained real intermediates was provided by finding the same intermediates using different mammalian cells (Chinese hamster ovary [CHO], Indian muntjac, murine preB, human K562, rat liver), reflecting a common pathway of condensation (Banfalvi et al., 2006). To the contrary, upon genotoxic treatment (Cd, gamma, and UVB irradiation), the chromatin structures were dramatically changed with most of the chromosomes unable to reach metaphase (Nagy et al., 2004; Banfalvi et al., 2005; Ujvarosi et al., 2007).
The most characteristic chromatin structures isolated from My1/De leukemia cells were veil-like fibrillary structures, supercoiled ribbons, round fibrous chromatin bodies, and precondensed chromosomal forms (Fig. 1). Major difference relative to other mammalian cells was that My1/De chromatin structures were more decondensed throughout the cell cycle. The nuclei of cells readily opened in their decondensed state, making the fine chromatin veil visible as a faint halo (Fig. 1a–h). Arrows show where the chromatin veil is turned to funnel-shaped forms (Fig. 1e–j). A characteristic feature of chromatin folding is the supercoiling of chromatin veil to ribbon (Fig. 1j–s). Further supercoiling of ribbon generates loops, which turn to funnel-shaped structures, the first visible chromosomal form. The third important structural manifestation during chromatin condensation was the tightening of chromatin funnels to round chromatin bodies, which were regarded earlier as the first visible forms of interphase chromosomes (Fig. 1s–u). By tightening the funnel-shaped