
Abstract
Recent studies suggest that change of macrophage phenotype (M1/M2) is associated with autoimmune diseases. Sodium valproate (VPA) is a class I histone deacetylase (HDAC) inhibitor, which has immunomodulatory function in graft-versus-host disease. However, its impact on macrophage polarization has not been defined. We evaluated the effects of VPA on both mouse macrophage cell line RAW264.7 and primary mouse bone marrow macrophages (BMMs). Exposure to VPA significantly repressed the production of interleukin 12 (IL-12), and tumor necrosis factor α by lipopolysaccharide (LPS)-induced macrophage activation, in contrast, promoted IL-10 expression. VPA also affected the costimulatory molecule expression on LPS-stimulated RAW264.7 and BMMs (downregulation of CD40 and CD80, and upregulation of CD86). Specifically, VPA inhibited macrophage-mediated T helper 1 (Th1) effector but enhanced Th2 effector cell activation. Together, our preclinical study demonstrates that VPA significantly affects the phenotype and function of macrophage, indicating an important role of HDAC activity in immune regulation and inflammation. It also provides a rationale to evaluate VPA activity for the treatment of macrophage dysfunction–associated diseases.
Introduction
Histone deacetylases (HDACs) and histone acetyltransferases play critical roles in controlling the dynamics of chromatin structure by regulating histone acetylation (Johnstone, 2002; Narlikar et al., 2002), while chromatin structure alteration is essential in controlling gene transcription and the consequent biological processes of immune cells (Mosser et al., 2006; Medzhitov et al., 2007). Previous studies showed that inhibition of HDAC activity has antiinflammatory properties. HDAC inhibitors were therapeutic in mouse models of septic shock, rheumatoid arthritis, and reduced disease severity in graft-versus-host disease (GVHD) (Yi et al., 2009; Postigo et al., 2011). HDAC activities are also required for innate immune cell control of Th1 but not Th2 effector cell function (Huang et al., 2007). However, it remains unclear if histone deacetylation is involved in macrophage polarization.
Sodium valproate (VPA) is a well-known class I HDAC inhibitor (Supplementary Fig. S1; Supplementary Data are available online at
Materials and Methods
Reagents
HDAC inhibitor sodium valproate (VPA) was obtained from Sigma-Aldrich, and dissolved in ddH2O at 0.4 M and kept at −20°C. LPS was obtained from Sigma (Escherichia coli 0111:B4) and used at 100 ng/mL in cell culture. Granulocyte-macrophage colony stimulating factor (GM-CSF) (Rat) was obtained from Peprotech. ELISA kits (IL-10, IL-12p70, TNF-α, interferon-γ [IFN-γ], and IL-4) were obtained from Shanghai ExCell Biology. Phycoerythrin (PE)-conjugated-anti-mouse CD40, PE-conjugated-anti-mouse CD80, and PE-conjugated-anti-mouse CD86 were from eBioscience. TRNzol-A+ Total RNA Reagent and QuantiTect SYBR Green PCR Kit were purchased from Tiangen.
Mice and cell lines
Six- to eight-week-old female C57BL/6 and BALB/c mice were purchased from the animal center of the Shanghai Second Military Medical University. RAW264.7 cells were maintained under conditions as described in the American Type Culture Collection. Mouse bone marrow–derived primary macrophages (BMMs) were isolated from bone marrow cells in vitro according to established protocol (Weischenfeldt and Porse, 2008). BMM cells and T lymphocyte were isolated from C57BL/6 and BALB/c, respectively.
Cell culture and cell treatment
RAW264.7 macrophage cells, mouse BMMs, and T lymphocytes were cultured at 37°C in a 5% CO2-humidified incubator and were grown in DMEM supplemented with 10% FBS, penicillin (100 U/mL), and streptomycin (100 g/mL). For BMM development, the cells were cultured in the presence of GM-CSF (10 ng/mL) for 7 days. For cell treatments, RAW264.7 cells were plated at a density of 2×105 cells/well in 12-well plates and 4×105 cells/well for BMM in 6-well plates, respectively; then, cells were treated with various concentrations of VPA in the presence or absence of LPS (100 ng/mL) for 24 h, and then cells and culture supernatants were collected for measurements.
Reverse transcription and quantitative real-time polymerase chain reaction
RNA was isolated with TRIzol-A+ (Tiangen), and reverse transcriptase reaction kit (Tiangen) was used to transcribe cDNA from 3000 ng of total RNA according to the manufacture's guidance. SYBR Green Real-time PCR Master Mix (Toyobo) was used for all reactions, and quantitative real-time polymerase chain reaction (PCR) was performed with an STRATAGENE® Mx3000p (Stratagene) using reverse-transcribed cDNA as templates. The primer sequences for the target genes were as follows: for IL-12p40, sense 5′ TGGTTTGCCATCGTTTTGCTG 3′ and antisense 5′ ACAGGTGAGGTTCACTGTTTCT 3′; for TNF-α, sense 5′ GATCGGTCCCCAAAGGGAT 3′ and antisense 5′ ACTTGGTGGTTTGCTACGACG 3′; for IL-10, sense 5′ GCTCTTACTGACTGGCATGAG 3′ and antisense 5′ CGCAGCTCTAGGAGCATGTG 3′; and for β-actin, sense 5′ GGCTGTATTCCCCTCCATCG 3′ and antisense 5′ CCAGTTGGTAACAATGCCATGT 3′. For all reactions, cycling conditions were 10 min at 95°C followed by 40 repeats of 95°C for 30 s, 62°C for 30 s, and 68°C for 30 s. At the completion of cycling, melting curve analysis was performed to establish the specificity of the PCR product. Data were collected and stored in Excel format, and analyzed with Mx3000P Software version 4.0 (Stratagene). The expression of each candidate gene was internally normalized using β-actin. The relative quantitative value was expressed by the 2−ΔΔCt method (Livak et al. 2001). Each experiment was performed in duplicates and repeated three times.
