
Abstract
The epidermal growth factor (EGF) pathway stimulates proliferation and differentiation of epidermal and epithelial tissues, and plays an important role in tumorigenesis. The association between EGF polymorphisms and cancer risk is controversial; thus, we performed this meta-analysis. Overall, 41 case–control studies with 9,779 cases and 15,932 controls were retrieved. We found that EGF +61A/G polymorphism increased overall cancer risk (G allele vs. A allele: OR=1.181, 95% CI=1.077–1.295, P heterogeneity < 0.001; GG vs. AA: OR=1.370, 95% CI=1.143–1.641, P heterogeneity < 0.001; GG+GA vs. AA: OR=1.175, 95% CI=1.047–1.318, P heterogeneity < 0.001). In the stratified analysis by cancer type, the +61 G allele was a risk factor for colorectal cancer, esophageal carcinoma, gastric cancer, and hepatocellular carcinoma. Individuals who carried +61G allele had higher cancer susceptibility in mixed and European racial subgroups. An increased association was detected in the hospital-based subgroup. No significant association was found among EGF −1380A/G, −1744G/A, rs6983267T/G polymorphisms and cancer risk.
Introduction
Shahbazi et al. (2002) first reported a functional SNP involving an A-to-G mutation at position 61 of the 5′-untranslated region of the EGF gene (rs4444903) in 2002. They demonstrated that the G allele increased EGF protein expression by affecting DNA folding or mRNA transcript in vitro and revealed that patients with malignant melanoma of skin had a significantly higher frequency of G allele compared with the general population. Jin et al. (2007) first found two SNPs (−1380A/G and −1744G/A) in the promoter region of EGF. They reported no significant association between −1380A/G polymorphism and gastric cancer; however, the −1744 G allele played a protective role. Moreover, Daraei et al. (2011) reported that rs6983267 G allele in the 8q24 region was a risk factor in the development of colorectal cancer.
To evaluate the effects of these four EGF promoter SNPs and cancer susceptibility, we analyzedt genotypes among SNPs of +61A/G, −1380A/G, −1744G/A, and rs6983267 in 9,779 cases and 15,932 controls.
Materials and Methods
Identification and eligibility of relevant studies
We searched the PubMed database, last search updated on August 1, 2011, with the keywords containing “EGF” or “epidermal growth factor,” “polymorphism” or “variant,” and “cancer” or “carcinoma,” without any restriction on language or publication year. Using these terms, a total of 122 articles were retrieved, of which 41 articles met the inclusion criteria. We also screened reference of the retrieved articles and review articles by hand.
Inclusion criteria and exclusion criteria
Studies that were included in our analysis met all of the following criteria: (a) the study assessed the correlation between cancer risk and EGF polymorphisms, (b) case–control studies, and (c) sufficient genotype numbers for cases and controls. Accordingly, the following exclusion criteria were also used: (a) no control population, (b) no available genotype frequency, and (c) duplication of the previous publications.
Data extraction
Two of the authors extracted all data independently. The following items were collected: first author's last name, year of publication, country of origin, ethnicity, cancer type, total case/control number, source of control, Hardy–Weinberg equilibrium (HWE) of controls, and genotyping methods.
Statistical analysis
Crude odds ratios (ORs) with 95% confidence intervals (CIs) were used to measure the strength of the association between EGF polymorphisms and cancer risk based on the genotype frequencies in cases and controls. Subgroup analysis stratified by cancer type was performed first. If one cancer type was reported in only one individual study, it is classified to the “others” subgroup. Ethnicity was categorized as European, Asian, and mixed. Source of control subgroup analysis was performed on two classifications: population based (PB) and hospital based (HB).
The fixed effects model and the random effects model were used to calculate the pooled OR value. The statistical significance of the summary OR was determined with the Z-test. Heterogeneity assumption was evaluated with a chi-square–based Q-test among the studies. A p value of more than 0.05 for the Q-test indicated a lack of heterogeneity among the studies. In case significant heterogeneity was detected, the random effects model (DerSimonian-Laird method) was used; however, the fixed effects model (Mantel-Haenszel method) was chosen (Mantel and Haenszel, 1959; DerSimonian and Laird, 1986). We investigated the relationship between EGF genetic variants and cancer risk in allelic contrast (G allele vs. A allele for +61A/G and −1380A/G; A allele vs. G allele for −1744G/A; G-allele vs. T-allele for rs6983267), homozygote comparison (GG vs. AA for +61A/G and −1380A/G; AA vs. GG for −1744G/A; GG vs. TT for rs6983267), and dominant genetic model (GG+GA vs. AA for +61A/G and −1380A/G; AA+AG vs. GG for −1744G/A; GG+GT vs. TT for rs6983267). Sensitivity analysis was performed by omitting each study in turn to assess the stability of the results. The departure of frequencies of EGF polymorphisms from expectation under HWE was assessed by χ 2 test in controls using the Pearson chi-square test; p<0.05 was considered significant. All statistical tests for this meta-analysis were performed with Stata software (version 10.0; StataCorp LP, College Station, TX).
