
Abstract
The mammalian target of rapamycin (mTOR) is an evolutionarily conserved protein kinase that belongs to the phosphatidylinositol kinase-related kinase family. We describe our molecular characterization of mTOR and its function (GenBank accession HM114224) in Cashmere goat (Capra hircus). The goat mTOR complementary DNA is 8617 bp, comprising an open reading frame of 7650 bp—corresponding to a polypeptide of 2549 amino acids—and a 909 bp 3′ untranslated region with a polyA tract and a polyadenylation signal at nucleotides 8575–8580. In a bioinformatics analysis, goat mTOR has typical sites of activity and domains. mTOR mRNA was measured in brain, heart, testis, liver, spleen, kidney, and lung by real-time polymerase chain reaction, and the expression of mTOR in fetal fibroblasts was detected by western blot. The viability of fetal fibroblasts was inhibited on treatment with CCI-779, a specific inhibitor of mTOR. Our data supplied evidence that the transcription of mTOR was detected in the seven tissues in Cashmere goat, and mTOR protein was translated in fetal fibroblasts. The proliferation of fetal fibroblasts decreases on inhibition of mTOR.
Introduction
mTOR is a downstream target of Akt. PI3K activates many signaling molecules, of which the Akt/mTOR pathway is notable, due its function in inhibiting apoptosis and mediating proliferation (Aoki et al., 2001). mTOR is believed to regulate growth, and governs protein synthesis by phosphorylating downstream substrates, including p70S6 kinase (p70S6K) and eukaryotic initiation factor 4E-binding protein 1 (4E-BP1) (Burnett et al., 1998).
The mTOR signaling pathway has significant physiological and pathological functions in cancers and metabolic diseases (Schmelzle and Hall, 2000; Proud, 2002; Wullschleger et al., 2006). mTOR is active in diabetes (Fraenkel et al., 2008; Zoncu et al., 2010), obesity (Um et al., 2004; Chakraborty et al., 2010), the nervous system (Li et al., 2010; Neasta et al., 2010), aging (Selman et al., 2009; Kapahi et al., 2010) and in diverse immune cells (Powell and Delgoffe, 2010).
Rapamycin (sirolimus) is a macrolide antibiotic that is produced by Streptomyces hygroscopicus and binds FKBP-12 (FK506-binding protein). The rapamycin-FKBP12 complex inhibits mTOR activity. Rapamycin decreases the expression of phospho-mTOR, phospho-S6K, cyclin D1, and VEGF-A (Wu et al., 2010). Rapamycin also interferes with the G2/M transition in murine one-cell embryos by inhibiting the activity of M-phase promoting factor (MPF). The mTOR signaling pathway regulates the activity value of MPF in murine one-cell embryos (Xu et al., 2009). CCI-779 (temsirolimus), an analog of rapamycin, induces cell cycle arrest in G1 and apoptosis (Teachey et al., 2006; Gridelli et al., 2008) and has been developed as oral and intravenous formulations for clinical use.
The mTOR gene and protein have been extensively examined in human, mouse, and rat but not in goat, due to the lack of basic data in this animal. To study the function and regulation of mTOR in Inner Mongolia Cashmere goat cells, we cloned full-length mTOR complementary DNA (cDNA), measured its transcription in various tissues by quantitative real-time polymerase chain reaction (PCR), and investigated its function in goat cell growth.
Materials and Methods
Animals and tissue collection
Inner Mongolia cashmere goats were bred on a natural diet in Inner Mongolia, China. Brain, heart, testis, liver, spleen, kidney, and lung were collected from five adult male goats after slaughter in a commercial goat slaughter farm in the spring. Tissue samples were flash frozen in liquid nitrogen immediately after harvesting and stored at −80°C.
Cell cultures
Inner Mongolia cashmere goat fetal fibroblasts (GFbs) were cultured in DMEM/F12 (Gibco), supplemented with 10% fetal bovine serum (FBS; HyClone Laboratories, Inc.), 100 U/mL penicillin G, and 100 mg/mL streptomycin (Sigma-Aldrich, Inc.), and maintained in a monolayer culture at 37°C in humidified air with 5% CO2. Morphology was examined on a light microscope.