forms into round-shaped chromatin bodies and by linearization, ring-shaped and seal-ring-shaped forms will be generated. Chromatin bodies are condensed and unknotted to linear precondensed chromosomal forms (Fig. 1v, w) and occasionally reach metaphase (Fig. 1x, y). Figure 1x clearly shows the final linear arrangement of chromosomes.
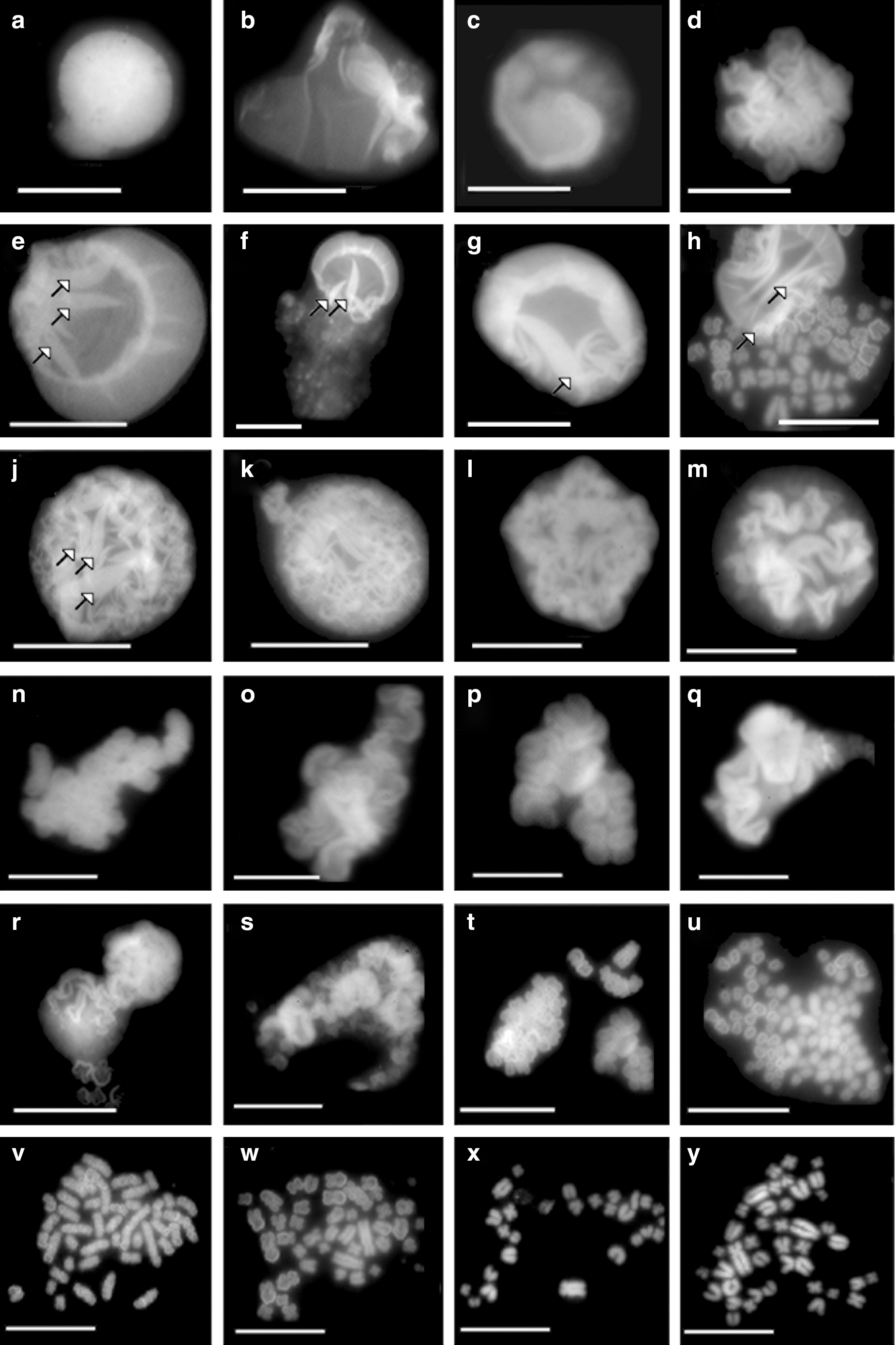
Chromatin structures of control My1/De leukemia cells. Isolation of nuclei and chromatin structures was carried out as described in the Materials and Methods. Chromatin structures with increasing compaction: highly decondensed chromatin veil
Chromatin funnel, an early chromosomal form
Figure 1h was magnified for closer scrutiny to confirm the formation of a new intermediate of the interphase chromosome called chromatin funnel after its shape. Chromatin funnels are indicated by the two arrows in Figure 2. Funnel-shaped structures were indicated by the arrows in Figure 1e–j. The chromatin funnel is generated by the supercoiling of the chromatin ribbon. Common intermediates of supranucleosomal chromosome condensation have been described earlier in four mammalian cells. These large-scale chromatin structures included chromatin veil, chromatin ribbon, fibrous chromatin, chromatin body, precondensed elongated chromosomes, and metaphase chromosomes (Banfalvi et al., 2006). The chromatin funnel was recently found in human K562 erythroleukemia cells (Banfalvi and Nagy, 2011). It is a newly revealed intermediate of chromatin condensation and the first distinguishable form of chromosomes.
