
Abstract
The generation of oocyte-like cells (OLCs) from stem cell differentiation in vitro provides an optimal approach for studying the mechanism of oocyte development and maturation. The aim of this study was to investigate the activation of bone morphogenetic protein 15 gene (BMP15) during the differentiation of human amniotic fluid stem cells (hAFSCs) into OLCs. After 15 days of differentiation, OLCs with a diameter of 50–60 μm and zona pellucida (ZP)-like morphology were observed. Reverse transcription-polymerase chain reaction (RT-PCR) analysis showed the BMP15 was activated from approximately day 10 of differentiating hAFSCs and thereafter. The reporter construct pBMP15-enhanced green fluorescent protein (EGFP) was transiently transfected into the differentiated hAFSCs and the EGFP expression driven by the BMP15 promoter was positive in the OLCs. Moreover, RT-PCR analysis showed that the oocyte-specific markers including ZP1, ZP2, ZP3, and c-kit were expressed in the differentiated hAFSCs, and the immunofluorescence assay confirmed that the ZP2 was detected in the OLCs. Quantitative RT-PCR revealed that ZP2 and ZP3 were significantly elevated in the differentiated hAFSCs. Further, in the OLCs derived from hAFSCs, the BMP15 promoter directing the EGFP reporter was colocalized with ZP2. Together, these results illustrated that the BMP15 could be used as an oogenesis marker to track hAFSCs differentiation into the OLCs.
Introduction
To track and identify specific stages of female gamete induction, reporter constructs have been generated by linking the promoter of germ cell markers with enhanced green fluorescent protein (EGFP), such as Oct4-EGFP (Hubner et al., 2003), GDF9-EGFP (Salvador et al., 2008), and Dazl-EGFP (Linher et al., 2009). However, no report has so far used the bone morphogenetic protein 15 (BMP15) gene as the marker. BMP15 is an oocyte-specific gene that shares high homology with GDF9 (Wu and Matzuk, 2002), and plays crucial roles in both oocyte maturation and fertilization (Pennetier et al., 2004). Further, BMP15 is essential for the granulosa cell development and oocyte function (Su et al., 2004). Therefore, BMP15 is a potential marker to monitor the oogenesis development.
In this study, we demonstrated that hAFSCs, containing extensive self-renewal ability and remaining highly viable in long-term cultures, differentiated into putative female gametes when hAFSCs were induced in the medium supplemented with porcine follicular fluid. The EGFP driven by the promoter of BMP15 gene was observed in the OLCs that were differentiated from hAFSCs, indicating that the BMP15 reporter vector could be used as a marker of studying oocyte differentiation.
Materials and Methods
Cell culture and differentiation
The protocols of handling human and animal specimens in this study were approved by the Ethical Review Board in the hospital of Xi'an Jiaotong University and the Ethics Committee of Northwest A&F University (Xi'an, China). hAFSCs were isolated from human amniotic fluid of the second trimester of gestation as previously described (Wang et al., 2008). Briefly, human amniotic fluid samples were centrifuged at 500 g for 8 min, and the pellets were washed three times with phosphate-buffered saline (PBS). The cell pellets were resuspended and plated onto a six-well culture plate (Costar) containing the alpha modified minimum essential medium (α-MEM; Invitrogen) supplemented with 10% fetal bovine serum (FBS; Gibco), 1% glutamine, and antibiotics (100 U/mL penicillin, 100 μg/mL streptomycin). When the cells came to 80% confluence, the cells were harvested by trypsinization and subcultured at 37°C in 5% CO2 incubator.