Reagents and antibodies
CCI-779 (temsirolimus), a bioavailable derivative of rapamycin, was synthesized by Wyeth Pharmaceuticals Inc. (Philadelphia) and provided by Dr. Naomoto, Okayama University, Japan. CCI-779, a TORISEL injection, 25 mg/mL was supplied with DILUENT for TORISEL, stored at 4°C, and diluted to an appropriate final concentration with culture media before use. The following primary antibodies were used: anti-β-actin (Sigma-Aldrich, Inc.) and anti-mTOR, a mouse serum polyclonal antibody that we raised against the C-terminal peptide of Cashmere goat mTOR kinase.
RNA extraction and cDNA synthesis
Total RNA was isolated using RNAzol (RNAiso Plus; TaKaRa Co. Ltd.,) from brain, heart, testis, liver, spleen, kidney, lung, and fetal fibroblasts of Inner Mongolia cashmere goat. RNA was reverse transcribed with an oligo (dT)12–18 primer using the AMV 1st Strand cDNA Synthesis kit (Takara Co. Ltd.) as per the manufacturer's instructions. cDNA from various tissues was subjected to quantitative real-time PCR, and full-length mTOR was cloned using cDNA from fetal fibroblasts. One microgram of total RNA was used for each reaction.
Cloning and sequencing of mTOR
Due to the length of mTOR, the coding sequence of mTOR was divided into three fragments for amplification. The predicted fragment length of the 5′ terminal fragment was 2218 bp; it was amplified with the following primers: forward: 5′ GAACCTCAGGGCAAGATGCTTGG 3′, reverse: 5′ TGAGCATCTTGCGCAGGAAAGG 3′. The 3′ terminal and the middle sections were 2886 bp and 2612 bp, respectively, primers for 3′ terminal fragment were as follows: forward: 5′ TGGTTTCTTGCCACATGCTGTCC 3′, reverse: 5′ CCAGTTACCAGAAAGGACACCAG 3′; and forward: 5′ CCTTTCCTGCGCAAGATGCTCATC 3′, reverse: 5′ TCGGACAGCATGTGGCAAGAAACC 3′ were primers for the middle section. Primers were designed using the sequence of homo sapiens mTOR in GenBank and commercially synthesized. These fragments were amplified for 35 cycles with cDNA as the template at various annealing temperatures (60°C, 55°C, 59.5°C). The PCR products were cloned into a plasmid and sequenced on an ABI PRISM 377XL DNA Sequencer (Applied Biosystems, Inc.).
3′ rapid amplification of cDNA ends
RNA was reverse transcribed with a 3′ rapid amplification of cDNA ends (3′ RACE) 1st strand cDNA synthesis kit and a 3′ RACE adaptor primer perthe manufacturer (Takara Co. Ltd.). The 3′ untranslated region (3′ UTR) fragment was amplified with cDNA as the template for 35 cycles at the appropriate annealing temperature (50.5°C) with the following primers: forward: 5′ CTGGATGTCCCAACACAAGTC 3′, reverse: 5′ CTGATCTAGAGGTACCGGATCC 3′. 3′ RACE generated a single fragment of 1070 bp.
Quantitative real-time PCR analysis of mTOR expression in various tissues
mTOR mRNA levels were measured by quantitative real-time PCR. mTOR cDNA was amplified with cDNA as the template by real-time PCR using the following primers: forward: 5′ CCCATTCACCAGTTTCCAGG 3′, reverse: 5′ CAGCGAGTTCTTGCTATTCC 3′; β-actin was amplified with forward: 5′ CCACTGGCATTGTCATGGACTC 3′, reverse: 5′ TTCCTTGATGTCACGGACGATTT 3′.