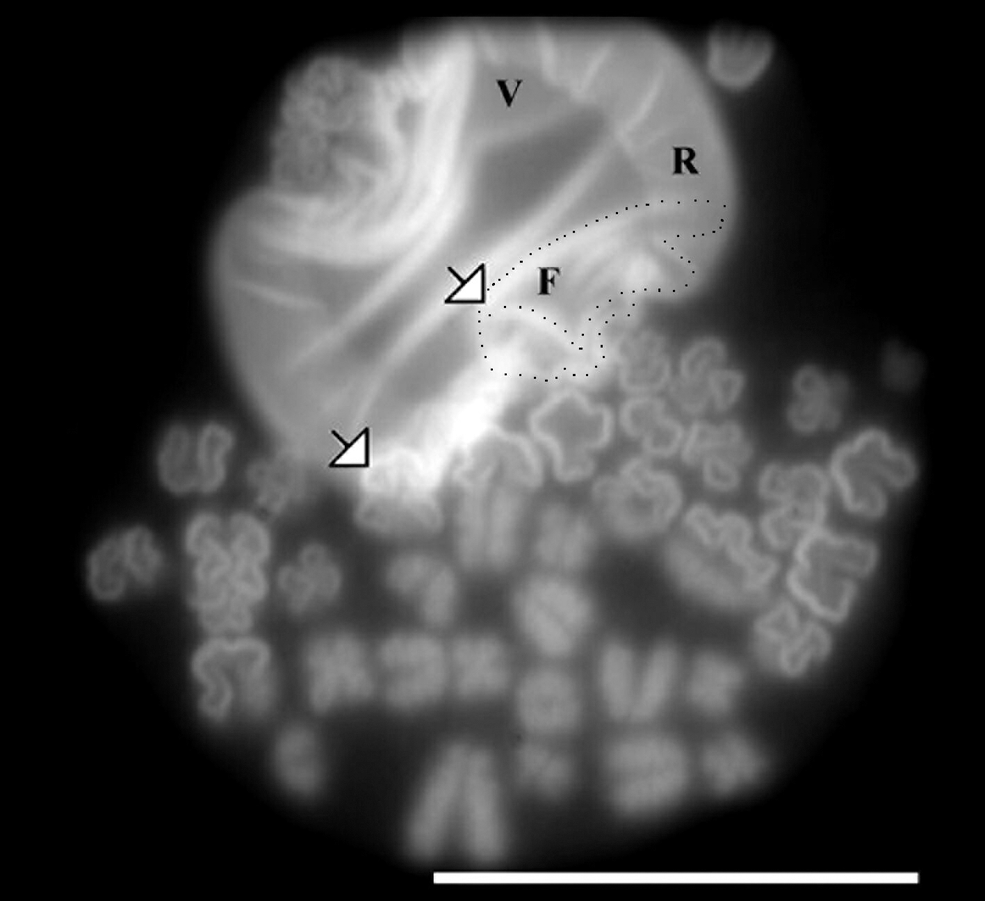
Formation of chromatin funnel from ribboned chromatin. Isolation of nuclei and chromatin structures was carried out as described in the Materials and Methods. Arrows show funnel-shaped chromatin structures. F: funnel; R: ribbon; V: veil. One chromatin funnel is outlined by black dots. Bar: 10 μm.
Spiral units of metaphase chromosomes
The linearization of round-shaped CHO chromosomes taking place around their flexible hinge regions suggested earlier that smaller units of mammalian chromosomes may exist (Banfalvi et al., 2006). The relatively uniform size and large number of chromatin bodies exceeding the diploid chromosome number (42) of rat myelocytes (Fig. 1u) seem to confirm this notion. Earlier we observed smaller chromosome subunits in Drosophila cells and called them chromosome rodlets (Banfalvi et al., 2007). Similar smaller units in leukemia My1/De cells are demonstrated in Figure 3, with arrows pointing to the spiral forms. Figure 3a is a magnified view of Figure 1v. The uniform size and large number of spirals is an indication that they are not chromosomes, but smaller chromosomal subunits. The size of individual chromosomes seems to depend on the number of spiral units.