Cytokine enzyme-linked immunosorbent assays
The RAW264.7 cells and BMM cells were treated with various concentrations of VPA in the presence or absence of LPS (100 ng/mL) for 24 h and then the culture supernatants were harvested. Concentrations of IL-12p70, TNF-α, and IL-10 were measured in culture supernatants by ELISA (Shanghai ExCell Biology). For IL-4 and IFN-γ detection by ELISA (Shanghai ExCell Biology), culture supernatants were harvested from various concentrations of VPA-treated BMM cells cocultured with allogeneic T cell for 48 h. Assays were performed according to the manufacturer's protocol and read at 450 nm by using a microplate reader (Bio-Rad).
Flow cytometric analysis
To analyze cell surface phenotype, the surface expressions of CD40, CD80, and CD86 were determined by flow cytometry using PE-conjugated-anti-mouse CD40, CD80, and CD86, from eBioscience. The steps were as follows: the harvested cells were washed twice with phosphate-buffered saline (PBS; containing 1% bovine serum albumin [BSA]), and centrifuged at 1000 rpm for 5 min every time. The supernatant was decanted and PBS (500 μL/tube) containing 0.5 μL PE-conjugated-anti-mouse CD40 (0.2 mg/mL), CD80 (0.2 mg/mL), and CD86 (0.2 mg/mL) was added. Then, samples were incubated in the dark at room temperature for 30 min. Samples were then centrifuged at 1000 rpm for 5 min, decanted, and washed twice with PBS (containing 1% BSA). Finally, 500 μL of PBS was added to the resuspended cells and then the cells were examined with the BD FACS Calibur. Flow cytometric data were quantitated using WinMDI 2.9 software.
Allo mixed lymphocyte reaction and MTT assays
Mouse BMMs were cultured in GM-CSF–containing DMEM supplemented with 10% FBS, penicillin (100 U/mL), and streptomycin (100 g/mL) for 6 days. At day 7, cells were seeded at a density of 2×105 cells/12-well and at day 8, various concentrations of VPA were added to the medium (containing GM-CSF) in the presence or absence of LPS (100 ng/mL) for 24 h. Then, the BMM cells were harvested and cocultured with T lymphocyte cells at a ratio 1:5 (4×103 cells/well:2×104 cells/well) in 96-well plates. After 48 h, the supernatants were harvested and IL-4 and IFN-γ were measured by enzyme-linked immunosorbent assay (ELISA). At the same time, DMEM was added to the 96-well plates to coculture BMM with T lymphocyte cells for another 20 h. Then, 50 μL of thiazolyl blue tetrazolium bromide salt (MTT) solution (5 mg/mL) was added to every well and samples were incubated at 37°C in a 5% CO2-humidified incubator for 4 h. Then, the supernatant was carefully decanted and 150 μL of dimethyl sulfoxide was added to every well and oscillated for 10 min. Finally, samples were read at 490 nm with a microplate reader and the data recorded.
Statistical analysis
Data were graphed and analyzed in GraphPad Prism 4.0 (GraphPad Software) and the p-values were determined using unpaired, two-tailed Student's t-test. Error bars represent average±standard error of the mean. The difference between two samples was considered significant if p<0.05.
Results
HDAC inhibition by VPA skews the phenotype of LPS-stimulated mouse macrophage cell line RAW264.7 from M1 to M2
As previously described, macrophage activation has been operationally defined into two separate polarization states, M1 and M2 (Mills et al., 2000). The M1 macrophage produces proinflammatory cytokines, nitric oxide synthase 2 (iNOS), and upregulates costimulatory molecules CD40 and CD80. In contrast, the M2 macrophage produces antiinflammatory cytokines and upregulates costimulatory molecule CD86. To observe the effect of VPA on regulating macrophage polarization, we assessed VPA effects on the expression of some of these genes and proteins in mouse macrophage cell line RAW264.7 by real-time PCR, ELISA, and FACS. As shown in Figures 1 and 2, VPA could block LPS-induced IL-12 and TNF-α mRNA expression and protein secretion effectively in a dose-dependent manner, but it greatly enhanced IL-10 mRNA and protein expression. IL-10 mRNA and protein expression reached peak values upon 2 mM VPA treatment. Meanwhile, we found that VPA treatment could inhibit the mRNA expression of iNOS (see Supplementary Fig. S2). The regulated expression of inflammatory mediators in macrophages is dynamic, and some genes are induced rapidly and transiently (see Supplementary Fig. S3). Subsequently, we evaluated the effects of VPA on the expression of a series of costimulatory molecules in RAW264.7 cells. Exposure to VPA significantly inhibited the expression of costimulatory molecules CD40 (Fig. 3A) and CD80 (Fig. 3B), whereas it enhanced the expression of CD86 (Fig. 3C).