Quantitative real-time PCR were performed on a Bio-Rad Chromo 4 PCR System using SYBR® Premix Ex Taq ™ (Perfect Real Time) (TaKaRa Co. Ltd.). One microliter of cDNA was amplified in a 25-μL mixture containing 10 μM forward primer (0.5 μL), 10 μM reverse primer (0.5 μL), 2XSYBR® Premix Ex Taq™ (12.5 μL), and nuclease-free water (10.5 μL). The program comprised an initial denaturation step at 95°C for 5 min; followed by 40 cycles at 95°C for 15 s, 54°C for 30 s, and 72°C for 30 s; 72°C for 10 min; and a final melting curve. Three technical replicas were run. Delta CT (ΔCT) values were calculated to determine tissue-specific expression. The PCR results were analyzed by one-way analysis of variance to compare expression between tissues.
Western blot analysis of expression of mTOR in Inner Mongolia Cashmere goat fetal fibroblasts
Cashmere GFbs were used to seed 6-well tissue culture plates at 3×105 cells per well and incubated. Subconfluent cells were harvested with trypsin and washed with cold phosphate buffered saline, and cells were lysed in a buffer containing 20 mM Tris (pH 8.0), 137 mM NaCl, 100 g/L glycerol, 50 g/L Triton X-100, 2 g/L Na2VO4, and 4 g/L EDTA; 10 μL PMSF (0.1 M) and 10 μL ALT (10 g/L) were added per 1 mL lysis buffer immediately before use. The cell lysates were put on ice for 15 min and centrifugated at 15,000 rpm at 4°C for 20 min, and the supernatant was transferred into a new tube. The concentrations of the lysates were measured by the Bio-Rad protein determination method (Bio-Rad Laboratories).
Equal amounts (40 μg) of protein were electrophoresed in 12% or 8% (w/v) sodium dodecylsulfate polyacrylamide gels. Proteins were transferred to Hybond-polyvinylidene difluoride membranes (Amersham) and incubated with the primary antibodies overnight at 4°C and horseradish peroxidase-conjugated sheep (polyclonal) anti-mouse IgG (GE Healthcare UK Limited) at room temperature for 1 h. Signals were detected by chemiluminescence (Amersham).
MTT assay and IC50 calculation
Exponentially growing cells were seeded in 96-well plates at 6×103 cells per well 24 h before drug treatment. Then, cells were incubated with CCI-779 at various concentrations (0.037 μM, 0.11 μM, 0.33 μM, 1.0 μM, 3.0 μM, 9.0 μM, 18.0 μM, and 27.0 μM) for 48 h. 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (20 μL) (MTT, 5g/L; Sigma-Aldrich, Inc.) was added to each well in fresh medium and incubated for 4 h at 37°C. The formazan product was dissolved in 100 μL dimethyl sulfoxide per well and incubated for 10 min at 37°C. The MTT absorbance value was detected at 490/630 nm on a spectrophotometer set (Bio-TEK ELx800; Bio-tek instruments, Inc.). IC50 values were calculated using the Logit model.
Bioinformatics analysis of mTOR
Theoretical molecular weights of the deduced polypeptides and isoelectric points were predicted by calculating the isoelectric point (
Statistical analysis
Descriptive statistics were generated for all quantitative data, expressed as mean±SD. Each assay was performed in triplicate.
Results
Cloning and sequence analysis of Cashmere goat mTOR
The mTOR cDNA (GenBank accession HM114224) from Inner Mongolia cashmere goat comprises an open reading frame (ORF) of 7650 bp and a 952-bp residue of the 3′ UTR, which contains a polyadenylation signal at nucleotides 8575–8580 (AATAAA), 24 bp upstream of the polyA tract. The full cDNA sequence is 92% identical to homo sapiens mTOR and 92%, 92%, 91%, 89%, and 89% identical to that of pan troglodytes, equus caballus, canis lupus familiaris, rattus norvegicus, and mus musculus, respectively. The amino acid sequence of mTOR is 99% identical to that of homo sapiens and 98% to others.
Primary and secondary structure of Cashmere goat mTOR
The deduced primary structure of Inner Mongolia Cashmere goat mTOR consists of 2549 amino acids. There are 8 N-glycosylation sites, 5 cAMP- and cGMP-dependent protein kinase phosphorylation sites, 21 protein kinase C phosphorylation sites, 3 tyrosine kinase phosphorylation sites, 3 endoplasmic reticulum targeting sequences (Fig. 1A), an FAT domain (residues 1382–1982), an FKBP12 rapamycin-binding domain (FRB, residues 2019–2112), a protein kinase-like domain (residues 2086–2480), and an FATC domain (residues 2517–2549) (Fig. 1B).