To perform the differentiation, hAFSCs in the passage 10 were dissociated and plated onto a cell plate with the induction medium which consisted of α-MEM supplemented with 5% FBS, 0.1 mM nonessential amino acids, 2 mM
Plasmid pBMP15-EGFP construction and transfection
The 1137 base pair fragment of human BMP15 promoter region (−1136 to +1 with respect to the start codon) was amplified from genomic DNA of hAFSCs by (PCR) using the primers listed in Table 1. The PCR product was cloned into pMD18-T vector and confirmed by sequencing. The BMP15 fragment was subcloned into the promoterless GFP reporter vector pEGFP1. Positive clones were confirmed by restriction enzyme digestion and named pBMP15-EGFP. The construct was transfected into the induced and uninduced hAFSCs using the Lipofectamin 2000 (Invitrogen) according to the manufacturer's protocol. Six hours after the transfection, the medium was replaced with fresh culture medium. Hoechst 33342 staining was employed to detect the nuclear DNA at 24 h after the transfection. The images were obtained by an Olympus BX-UCB microscope.
Primers were used for real-time reverse transcription-polymerase chain reaction.
Reverse transcription-PCR and quantitative reverse transcription-PCR
Total RNA was isolated from cells at different days of differentiation with Trizol (Invitrogen) according to the manufacturer's protocol. One microgram of total RNA was reverse transcribed into cDNA using the oligo-dT primers and RevertAid™ reverse transcriptase (Fermentas). The PCR programs were as follows: the initial denaturation at 95°C for 5 min, 35 cycles of denaturation at 94°C for 30 s, annealing at 57°C for 30 s, and extension at 72°C for 60 s, and a final extension step at 72°C for 10 min. The expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal control. Sequences of primers were listed in Table 1. Relative mRNA levels were evaluated by real-time reverse transcription-PCR that was carried out on a programmed thermal cycler (LineGene 9600; BIOER) by using the SYBR Premix Ex Taq™ II kit (TaKaRa). Generally, the reaction mixture consisted of 2 μL of cDNA, 0.8 μL of forward and reverse primers (10 μM), 10 μL of 2×SYBR Premix Ex Taq, and RNase-free H2O to make a final volume of 20 μL. The PCR programs contained the initial denaturation at 95°C for 3 min and 40 cycles at 94°C for 15 s and 60°C for 25 s. Relative mRNA levels were determined using the 2−ΔΔCt method and normalized to GAPDH.
Western blotting
To examine the expression level of VASA, both induced and uninduced hAFSCs were collected and treated with RIPA lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM ethylenediaminetetraacetic acid, 1% Triton X-100, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS]). Protein was collected by centrifugation (10 min, 12,000 g, 4°C) and then stored in −80°C. The same amount of protein sample was added into each well and separated on 10% SDS-polyacrylamide gel electrophoresis at 80 V for 2 h. After the electrophoresis, proteins were transferred onto the nitrocellulose membrane that was then blocked by 1% bovine serum albumin (BSA) for 2 h. The membranes were incubated with either polyclonal rabbit anti-human DDX4 (also called Vasa in human) antibody (ab13840, Abcam; 1:1000) or anti-beta actin antibody (ab8229, Abcam; 1:1000) for 2 h, and followed by incubation with HRP-conjugated secondary antibody. Immunoreaction was detected using the ECL kit (Pierce).
Immunofluorescence
To conduct the immunofluorescence, cells were washed twice with PBS and fixed in 4% paraformaldehyde in PBS for 20 min. Fixed cells were washed twice, incubated in PBS containing 0.2% Triton X-100 for 10 min, and then washed three times. Cells were blocked with BSA-blotting buffer (1% BSA, 0.1% Tween 20 in PBS) for 30 min, and then incubated with BSA-blotting buffer plus anti-ZP2 (SC-30222, Santa Cruz; 1:500) antibody overnight at 4°C. After washing three times with PBS, cells were incubated with TRITC conjugated anti-rabbit IgG (Santa Cruz; 1:200) for 45 min at room temperature, and visualized by the fluorescence microscope.