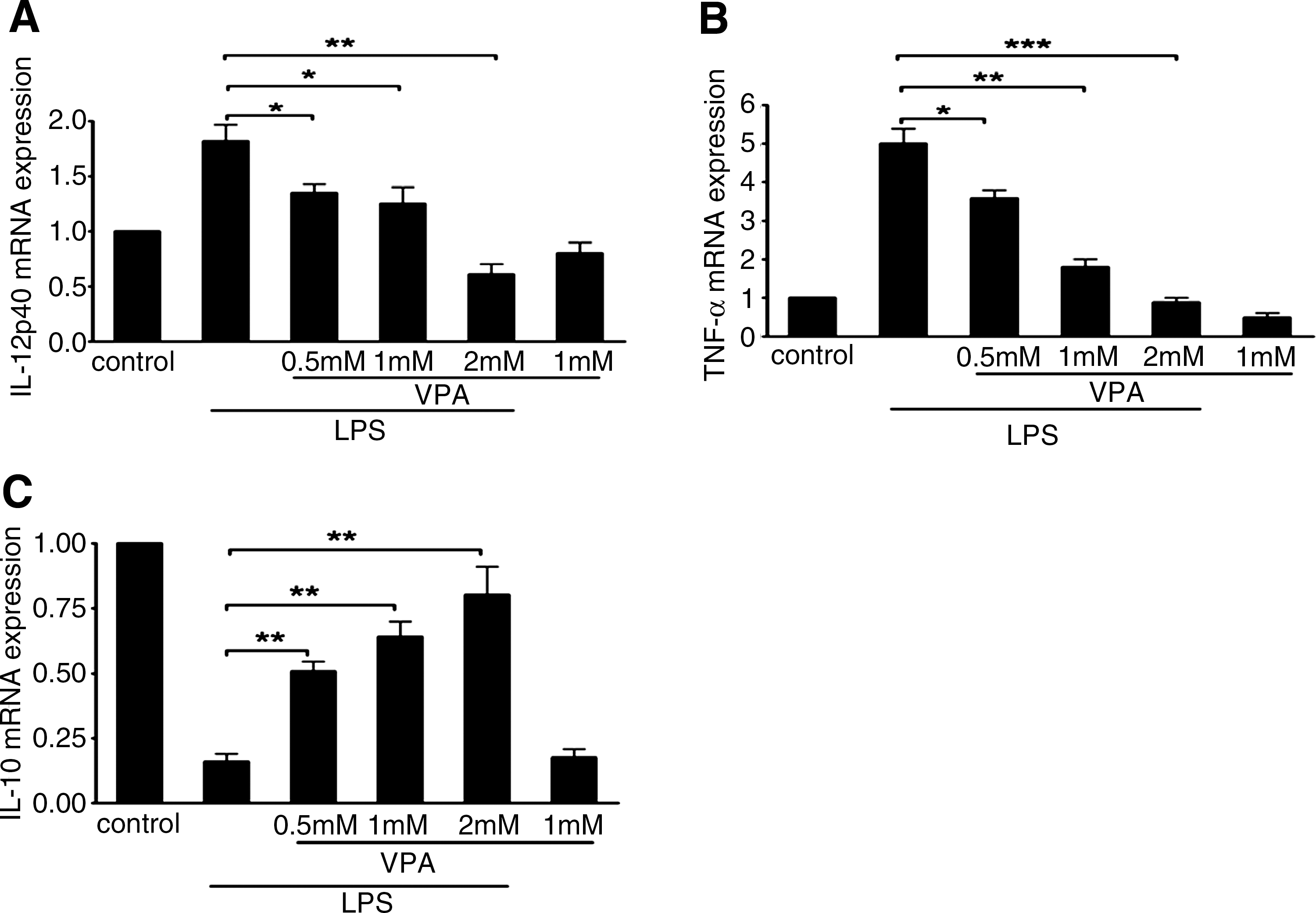
Effect of VPA on transcription of IL-12p40 TNF-α, and IL-10 in LPS-stimulated macrophage RAW264.7 cells.
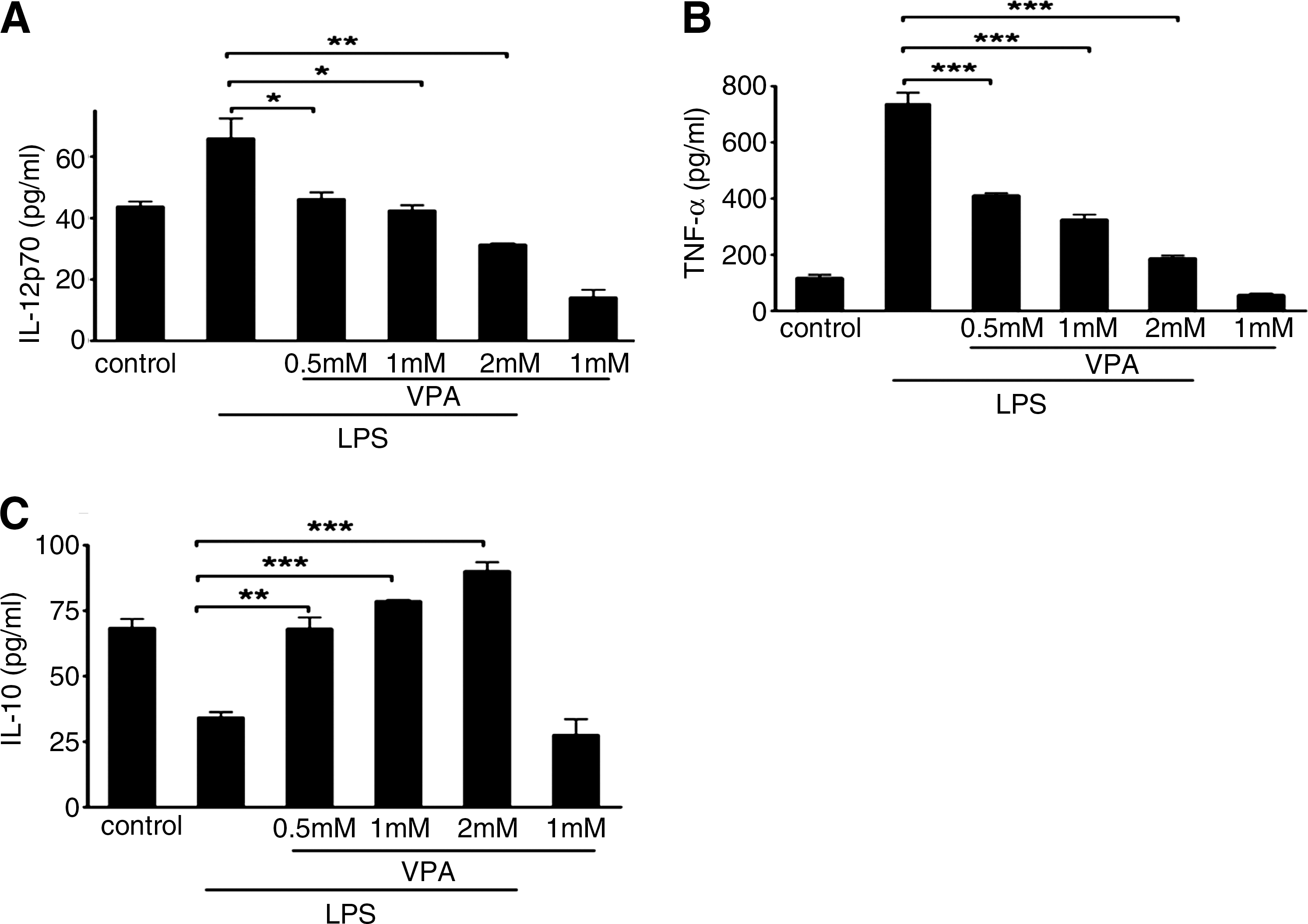
Effect of VPA on LPS-induced secretion of IL-12p70, TNF-α, and IL-10 in RAW264.7 cells.