The predicted amino-acid sequence and domains of mTOR
The predicted molecular weight of unmodified mTOR is M r 288.6 KDa, and the estimated isoelectric point (pI) is 7.24. With regard to basic residues, mTOR contains 5.1% lysine, 6% arginine, 3.26% histidine, and has 5.02% aspartate and 6.51% glutamate for acidic residies. mTOR was predicted to be an integral membrane protein using PSORT (
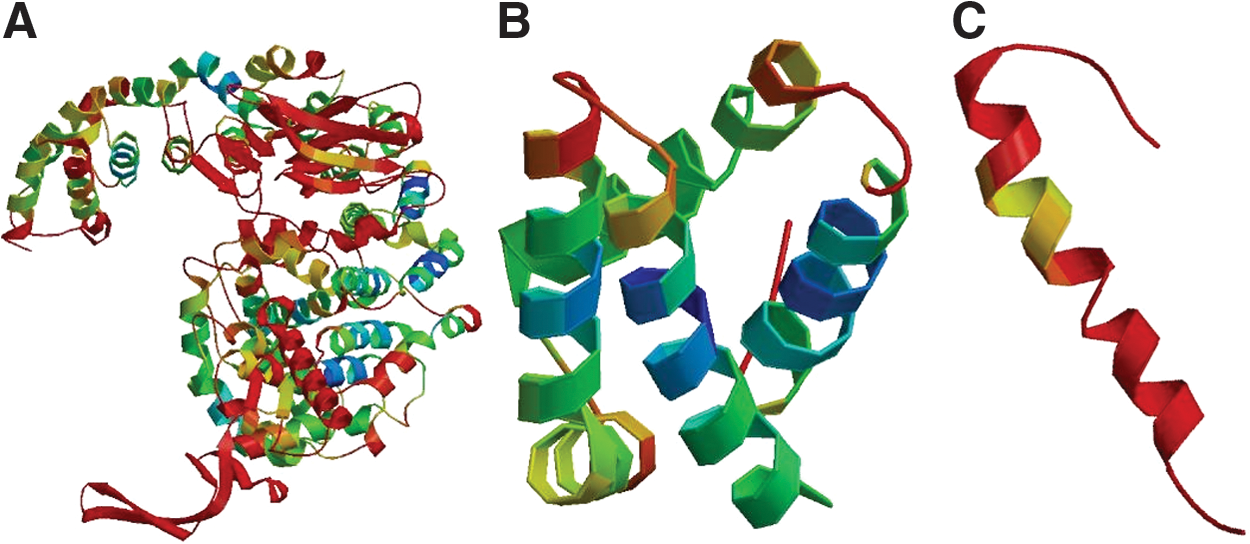
Three-dimensional model of mTOR regions.
Tissue distribution of the Cashmere goat mTOR mRNA
By quantitative real-time PCR, mTOR mRNA was found to be more abundant in brain, testis, kidney, and spleen compared with liver, heart, and lung tissue from Cashmere goat (Fig. 3A).