Results
Cell morphology changes during hAFSC differentiation
hAFSCs derived from human amniotic fluid, with the fibroblastic morphology (Fig. 1B1), have been propagated in vitro for more than 45 passages (Wang et al., 2008). The hAFSC cell line hAF-07, which maintained the normal female karyotype (46, xx) and showed the optimal growth potential (Wang et al., 2008), was selected for the induction experiments (Fig. 1A). When hAFSCs were induced in the medium supplemented with 5% porcine follicular fluid for 5 days, a subpopulation of round cells were observed in culture plates (Fig. 1B2). After differentiating hAFSCs for 10 days, cell aggregates were formed and the large extruded cells were observed with a diameter of 30–40 μm (Supplementary Fig. S1A; Supplementary Data are available online at
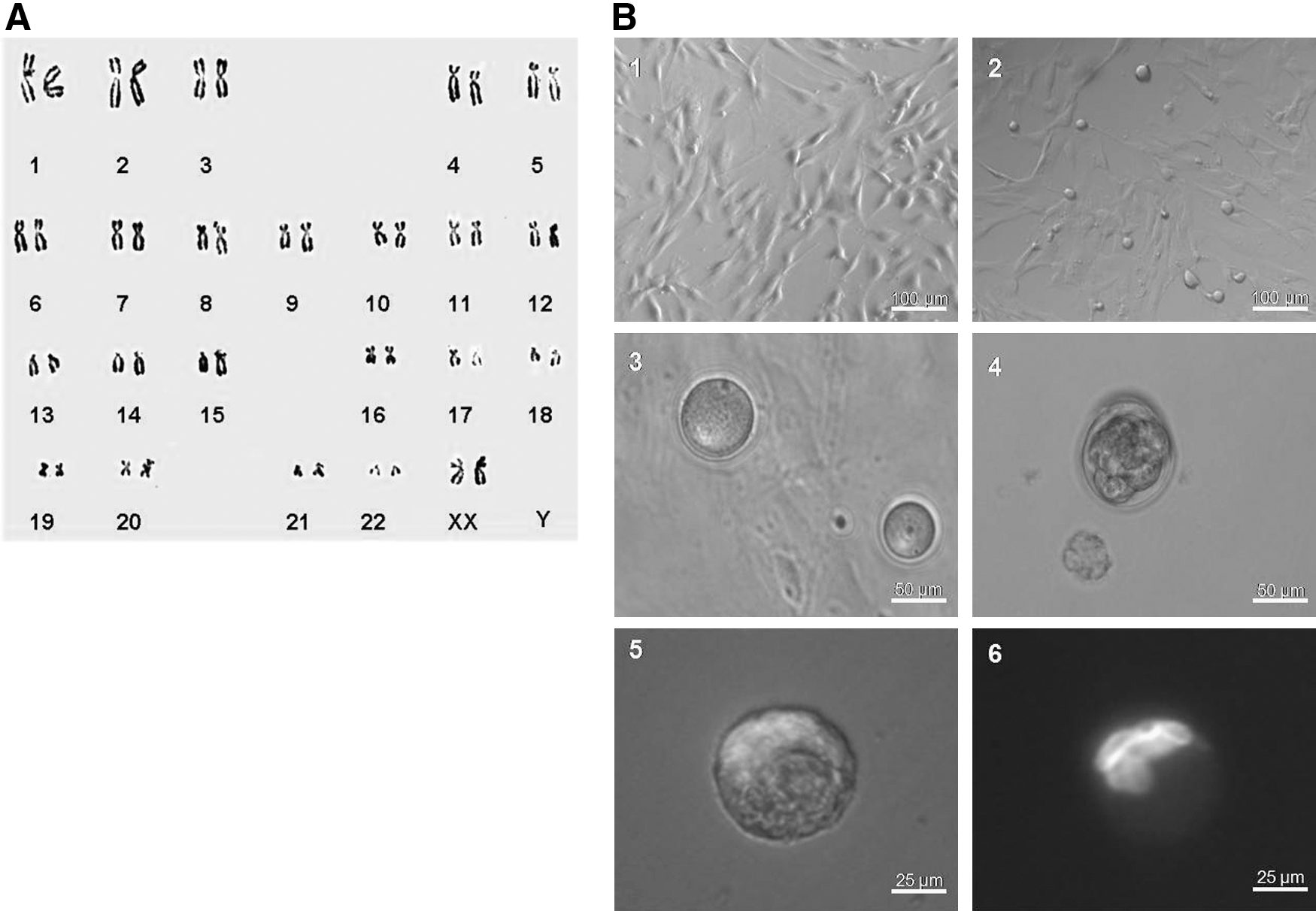
Human amniotic fluid stem cells (hAFSCs) differentiated into oocyte-like cells (OLCs).