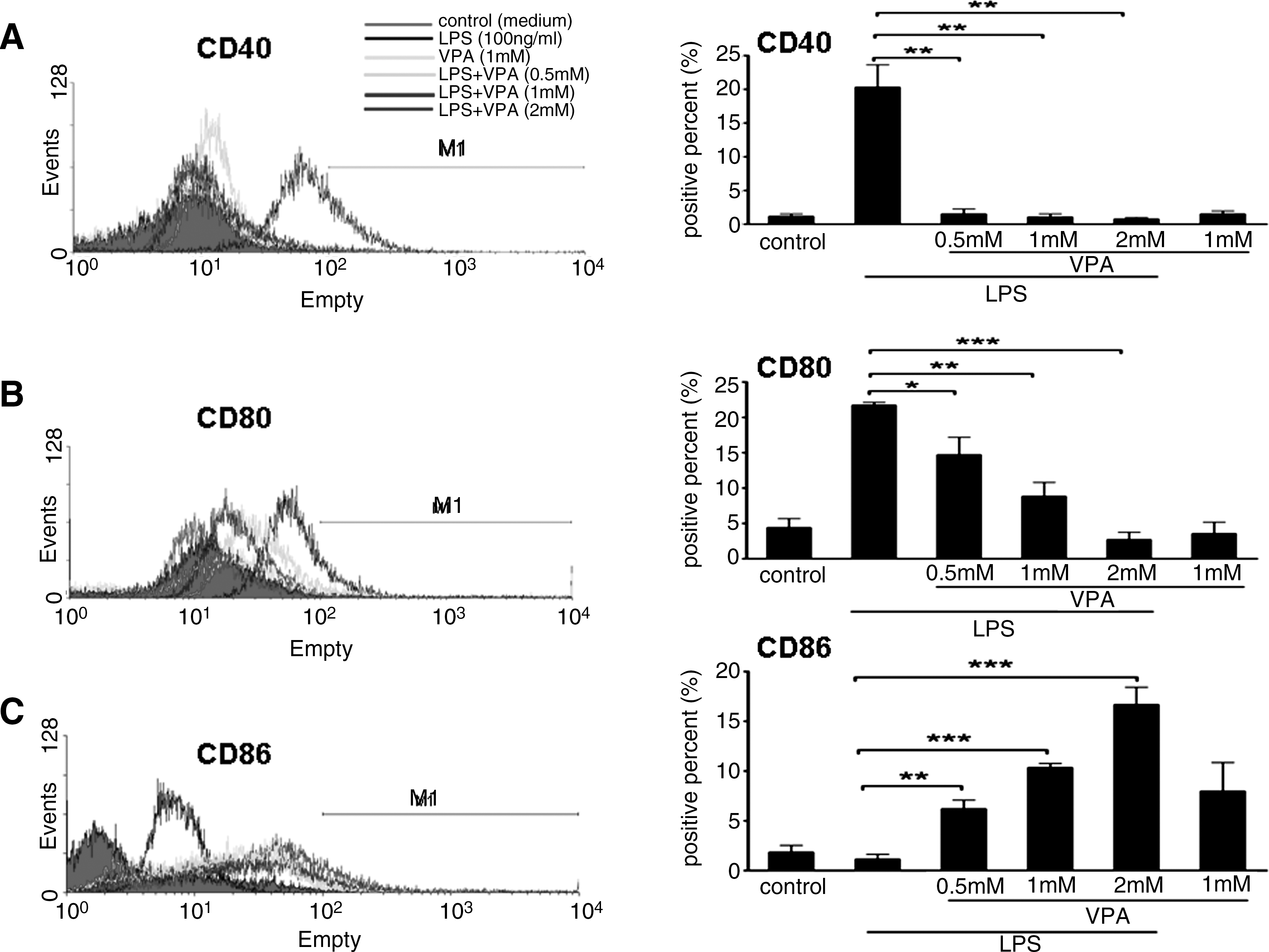
Effect of VPA on the surface phenotype of RAW264.7 cells. RAW264.7 cells were preincubated for 10–12 h with indicated concentrations of VPA or medium (control) and were then stimulated overnight with LPS (100 ng/mL) for 24 h. The control or drug-treated RAW264.7 cells were stained with phycoerythrin (PE)-conjugated-anti-mouse CD40, anti-mouse CD80, and anti-mouse CD86 monoclonal antibodies for the phenotypic analysis of CD40
HDAC inhibition by VPA alters the phenotype of LPS-stimulated mouse primary macrophages from M1 to M2
To assess whether the effect of VPA on primary macrophage polarization is similar to its effect on the RAW264.7 cell line, we next investigated the effects of HDAC inhibition on the expression of a series of costimulatory molecules and cytokines that are critical for macrophage phenotype in primary macrophages. LPS-stimulated BMMs were cultured with or without VPA for 24 h at various concentrations. Compared with the LPS-stimulated BMMs, VPA treatment (0.5, 1, and 2 mM) resulted in decreased mRNA and protein expression of IL-12 and TNF-α in LPS-stimulated primary BMM cells, but it amplified LPS-inducible IL-10 mRNA expression and protein secretion in primary BMM cells in a dose-dependent manner (Figs. 4 and 5). Similar to what was observed with the RAW264.7 cells, VPA treatment also inhibited mRNA expression of iNOS (see Supplementary Fig. S2). Meanwhile, VPA treatment at 0.5, 1, and 2 mM led to a significant downregulation of CD40 and CD80 expression on LPS-stimulated BMM cells. In contrast, we also observed the upregulation of CD86 expression on BMM cells by VPA treatment compared with LPS-stimulated BMM cells (Fig. 6).