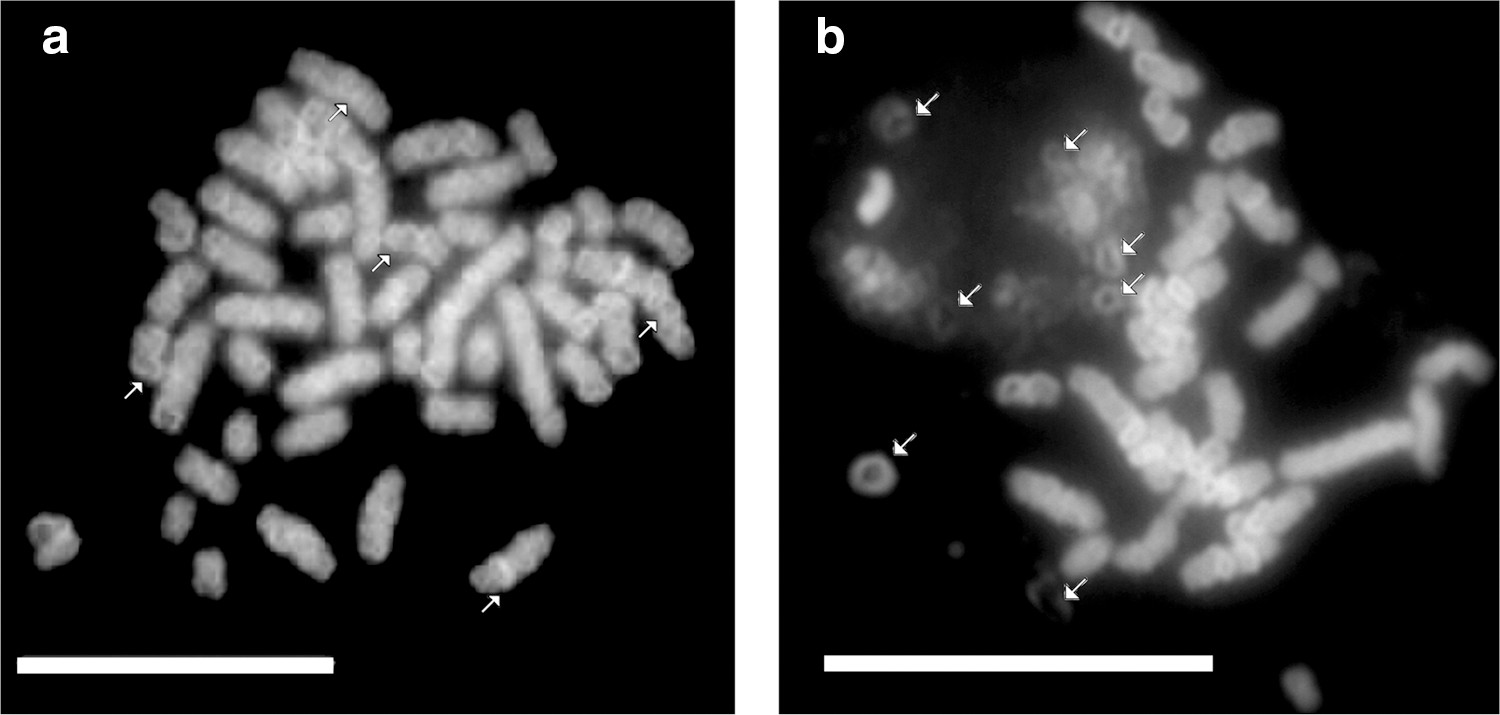
Linear chromosomes composed of spiral subunits. Isolation of nuclei and chromatin structures was carried out as described in the Materials and Methods.
Multiple stages of chromosome formation
Chromosome condensation involves several intermediates, some of them may be not yet known. That this could be the case is indicated by Figure 4a, showing the content of three nuclei, two of them maintaining the round shape of the nucleus and the third one consisting of different transitory chromosomal forms. Because of the general tendency of the lower compaction of chromatin in My1/De cells, these transitory forms differ slightly from one another, and consequently, their categorization can be only arbitrary. To distinguish them, we have selected four groups of condensing chromosomes (Fig. 4b). The upper row of Figure 4b contains closed, but not circular, chromosomal forms, resembling enlarged chromatin bodies, which are early visible chromosomes (similar to two closed fists); in the second row, open u- and v-shaped chromosomes (resembling two opening fists) have been collected; the third row consists of coiled, bent chromosomal forms; and the last row contains those chromosomes that approach metaphase. The most compact and distinguishable supercoiled forms seen at the right end of each row in Figure 4b are represented by spiral chromosomes that obviously did not reach the compactness of metaphase chromosomes. These supercoiled forms indicated by asterisks have been selected and are separately shown in Figure 4c. The condensed forms demonstrate that supercoiling takes place around the helical axis. The magnification of Figure 1x and z has also revealed spiral forms (not shown).