Construction of pBMP15-EGFP vector
Originally we planned to use GDF9, a member of transforming growth factor beta superfamily and an oocyte specific marker previously reported (Salvador et al., 2008), as the reporter to monitor hAFSCs differentiation. However, the semi-quantitative RT-PCR result showed that GDF9 gene was constitutively expressed in hAFSCs, irrespective of the differentiation status (Fig. 2A). On the other hand, BMP15 that is highly homologous with GDF9 (Wu and Matzuk, 2002) and an oocyte-specific gene was activated from day 10 (D10) of differentiation and thereafter (Fig. 2B), suggesting that BMP15 could be a potential reporter for screening OLCs. Therefore we constructed the BMP15 reporter plasmid pBMP15-EGFP that contained the 1137 base pair BMP15 promoter region that was ligated into the promoterless EGFP reporter vector pEGFP1. The schematic representation of pBMP15-EGFP was shown in Figure 2C.
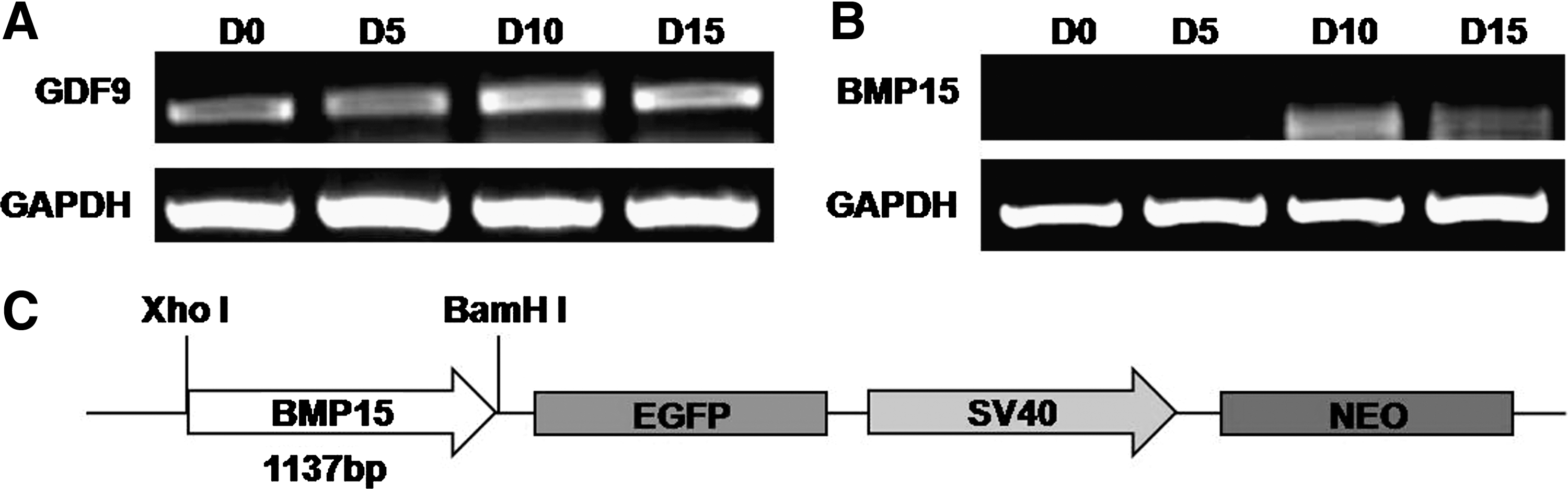
Construction of pBMP15-enhanced green fluorescent protein (EGFP) vector.