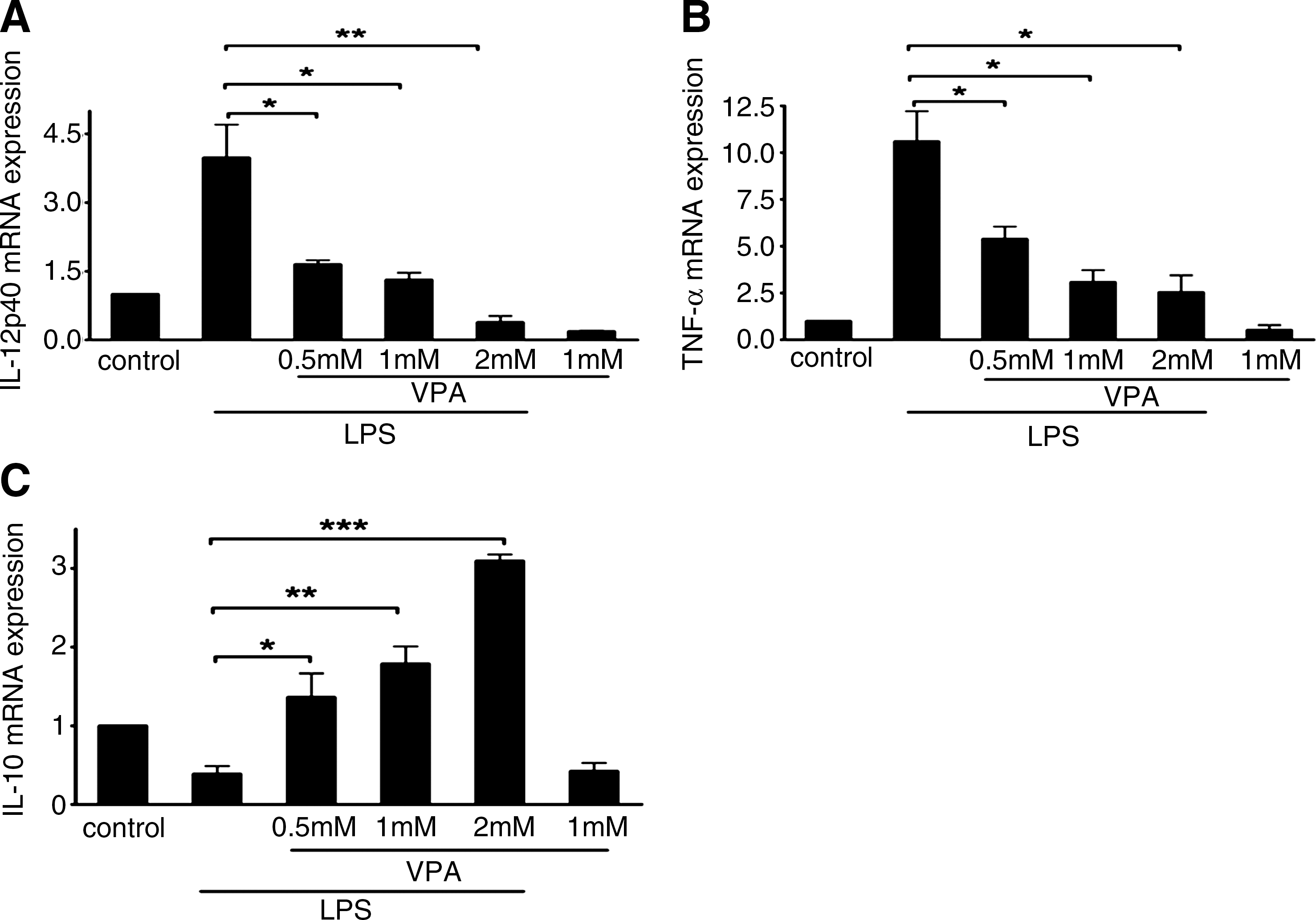
Effects of VPA on mRNA expression of cytokine genes in LPS-stimulated BMM cells.