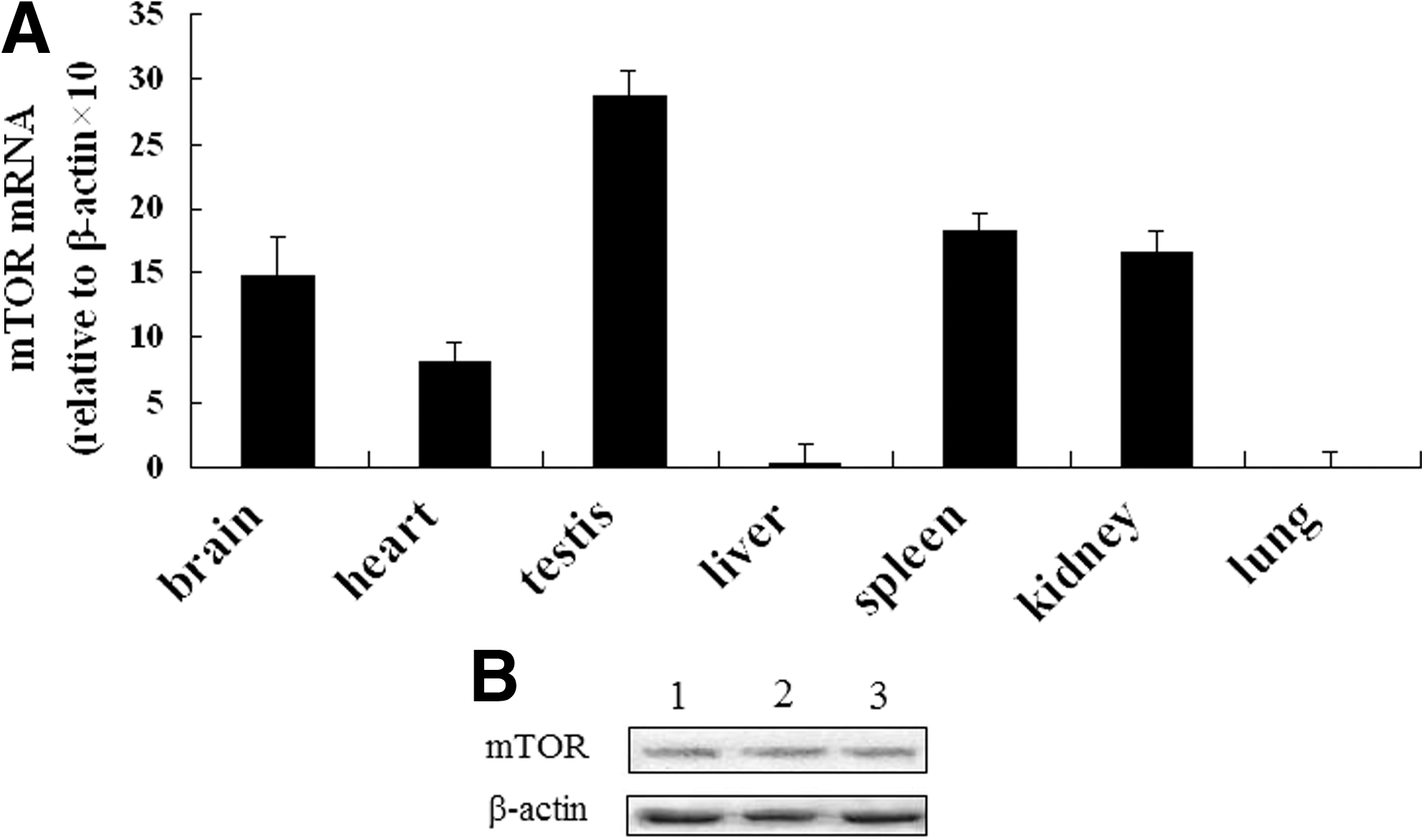
Quantitative real-time PCR analysis of mTOR mRNA expression in various tissues and Western blot analysis of mTOR expression in Cashmere goat fetal fibroblasts.
Expression of mTOR in Cashmere goat fetal fibroblasts
Total protein from Cashmere GFbs was extracted and assayed by western blot. mTOR was expressed in GFbs (Fig. 3B).
CCI-779 inhibits Cashmere goat fetal fibroblast proliferation
We measured the IC50 value of CCI-779 to examine its cytostatic effects on GFbs and optimize its concentration for further experiments. GFbs were treated with various concentrations of CCI-779 (0.037 μM–27 μM) for 48 h, and their susceptibility was measured by MTT assay. The IC50 of CCI-779 against GFbs was 10.88 μM (Fig. 4), and their proliferation was inhibited.