Genotyping methods
Genotyping for four SNPs in EGF gene within the studies analyzed was conducted using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP), primer introduced restriction analysis-polymerase chain reaction (PIRA-PCR), matrix-assisted laser desorption ionization-time of flight (MALDI-TOF), polymerase chain reaction–single strand conformation polymorphism (PCR-SSCP), Taqman and ARMS test.
Results
Study selection and characteristics in the meta-analysis
A total of 122 articles was identified: 13 review articles were excluded, 1 duplicated article was also excluded (if two studies with overlapping data were published by the same investigators, only the most recent or complete study was included in our meta-analysis.) (Qi et al., 2008, 2009). Overall, 41 case–control studies with 9,779 cases and 15,932 controls were retrieved based on the search criteria for whole cancer susceptibility related to the four EGF polymorphisms. The controls were mainly healthy populations. If one study contained two or more races as its research subjects, we consider it mixed. In our study, there were 18 European case–control studies, 19 Asian case–control studies, and 4 mixed case–control studies for +61A/G; two Asian studies for −1380A/G and −1744G/A; and one Asian study for rs6983267. The controls in 26 studies were HB and 15 were PB for +61A/G. For the +61A/G polymorphism, the G allele frequency of Asians was significantly higher than mixed individuals in controls (p<0.001); moreover, the mixed population that carried the G allele was significantly higher than Europeans (p=0.001) (Fig. 1). A similar trend was found in cases. The distribution of genotypes in all the controls was in agreement with HWE, except for four studies (Shahbazi et al., 2002; Vauleon et al., 2007; Abu Dayyeh et al., 2011; Araújo et al., 2011b).
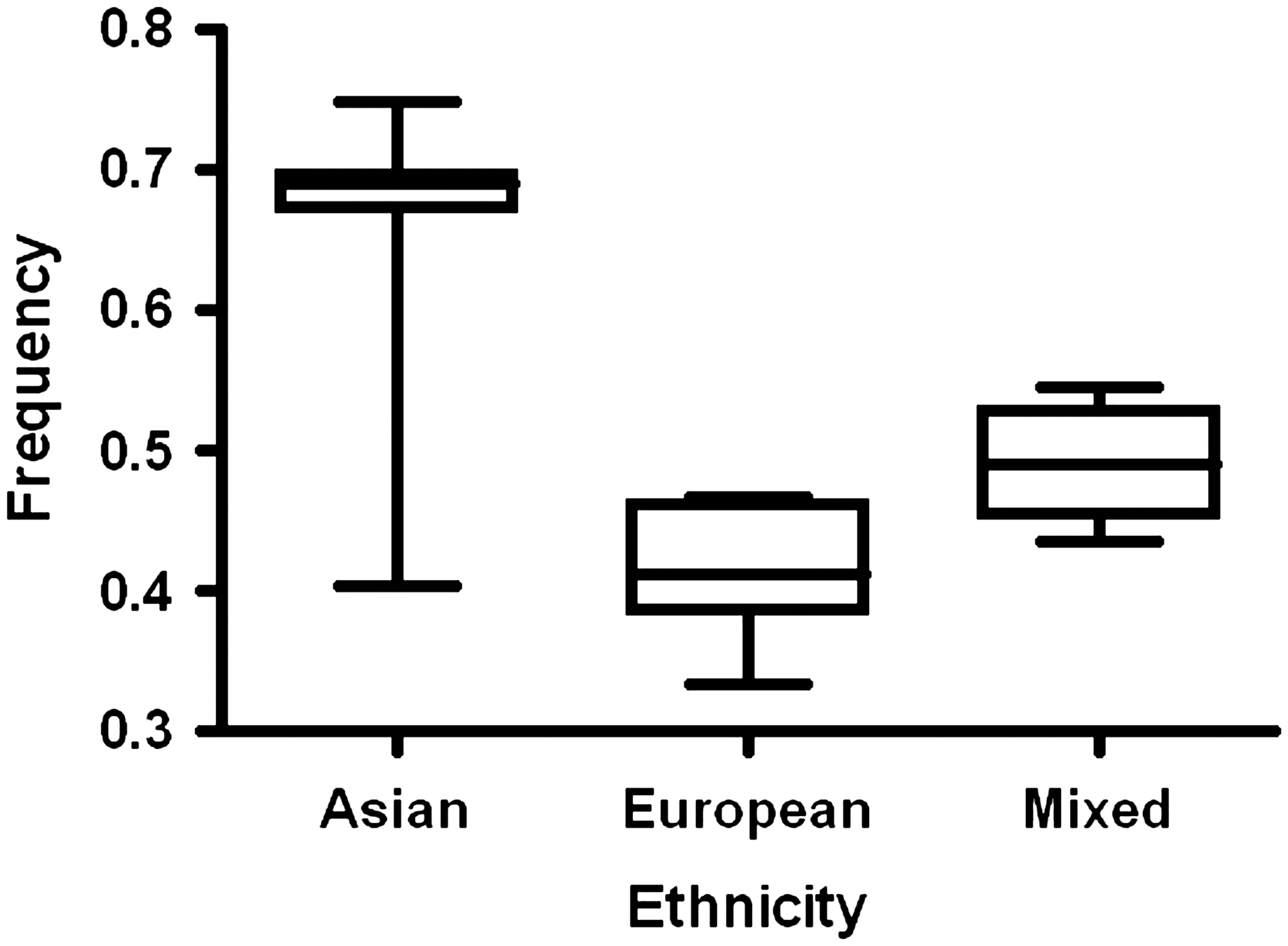
Frequencies of the epidermal growth factor +61G allele among control subjects stratified by ethnicity.
Meta-analysis
Table 1 shows the summary OR of EGF +61A/G on the basis of 9,779 cancer cases and 15,932 controls. Overall, we observed an increased association between the EGF +61A/G polymorphism and cancer risk in all three genetic models (G allele vs. A allele: OR=1.181, 95% CI=1.077–1.295, P