Activation of BMP15 gene in OLCs
The pBMP15-EGFP plasmid was transiently transfected into the induced and uninduced hAFSCs. The EGFP fluorescence signal was observed in the differentiated round cells at D10, but was absent in uninduced hAFSCs cells (Fig. 3A), indicating that BMP15 gene was activated during hAFSCs differentiation. By extending the duration of induction (D15), the OLCs with a diameter of 50–60 μm were formed, and the EGFP expression driven by BMP15 promoter was positive in OLCs (Fig. 3B1, B2). This observation plus the previous RT-PCR result (Fig. 2B) further confirmed that BMP15 gene was activated in OLCs and could be used as the marker to monitor the oocyte development in vitro.
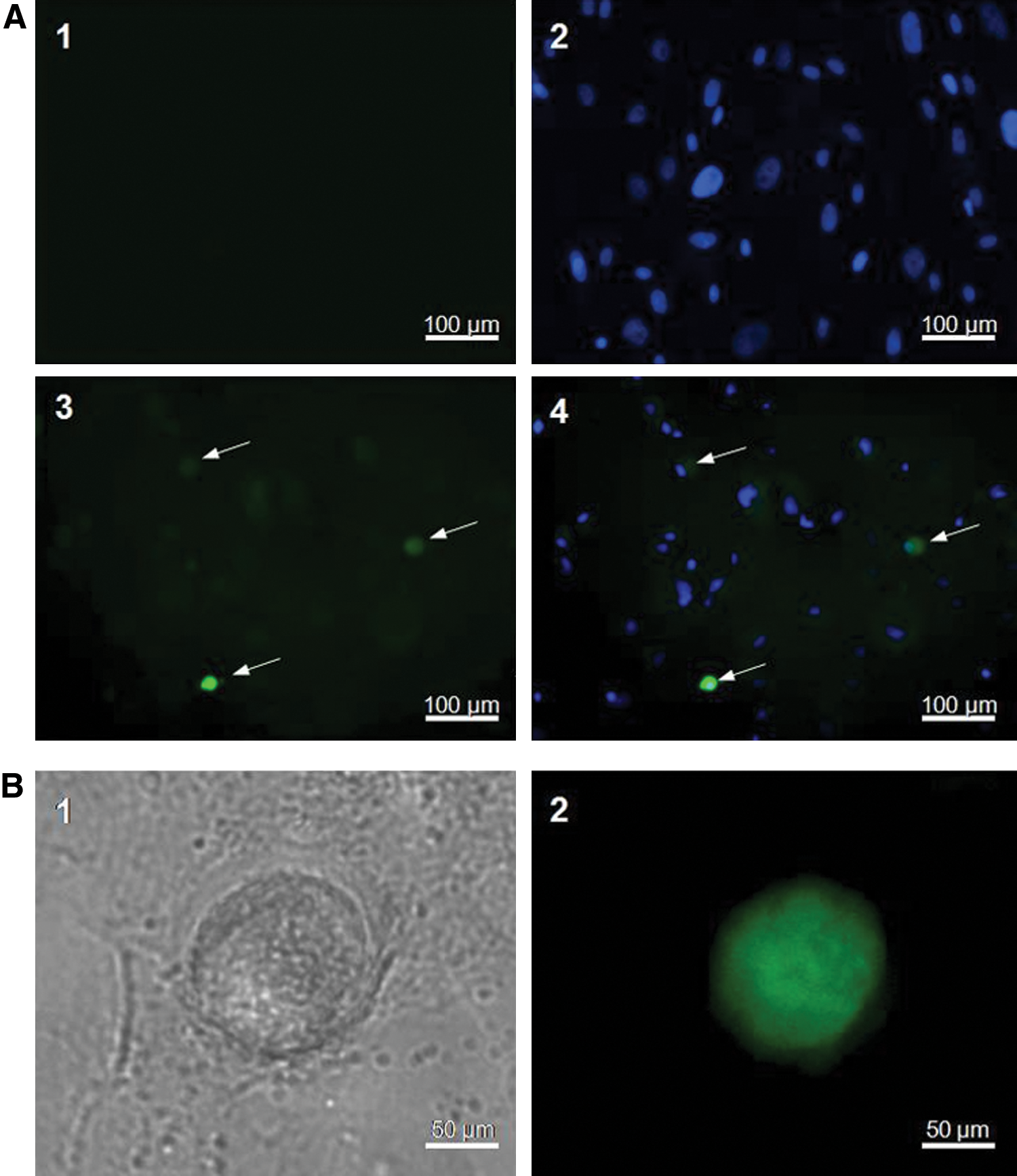
BMP15 is activated during hAFSCs differentiation into OLCs.