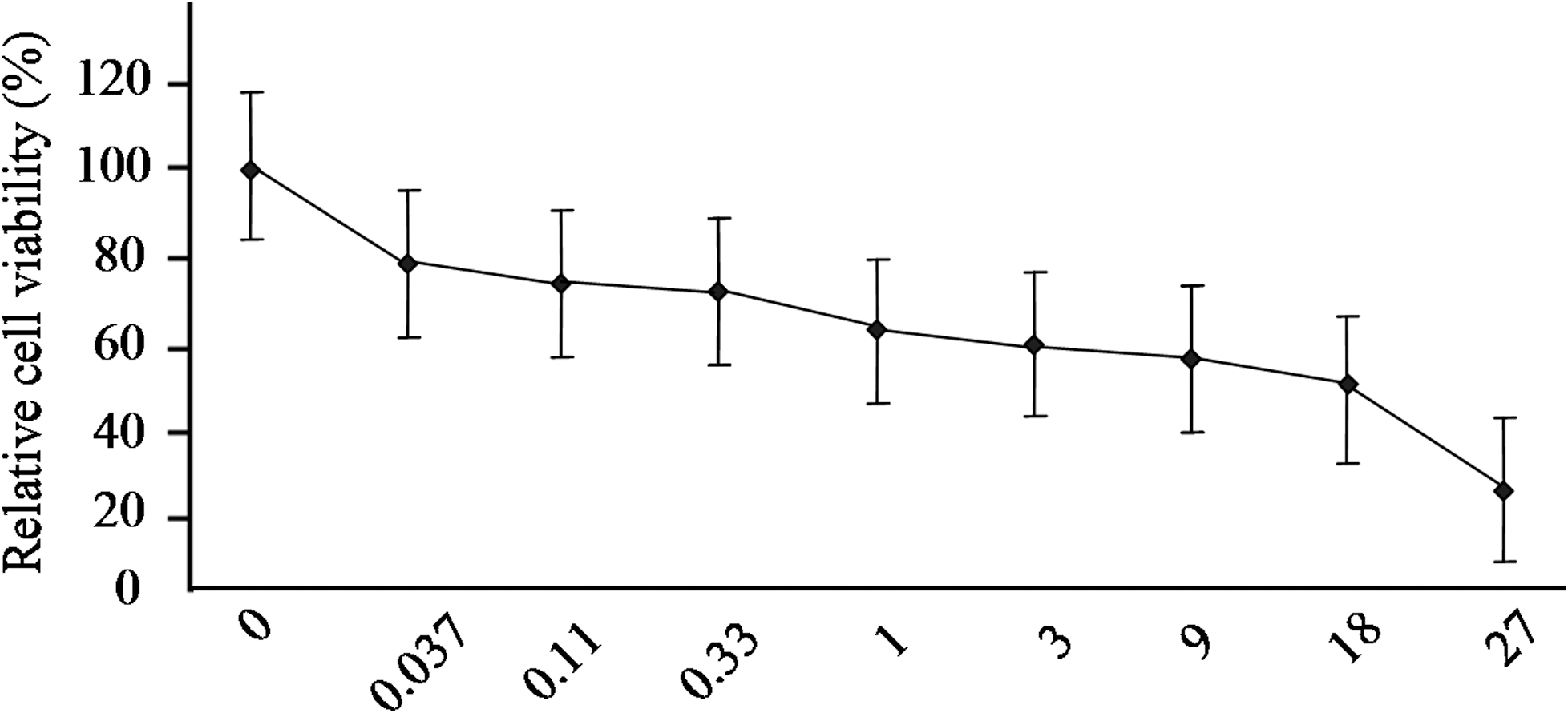
Inhibition curve of CCI-779 on GFbs growth. GFbs were treated with various concentrations of CCI-779 (0.037 μM to 27 μM) for 48 h and subjected to MTT assay.
Discussion
By understanding how signaling molecules regulate mTOR function, we can gain insights into mechanisms of cell growth, proliferation, and apoptosis. mTOR governs important functions in peripheral organs (Catania et al., 2010)—its activation is sufficient to stimulate specific metabolic pathways, such as glycolysis, de novo lipid biosynthesis (Duvel et al., 2010), cholesterol trafficking, and fasting-induced ketogenesis (Sengupta et al., 2010; Xu et al., 2010).