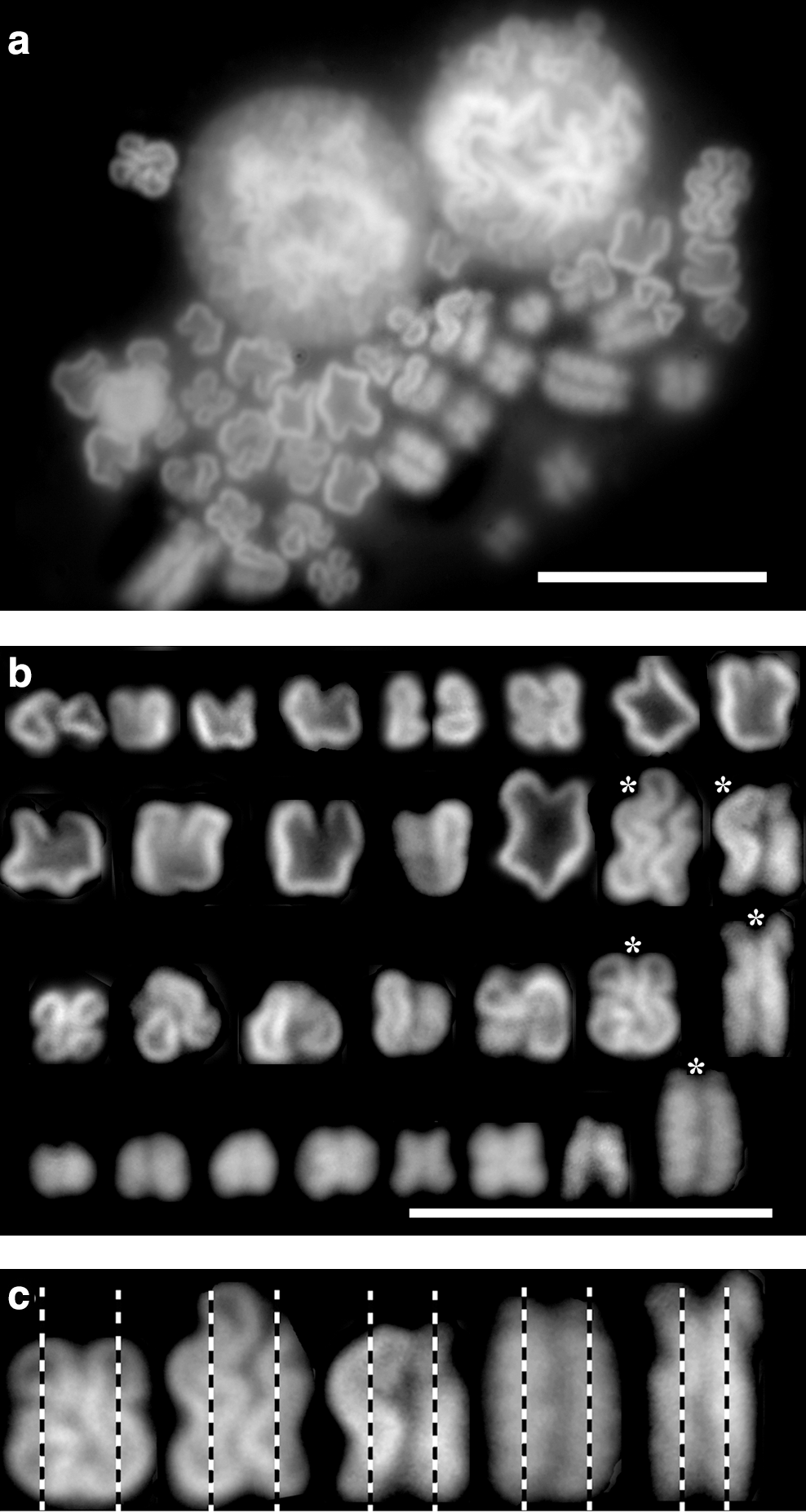
Final stage of chromosome condensation.