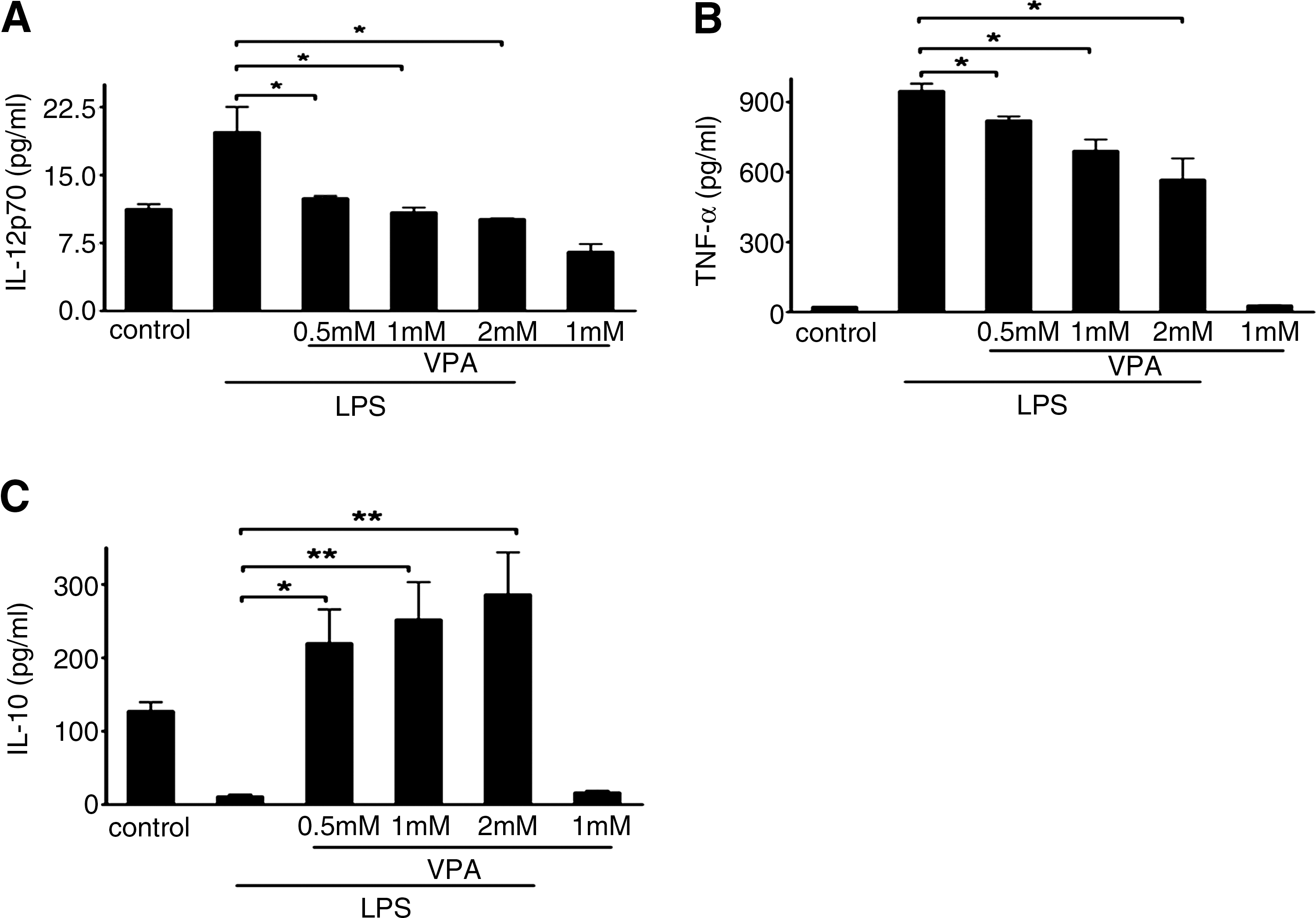
Effects of HDAC inhibition by VPA on LPS-induced secretion of cytokines by BMM cells.
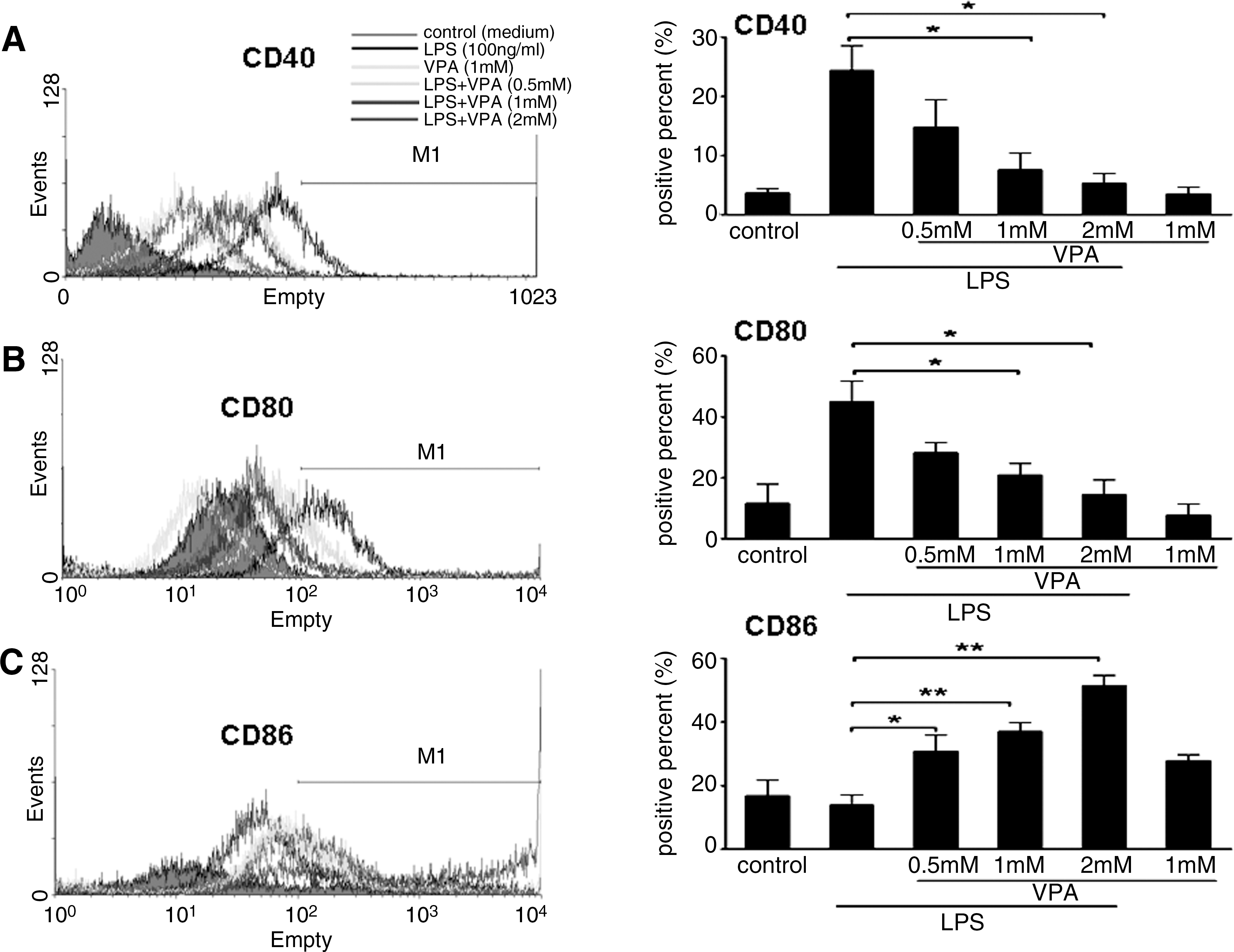
Effect of HDAC inhibition by VPA on the surface phenotype of BMM cells. BMM cells were preincubated for 10–12 h with indicated concentrations of VPA or medium (control) and were then stimulated overnight with LPS (100 ng/mL) for 24 h. The control or drug-treated BMM cells were stained with PE-conjugated-anti-mouse CD40, anti-mouse CD80, and anti-mouse CD86 monoclonal antibodies for the phenotypic analysis of CD40
HDAC inhibition by VPA is required for macrophage-mediated Th2 effector cell activation
To test whether the effects of VPA on the macrophage polarization had any functional consequences on T helper cell activation, we measured primary BMM-mediated Th1 and Th2 effector activation in a mixture lymphocyte reaction (MLR) system. First, to distinguish the proliferation of T cells, BMM cells were treated with mitomycin C (100 μg/mL) for 24 h to reduce cell viability. We cocultured BALB/c mouse–derived T cells with C57BL/6 mouse–derived BMM. BMMs were generated from C57BL/6 mice, and treated with LPS and/or VPA. BMM cells were washed extensively before being cultured with primary effector Th1 or Th2 cells derived from BALB/c mice. Coculture was carried out for 72 h, and the MTT assay was measured to monitor T-cell