The aim of our study was to describe mTOR in Inner Mongolia Cashmere goat. The ORF of goat mTOR cDNA encodes the same number of amino acids as those of rattus norvegicus, mus musculus, and homo sapiens (2549), and its sequence is 99% identitcal to that of the human ortholog and 98% to others.
mTOR is an evolutionarily conserved, nutrient-sensing protein kinase that regulates growth and metabolism in all eukaryotic cells (Wullschleger et al., 2006). mTOR was predicted to be an integral membrane protein, and mTOR lies on the surface of lysosomes (Sancak et al., 2010). There are three predicted endoplasmic reticulum-targeting sequences in mTOR, but it is unknown whether these sequences are related to the location of mTOR or which one takes effect. Nevertheless, mTOR is active in the cytoplasm and nucleus (Kantidakis et al., 2010; Wei and Zheng, 2009), regulating transcription. However, how mTOR executes its various functions is unknown.
The 3′ UTR of goat mTOR is much longer than those of five other cDNAs with intact 3′ UTRs of goat genes in GenBank: S6, GPx4, K26, LDH-A, and S6K1. In transiently transfected Chinese hamster ovary cells with a firefly luciferase reporter, the length of the 3′ UTR regulates the translational efficiency and stability of mRNA (Tanguay and Gallie, 1996). On average, genes that have more miRNA sites have longer 3′ UTRs; proteins with 3′ UTRs that have more target sites are predominantly active during development, whereas with fewer target sites in the 3′ UTR execute basic functions (Stark et al., 2005), suggesting that mTOR is an important regulator of cellular growth.
As an evolutionarily conserved protein kinase that belongs to the PIKK family, mTOR has a C-terminal protein kinase-like domain that is common to other members. Immediately upstream of the catalytic domain lies the FRB domain, to which FKBP12-rapamycin binds to mTOR to inhibit its activity. In addition, FAT is another ubiquitous domain of the PIKK family, located upstream of the FRB domain. The C-terminus contains a FATC domain that regulates the activity of mTOR (Peterson et al., 2000; Takahashi et al., 2000). In this study, we described the basic data about goat mTOR, and the predicted domains of goat mTOR were as same as those in former research.
S6Ks are well-characterized downstream molecules that are regulated by mTOR (Chung et al., 1992; Hay and Sonenberg, 2004). S6K is suppressed by the mTOR inhibitor RAD001 in SEG-1 esophageal cancer cells, (Wang et al., 2010), and S6K regulates the lifespan (Blagosklonny and Hall, 2009). Akt is an upstream regulator of mTOR, and S6Ks negatively regulate insulin signaling through insulin receptor substrate 1, impairing PI3-kinase/Akt signaling downstream of the insulin receptor (Shah and Hunter, 2006).
The mTOR signaling pathway has been extensively investigated in humans and some model animals, and mTOR has recently been demonstrated to regulate protein synthesis in pigs and bovines (Burgos et al., 2010; Murgas Torrazza et al., 2010; Suryawan et al., 2010; Toerien et al., 2010). Lack of the fundamental information about genes and their expression as well as regulation has become a bottleneck for the molecular studies in these important livestock. Our study provides an original work about mTOR of Cashmere goat, and the results may be useful to the researchers of goat and sheep.
In this study, we examined the function of mTOR in GFbs, demonstrating that it is important for their growth. We infer that mTOR regulates the proliferation of GFbs in addition to human esophageal cancer cells SEG-1, but further study is needed to generate more data on how the mTOR signaling pathway is regulated in Inner Mongolia Cashmere goat cells.
Conclusions
We cloned mTOR from Inner Mongolia Cashmere goat fetal fibroblasts, which includes an ORF of 7650 bp and a 3′ UTR of 909 residues. mTOR is expressed at higher levels in testis, spleen, kidney, and brain compared with heart, liver, and lung. The treatment of CCI-779 decreases the viability of GFbs, and the IC50 of CCI-779 against GFbs was 10.88 μM. mTOR plays a key role in growth regulation of Inner Mongolia Cashmere goat fetal fibroblasts.
Footnotes
Acknowledgments
This work was supported by a grant from the Natural Sciences Foundation of China (No. 31160469), Natural Sciences Foundation of Inner Mongolia (No. 2011MS0521), and graduate student research projects of Inner Mongolia University.
Disclosure Statement
No competing financial interests exist.