Discussion
Granulocytic leukemia was induced in rats after treatment with the carcinogen DMBA (Kozma et al., 1993). The myelomonocytic leukemia cell culture (My1/De) was established from transformed cells of the tumor-bearing rat. Because of the dysplastic changes in size, the enormously enlarged nuclei of logarithmically growing My1/De myelomonocytic cells have been chosen to identify intermediates of interphase chromosomal structures. Typical chromatin structures isolated from control, untreated leukemia cells were identical with those isolated earlier from other mammalian cells, including decondensed fibrillary chromatin-veil, supercoiled chromatin, fibrous chromatin, ribboned chromatin, round chromatin bodies, early bent precondensed, elongated prechromosomes, and highly condensed metaphase chromosomal forms (Banfalvi et al., 2006). The new chromatin structure not observed earlier has been named chromatin funnel. Turning the chromatin ribbon around itself forms a loop, which will form the funnel. By tightening the loop, round-shaped chromatin bodies will be generated. The reason that the chromatin funnel could not be visualized earlier is due to the fact that the average size of nuclei of mammalian cells used for the analysis (CHO, Indian muntjac, murine preB, K562 erythroleukemia) was much smaller (5–10 μm) than that of My1/De cells (10–15 μm). The large nucleus of My1/De cells providing better resolution of nuclear structures enabled the discovery of this early visible chromosomal form. One could argue that the extreme enlargement of My1/De nuclei would not preserve chromatin structures. However, after chemical carcinogenesis induced with n-nitrosodimethylamine, the chromatin structure of rat hepatocarcinoma tumor cells (hepatocarcinoma tumor isolated at the University of Debrecen [He/De]) was similar to normal cells. The exception similarly to human malignancies (Bronstein et al., 2006) was that in He/De cells the tendency of supercoiling was higher throughout the cell cycle, suggesting that increased supercoiling may contribute to the reduced length of cell cycle of tumor cells (Trencsenyi et al., 2007). The similar feature observed during the chromatin folding of My1/De leukemia cells seems to confirm this notion.
Another new observation was the larger than expected number of round chromatin bodies turning to spiral forms, indicating that chromosomes may consist of smaller units. At this stage of study, we would neither exclude nor confirm the possibility that such subunits of chromosomes are parts of chromatin bodies. Small chromosomal subunits named rodlets were seen in Drosophila cells (Banfalvi, 2008b). However, the shape and form of the Drosophila rodlets are not similar to the chromatin spirals of My1/De cells. Rather, the spiral forms are likely part of the supercoiled thick fibers. Contrary to Drosophila cells, these supercoiled units do not resemble rodlets, but are round forms. Confirming these observations, it was suggested by others that higher-order chromatin fibers named chromonemas (100 nm) could consist of 30-nm fibrils by successive folding into progressively thicker rodlets and could be functional intermediates. This folding mechanism in Drosophila contradicted the radial-loop model and provided evidence for the hierarchical model of chromosome architecture (Dietzel and Belmont, 2001; Strukov et al., 2003).
Mitotic chromosomes are orderly and reproducibly packaged, but the supranucleosomal organization above the 10-nm “beads on string” chromatin fibril remained poorly characterized (Gassmann et al., 2004; Hozak and Fakan, 2006). The mechanisms underlying chromatin condensation represent a long-standing unresolved problem exemplified by several models of mitotic chromosome formation. Depending on experimental approaches, the structural analysis of fully compact, native chromosomes led to at least four models for the metaphase chromosome architecture. These models are known as (1) radial-loop (Paulson and Laemmli, 1977; Laemmli et al., 1978; Marsden and Laemmli, 1979), and a variation of the radial-loop model named radial-loop/helical coil model (Rattner and Linn, 1985; Boy De La Tour and Laemmli, 1988), (2) hierarchical folding and the two variants of this group of models (Sedat and Manuelidis, 1978; Belmont et al., 1987; Belmont and Bruce, 1994), (3) the network model (Poirier and Marko, 2002; Poirier et al., 2009), and (4) the axial glue mechanism (Kireeva et al., 2004). The spiral forms we have observed at the end of chromosome condensation in My1/De cells suggest that they are wound around the nonhistone scaffold protein axis. The scaffold core implies that the chromosome is regularly compacted along the long axis of the chromosome. Originally, nuclear scaffolding in the nucleus was proposed by Flemming (1882) and was developed as the radial-loop model by Laemmli et al. (1978). The large-scale structure of mitotic chromatids remained controversial (Koshland and Strunnikov, 1995; Strunnikov, 1998; Hirano, 2000), as indicated by further studies favoring the folding of chromatin loops onto a central protein axis (Earnshaw and Laemmli, 1983). Other studies suggested successive levels of helical folding (Belmont et al., 1987). This controversy could be resolved by assuming that chromosome condensation in higher eukaryotes (mammalian cells) follows a radial-loop/helical coil mechanism, whereas in lower eukaryotes (e.g., Drosophila) the rodlets are the representatives of helical (plectonemic) folding. This assumption is based on the view regarding the helical (plectonemic) chromatin folding in Drosophila cells (Banfalvi, 2008b) and on experiments with My1/De leukemia cells favoring the helical coil mechanism of chromosome condensation.