Analysis of oocyte specific gene expression during hAFSC induction
The OLCs derived from hAFSCs were characterized by RT-PCR to determine the expression of female gamete specific markers. In the uninduced hAFSCs, the expressions of oocyte markers including ZP1, ZP3, and c-kit were recognizable, whereas the expression of ZP2 was absent. With extending the differentiation term, ZP2 expression was detected at D10 and D15, and the mRNA level of ZP3 was also noticeably elevated. Compared with that in uninduced hAFSCs, c-kit expression was dramatically reduced in the early induction stage (D5). However, c-kit expression was recovered in the later stages of induction (D10 and D15) (Fig. 4A), which was consistent with previous reports (Hubner et al., 2003). Further, the immunofluorescence assay illustrated that the ZP2 expression was positive in the OLCs derived from hAFSCs at day 15 (Fig. 4B). Also, the induced ZP2 was colocalized with EGFP driven by BMP15 promoter in OLCs (Supplementary Fig. S2). The quantitative RT-PCR analysis verified ZP2 and ZP3 activation during hAFSCs differentiation, though the levels were still lower than those of the ovarian control (Fig. 4C). The VASA protein is specifically expressed in the germ cell lineage in both male and female sexes (Noce et al., 2001). In human, VASA is abundant in oocytes and plays a key role in oocyte function (Castrillon et al., 2000). To detect VASA expression, we conducted the western blotting analysis. The results showed that Vasa was not expressed at the early stage of differentiation, but Vasa protein was detected during the later stage of induction (D15) (Fig. 4D).
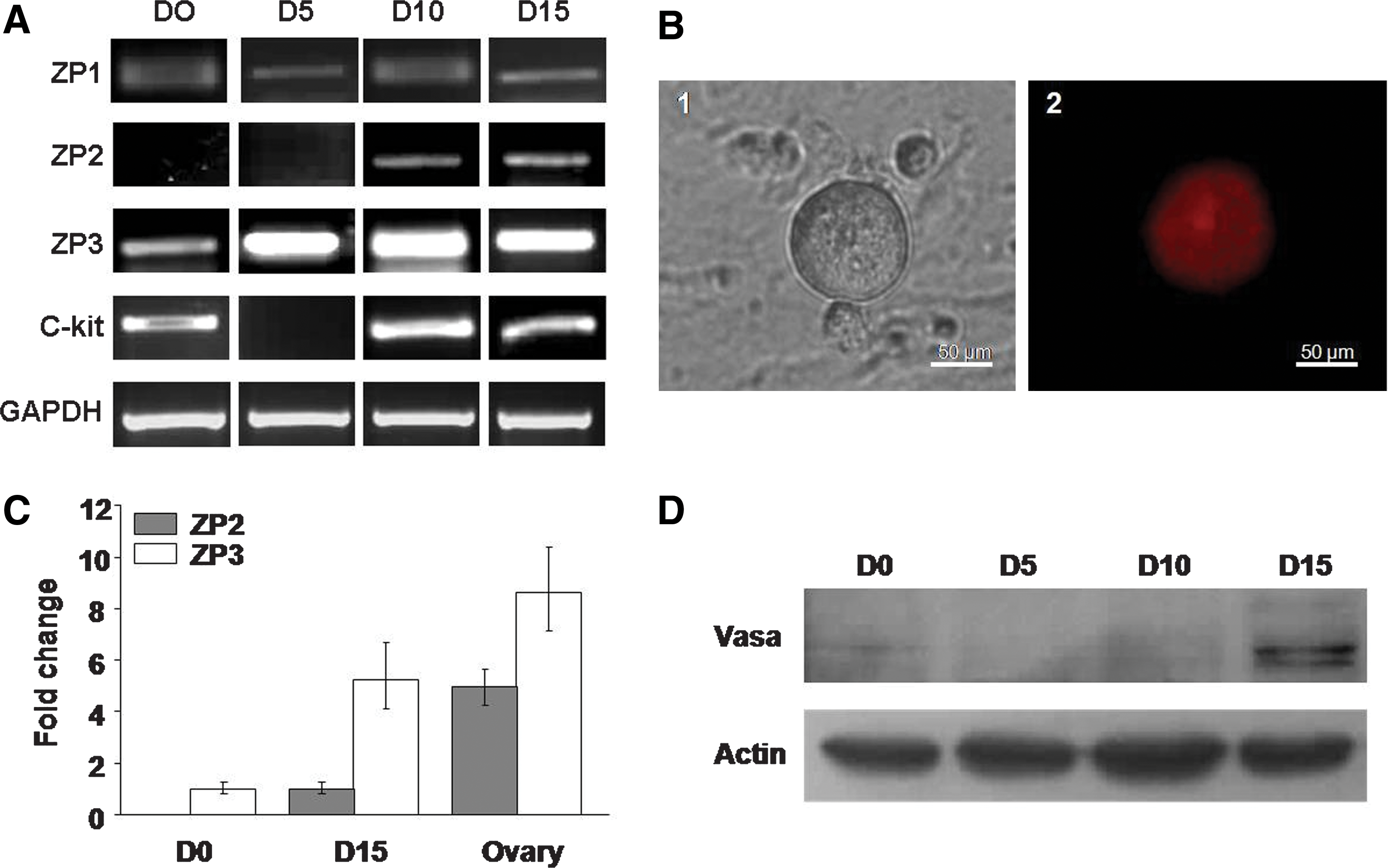
Determination of oocyte-specific markers in OLCs derived from hAFSCs. The hAFSCs were induced for 15 days, and RNA and protein were isolated from induced cells at different time points. D0–D15, cells induced from day 0 to day 15.