activation and proliferation. ConA (5 mg/L) was used to confirm the T cell viability. As shown in Figure 7A, VPA significantly reduced T cell proliferation at 72 h in a dose-dependent manner. Meanwhile, to monitor Th cell differentiation, IFN-γ or IL-4 secretion was measured by ELISA. As shown in Figure 7B, LPS treatment increased the expression of Th1 activation signals and induced more IFN-γ secretion from Th1 cells when compared with unstimulated BMM cells. However, IFN-γ secretion by CD4+ Th cells was significantly blocked in response to VPA treatment of LPS-stimulated BMM cells in a dose-dependent manner. In contrast, when Th effector cells were incubated with stimulated BMM cells, IL-4 secretion was upregulated in the presence of VPA in a dose-dependent manner (Fig. 7C). These data demonstrate that the macrophage polarization by VPA also selectively modulates macrophage-mediated Th cell differentiation. Macrophages treated by VPA could inhibit Th1 function but promote Th2 effector cell activation.
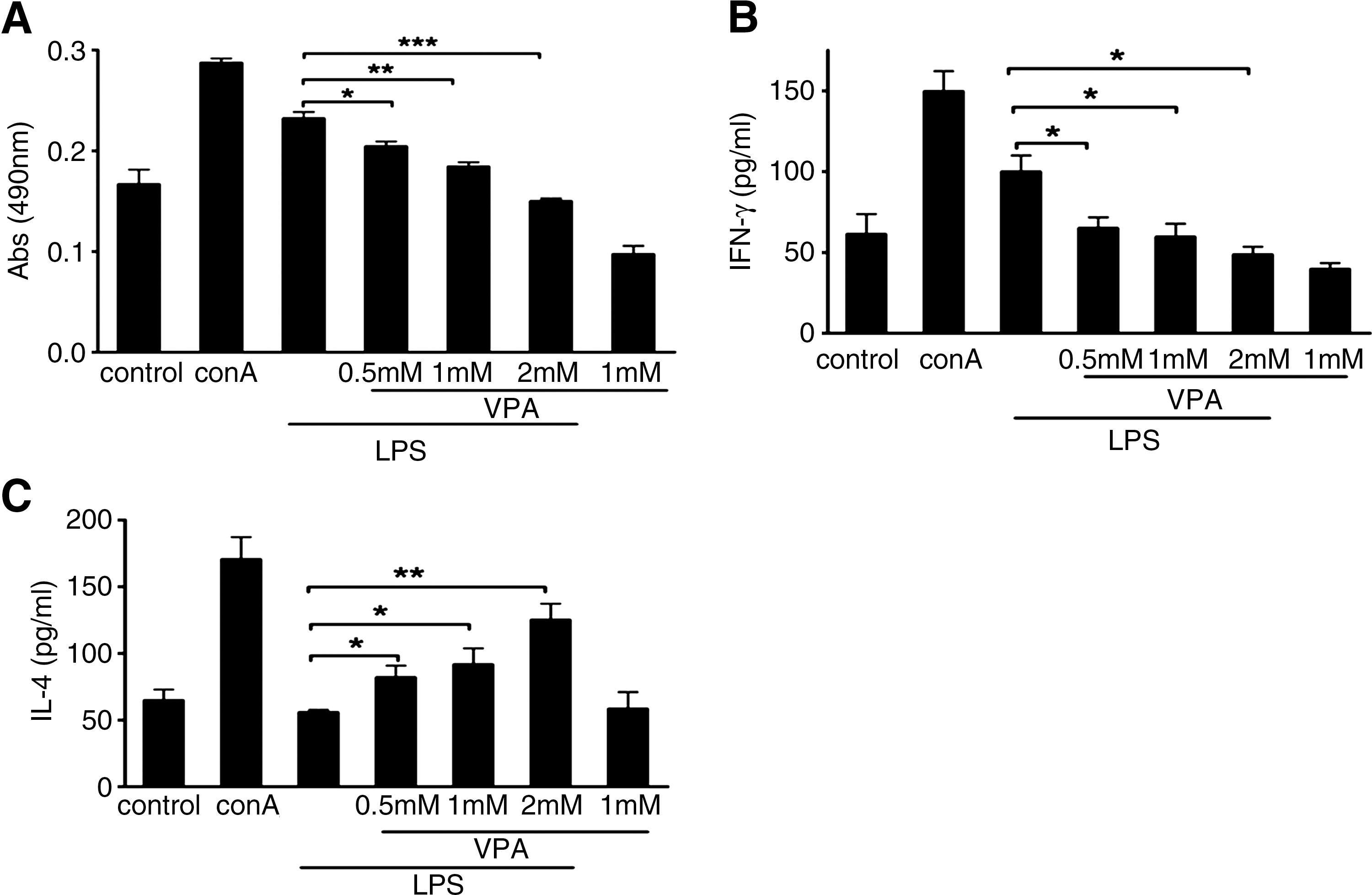
VPA modulated allogeneic T cell proliferation, interferon γ (IFN-γ), and IL-4 production in vitro. VPA was added at the indicated concentrations to LPS-induced BMM cells isolated from C57BL/6 mice that were then immediately used as stimulators in an MLR with T cells from BALB/c (allogeneic) mice as described in the Materials and Methods section.