Regarding the mechanism of chromosome condensation, we know only the diameters of chromatin structures at increasing compaction. The double helical DNA turned around the octameric histone core creates disc-shaped nucleosomes of 11 nm diameter, the attachment of which is known as “beads on string” or chromatin string. Nucleosomes are packed together to form the 30-nm chromatin fiber, whose exact structure is not known. The 30-nm fiber is thought to form chromatin loops of an estimated 300-nm-diameter. These loops turn around the nonhistone protein core or scaffold (Jackson, 1991), bringing about the 600–700-nm condensed chromatin. The helical coiling is favored over axial-loop folding schematically shown in Figure 5. Scaffold proteins are essential for several aspects of chromosomal behavior (Balczon, 1993). The estimated diameter of the scaffold proteins is about 150 nm. The existence of lampbrush chromosomes during the meiosis of amphibian oogenesis supports the concept that chromatin fibers are organized in a series of loops. The condensation of chromatin loops takes place by an unknown mechanism. Of the two coiling possibilities, the relatively homogeneous distribution of thick chromatin fibers favors the helical coiling mechanism over the axial-loop folding (Fig. 5). This idea is consistent with the view that macroscopic folding of homogeneously stained regions in late S phase leads to the appearance of bands related to replication that are close to the G/R bands in mitotic chromosomes (Shimizu and Shingaki, 2004).
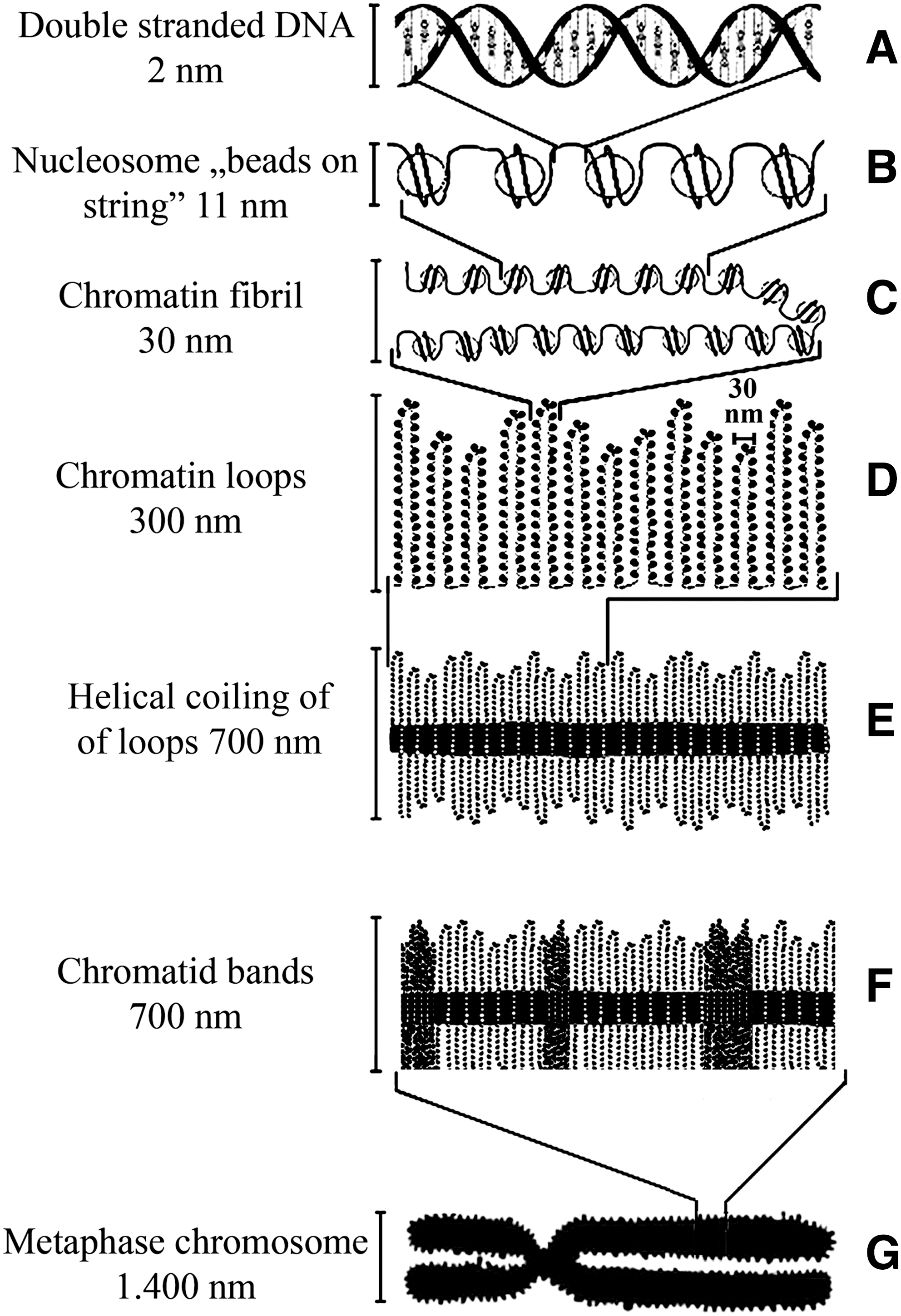
Helical-coiling model of chromosome condensation.