Discussion
Compared with the in vitro generation of oocytes from ESCs, the investigations on derivation of female gametes from somatic stem cells are still preliminary (Aflatoonian and Moore 2006; Chen et al., 2007). Several laboratories reported that OLCs could be differentiated from both mouse ESCs (Hubner et al., 2003; Lacham-Kaplan et al., 2006; Novak et al., 2006) and human ESCs (Clark et al., 2004; Kee et al., 2006; Chen et al., 2007) either through the formation of EBs or through the spontaneous differentiation method by removing the factors which promoted the pluripotency (Marques-Mari et al., 2009). Recent studies have shown that the OLCs could be derived from the somatic stem cells such as the skin-derived stem cells (Dyce et al., 2006; 2011; Linher et al., 2009) and the pancreatic stem cells (Danner et al., 2007). In this study we demonstrate that the OLCs could also be induced from hAFSCs.
Follicular fluid has been reported as one major component that facilitates differentiating stem cells into OLCs (Dyce et al., 2006). In our study we found that porcine follicular fluid in the induction medium was one crucial factor that affected hAFSCs differentiation efficiency. The fluid derived from follicles with a diameter of 2–8 mm was able to generate better results than that from the smaller or larger size of follicles. In addition, follicular fluid contains factors such as GDF9, BMP15, stem cell factor (SCF), basic fibroblast growth factor, and oestrogen (Dyce et al., 2006). These factors, which are secreted from granulosa cells, theca cells, and oocytes, play important roles in regulating the follicular development (Jiang et al., 2003; Webb et al., 2003).