Discussion
Macrophages are a heterogeneous population of immune cells that are essential for the initiation and resolution of pathogen- or tissue damage–induced inflammation (Gordon and Taylor, 2005). M1 (classic) macrophages produce proinflammatory cytokines, mediate resistance to pathogens, and contribute to tissue destruction, while M2 (alternative) macrophages produce antiinflammatory cytokines, promote tissue repair and remodeling, as well as ameliorate Th1-induced autoimmune disease (Medzhitov and Horng, 2009).
Histone acetylation and deacetylation are linked to cell cycle progression, and have been correlated with repair and recombination events as well as gene transcription (Marks et al., 2001). Recent discovery of the enzymes controlling histone acetylation and deacetylation confirmed that acetylation of histones is an important step in regulating gene expression for many immune processes (Skov et al., 2003). In our study, we found that VPA treatment not only inhibited HDAC activity in vivo (see Supplementary Fig. S1), but also significantly changed the phenotype and function of both mouse macrophage cell line RAW264.7 and primary BMMs.
CD40, CD80, and CD86 are important surface molecules on macrophages that are responsible for antigen-specific immune responses and mediating T-cell activation (Napolitani et al., 2005). Previous studies showed that upregulation of CD40 and CD80 reflected M1 macrophages, while upregulation of CD86 reflected M2 macrophages (Krausgruber et al., 2011). Our study found that HDAC inhibition led to the downregulation of both CD40 and CD80 and the upregulation of CD86, indicating that HDAC inhibition by VPA possibly induced macrophage phenotype shift.
IL-12, IL-10, and TNF-α are well-documented cytokine markers for macrophage phenotype (Gordon, 2003). IL-10 represses immune responses by downregulating inflammatory cytokines such as TNF-α and is important for the generation of regulatory T cells that act to suppress activation of the immune system and thereby maintain immune system homeostasis and tolerance to self-antigens (Moore et al., 2001). The main producers of IL-10 include M2 macrophages, B cells, and T cells (Li and Flavell, 2008), whereas M1 macrophages and dendritic cells (DCs) are only weak producers (Rubtsov et al., 2008). We found that VPA treatment amplified LPS-inducible IL-10 expression in both RAW264.7 and primary BMM cells. Interestingly, different from the combination of VPA and LPS treatment, VPA alone suppressed the expression of IL-10, indicating that signal pathways involved in IL-10 expression induced by VPA alone or combination of VPA and LPS may be different.
In preclinical studies that used HDAC inhibitors as immunomodulatory agents, reports of GVHD and rheumatoid arthritis were observed. Macrophage activation and phenotype determination play vital role in initiation and development of GVHD and rheumatoid arthritis, where macrophages can alter the immune response to Th1 (Yi et al., 2009; Postigo et al., 2011), but the effects of HDAC inhibitors on macrophage polarization have not been well examined. The modulation of costimulatory molecule expression and cytokine production by VPA can help to explain the considerable plasticity of macrophages in adjusting their phenotype in the therapy of GVHD (Reddy et al., 2004, 2008) and rheumatoid arthritis (Mishra et al., 2003).
Importantly, our study also demonstrated that HDAC activity had a crucial role in macrophage-mediated antigen-specific immune responses through classic (MHC-mediated) antigen-presenting pathways. In an MHC-restricted manner, DCs present peptides to activate T cells, the key mediators in the adaptive immunity. Our study showed that VPA treatment of LPS-stimulated RAW264.7 or primary BMM cells could skew the macrophage-mediated antigen-specific immune responses from Th1 to Th2. This result is consistent with the previous finding that VPA treatment can reduce experimental GVHD, which is caused by robust Th1 immune response (Reddy et al., 2004).
Conclusion
In summary, the expression pattern of VPA-induced genes involved in M1/M2 activation and the macrophage-mediated Th1/Th2 effector activation demonstrated that VPA altered LPS-stimulated macrophage polarization and reduced their capacity to activate and attract Th1 effector cells while preserving Th2 activation signals. Given that VPA is a class I HDAC inhibitor, its high efficiency in altering LPS-stimulated macrophage phenotype is significant in understanding the mechanism of macrophage polarization and development of therapeutic strategy. Our data suggest that the doses needed for immunomodulation are substantially lower than those required for antitumor efficacy. Whether reduced doses also alter macrophage immune responses in clinical trials needs to be determined.
Footnotes
Acknowledgments
This work was supported by grants from the NSFC (30971451, 90919028, and 31071306), MOST (2011CB965100, 2010CB944900, 2010CB945000, 2011CBA01100, and 2011DFA30480), and Science and Technology Commission of Shanghai Municipality (09DZ2260100, 11ZR1438500, and 11XD1405300).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.