We expected that large nuclei of cells offer a better microscopic resolution to visualize large-scale chromosomal changes. For this purpose we have established a monomyelocytic leukemia cell culture (My1/De; average nuclear size: 10–15 μm) by chemical carcinogenesis. The isolation of large nuclei of exponentially growing My1/De leukemia cells resulted in the visualization of two new intermediates of large-scale chromosome structures, namely, chromatin funnels and spiral units. Chromatin funnel represents the missing link between chromatin ribbon and chromatin bodies and is the earliest visible chromosomal form in the process of chromatin condensation. The process of chromatin condensation is clearly visible in K562 cells, showing the formation from chromatin ribbon to funnel to chromatin body (Banfalvi and Nagy, 2011). The existence of spiral units allowed us to turn from earlier theoretical considerations to the suggestion based on experimental evidence, that is, the thick chromatin fiber (300 nm) is turned around the nuclear scaffold, favoring the helical coil mechanism of chromosome formation.
Finally, one can propose the relationship between decondensed chromatin structures and elevated gene expression in AML cells. As chromatin structures are less condensed in each stage of the condensation process, nuclei of leukemia cells are enlarged. Decondensed chromatin implies the expression of euchromatized genes in AML cells that were silent in normal cells. The expression of euchromatized genes is regarded as the major cause of carcinogenesis in acute leukemia and myelogenous leukemia. As the gene expression is associated with morphological changes, the fluorescent visualization of decondensed chromatin structures in AML cells with relative ease is of diagnostic importance.
Footnotes
Acknowledgment
This work was supported by Hungarian Scientific Research Fund (OTKA grant T42762 to G.B.).
Disclosure Statement
No competing financial interests exist.