Though the promoter of oocyte-specific gene GDF9 has been applied to monitor the differentiation of mouse ESCs into cells with an oocyte phenotype (Salvador et al., 2008), our data suggest that BMP15 rather than GDF9 is a specific and efficient marker to scrutinize oocyte development in hAFSCs differentiation. BMP15 is produced by oocytes and plays important roles within the ovary, such as promoting early folliculogenesis and enhancing cumulus cell expansion (Peng et al., 2009). Previous reports have revealed that BMP15 prevented precocious oocyte maturation in zebrafish to maintain oocyte quality (Clelland et al., 2006; Peng et al., 2009). In addition, c-kit (also known as CD117) is a member of the tyrosine kinase and the receptor of the SCF. c-kit binding the SCF is critical for the survival, proliferation, and development of the stem cells (Linnekin, 1999). c-kit is also used as a critical marker for identifying AFS cells (Chiavegato et al., 2007; Martin-Rendon et al., 2008; Chen et al., 2011) and an early germ lineage maker (Hubner et al., 2003). During the induction of OLCs, c-kit was dramatically reduced at first but was recovered later, which was accordant with the previous report (Hubner et al., 2003).
There are several similarities between OLCs derived from mouse ESCs (Hubner et al., 2003) and those induced from hAFSCs. For instance, both ESCs- and hAFSCs-derived OLCs can express several oocyte-specific genes such as the VASA, c-kit, ZP2, ZP3, GDF9, and BMP15. Also, both OLCs could be spontaneously activated and developed into parthenogenetic embryos at different stages. In addition, there are differences between ESCs-derived OLCs and hAFSCs-derived OLCs. One major difference is the expression pattern of GDF9 that is constantly expressed in hAFSCs, and therefore this may explain why hAFSCs differentiation into OLCs is easier and needs shorter time. In contrast, GDF9 is only detected in OLCs, but not in mouse ESCs. And BMP15 is not detected in either mouse ESCs or hAFSCs, whereas BMP15 is activated in both mouse ESCs-derived OLCs and hAFSCs-derived OLCs. Thus, we chose BMP15 instead of GDF9 as an oocyte-specific reporter during differentiating hAFSCs into OLCs. Another difference is the expression of ZP gene. The ZP that first appears in multilaminar primary oocyte is a glycoprotein membrane surrounding the plasma membrane of an oocyte. The ZP is composed of three major glycoproteins including ZP1, ZP2, and ZP3. ZP1 serves as a linker for ZP2 and ZP3. ZP2 functions as a secondary sperm receptor during fertilization and ZP3 serves as a primary sperm receptor and acrosome reaction-inducer (Wassarman et al., 2004). In the hAFSC cell line hAF-07, ZP1, and ZP3 mRNA were detectable in uninduced cells, and ZP3 expression level was significantly increased during hAFSCs differentiation into OLCs. Further, the expression of ZP2 was only detected in later induction (D10–D15), but undetectable in both uninduced and in early induced cells (D5). Similar observations were reported by Hubner et al. (2003), whereas ZP1 expression was not thoroughly observed in ESCs-derived OLCs.
In summary, our study investigated the hAFSCs differentiation into OLCs that was monitored by BMP15-EGFP reporter, and showed that OLCs derived from hAFSCs maintained multiple features of oocyte morphologically and biologically. The generation of stably transfected cell line with pBMP15-EGFP reporter is the ongoing study that may facilitate sorting OLCs through the flow cytometry. Meanwhile the physiological function of induced OLCs also needs to be investigated.
Footnotes
Acknowledgments
This work was supported by the Major Project of Chinese National Programs for Fundamental Research and Development (“973” plan, 2009CB941002 and 2011CBA1002).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.