heterogeneity<0.001; GG vs. AA: OR=1.370, 95% CI=1.143–1.641, P
heterogeneity<0.001; GG+GA vs. AA: OR=1.175, 95% CI=1.047–1.318, P
heterogeneity<0.001). In the subgroup by cancer type, significantly increased relationships were found in four kinds of cancers (colorectal cancer: G allele vs. A allele, OR=1.144, 95% CI=1.060–1.235, P
heterogeneity=0.100; GG vs. AA, OR=1.293, 95% CI=1.138–1.469, P
heterogeneity<0.001; GG+GA vs. AA, OR=1.064, 95% CI=1.000–1.132, P
heterogeneity=0.119; esophageal carcinoma: G allele vs. A allele, OR=1.168, 95% CI=1.092–1.249, P
heterogeneity=0.248; GG vs. AA, OR=1.298, 95% CI=1.150–1.465, P
heterogeneity<0.001; GG+GA vs. AA, OR=1.087, 95% CI=1.024–1.155, P
heterogeneity=0.108; gastric cancer: G allele vs. A allele, OR=1.048, 95% CI=1.010–1.087, P
heterogeneity=0.191; hepatocellular carcinoma: G allele vs. A allele, OR=1.105, 95% CI=1.057–1.156, P
heterogeneity=0.007; GG vs. AA, OR=1.172, 95% CI=1.105–1.242, P
heterogeneity=0.002; GG+GA vs. AA, OR=1.073, 95% CI=1.039–1.108, P
heterogeneity=0.111). Given the ethnicity differences in the allele frequency of this sequence variant, we evaluated the effect of EGF +61A/G polymorphism in Asian, European, and mixed populations, respectively. Association of +61A/G polymorphism with increased cancer risk in European population was observed under allelic contrast (OR=1.155, 95% CI=1.018–1.310, P
heterogeneity<0.001) and homozygote comparison (OR=1.322, 95% CI=1.028–1.700, P
heterogeneity<0.001). Mixed individuals that carried the G allele had an increased cancer risk (G allele vs. A allele, OR=2.015, 95% CI=1.095–3.709, P
heterogeneity<0.001; GG vs. AA, OR=3.258, 95% CI=1.220–8.698, P
heterogeneity=0.001; GG+GA vs. AA, OR=2.230, 95% CI=1.058–4.701, P
heterogeneity=0.006). Summary OR for +61A/G polymorphism stratified by source of control was evaluated, and we also observed an increased association in the allelic contrast (G allele vs. A allele, OR=1.252, 95% CI=1.080–1.451, P
heterogeneity<0.001) and in the other two genetic models with HB controls. No significant association was detected between
Ph, p-value of Q-test for heterogeneity; in case significant heterogeneity was detected, the random effects model was used; however, the fixed effects model was chosen.
PB, population-based of control group; HB, hospital-based of control group; OR, odds ratio; CI, confidence interval.
Sensitivity analysis
We used sensitivity analysis to evaluate whether modification of the inclusion criteria of the meta-analysis affected the results. We examined the influence of each study on the pooled OR by repeating the analysis while omitting each study one at a time. Moreover, the estimated pooled OR did not change when excluding the studies that were not in HWE.
Discussion
The EGF protein, identified as a potent mitogenic peptide, has multiple biological roles, including proliferation, differentiation, and tumorigenesis of epithelial tissues via interaction with its receptor EGFR (Fisher and Lakshmanan, 1990; Normanno et al., 2001; Harari et al., 2007).
We performed a meta-analysis of association among EGF SNPs (+61, −1380, −1744, and rs6983267) and risk for different types of cancer based on 41 case–control studies. There was significant association between EGF SNP +61A/G and risk of most cancers. Variant genotypes of +61 were associated with a significantly increased cancer risk among both mixed and European populations. Finally, individuals who carried the +61 G allele may have a higher frequency of cancer risk. No association was detected among −1380, −1744, and rs6983267 SNPs and cancer risk. These findings indicate that EGF +61A/G polymorphism may play a critical role in cancer development.
Many studies provided evidence that variant genotypes of EGF were associated with a significantly increased risk for cancer; for example, Teixeira et al. (2008) indicated that carriers of the G-allele had a significantly higher age-adjusted risk for being diagnosed with metastatic prostate cancer and with higher Gleason grade tumors; Hamai et al. (2005) suggested that the A/G polymorphism of EGF was involved not only in the occurrence but also in the malignant progression of gastric cancer. However, it was shown that the GG genotype was not associated with colorectal cancer, gastric cancer, lung cancer, nasopharyngeal carcinoma (Goto et al., 2005; Kang et al., 2007a; Gao et al., 2008; Daraei et al., 2011). Moreover, several studies also found that the variant genotypes of EGF were associated with decreased cancer risk (Araújo et al., 2009b; Bao et al., 2010). To clarify the association between the polymorphisms and cancer risk, we performed a meta-analysis. The results showed that the variant genotypes of +61 were associated with a significantly increased cancer risk for all cancer types. As for cancer type, we found that the variant G allele of EGF +61 was associated with a significantly increased risk for most cancers (colorectal cancer, esophageal carcinoma, gastric cancer, and hepatocellular carcinoma). And we also demonstrated that this SNP does not have an impact on the risk of breast cancer, cervical cancer, glioma, lung cancer, and melanoma. It is not uncommon for the same polymorphism to play different roles in cancer susceptibility across different tumor locations because cancer is an extremely complex disease and genetic heterogeneity exists in different cancer types (Hirschhorn et al., 2002). One important property of the gene polymorphisms is that their incidence can vary substantially among different racial or ethnic populations. In the subgroup analysis by ethnicity, we found that the association among the EGF +61 polymorphism and increased risks of cancer was significant in mixed individuals and Europeans, but not in Asians. Two main reasons can explain this difference: (a) differences in genetic and environmental background exist among different ethnicities; (b) different populations usually have different linkage disequilibrium patterns.
We divided control populations into two parts: HB and PB. In our study, we detected that there was increased association between EGF +61 SNP and cancer risk in HB subgroup. Daraei et al. (2011) found that GG genotype of rs6983267 was a risk factor for the development of sporadic colorectal cancer; in addition, Jin et al. (2007) showed that −1380A/G and −1744G/A may alter plasma EGF levels together with +61A/G, resulting in disturbance of the EGF signaling. However, no relationships were detected in our study. The small sample size could explain this reason.
Some limitations in our meta-analysis should be mentioned. First of all, the number of published studies included in our meta-analysis was not sufficiently large for a comprehensive analysis, especially for −1380A/G, −1744G/A, and rs6983267T/G polymorphisms. Second, the interactions between gene–gene, gene–environment, and even different polymorphic loci of the same gene may modulate cancer risk, so future research and analysis should include these factors. Third, our meta-analysis was based on unadjusted estimates. A more precise analysis could be conducted if individual data were available, which would allow for the adjustment by other covariates including age, sex, family history, environmental factors, cancer stage, and lifestyle. Fourth, the controls were not strictly healthy controls. In summary, our meta-analysis showed evidence that EGF +61A/G polymorphism was associated with significantly increased risk for cancer. Therefore, further well-designed large studies focused on gene–gene and gene–environment interactions are warranted.
Footnotes
Disclosure Statement
No competing financial interests exist.