
Abstract
Transforming growth factor-β1 (TGF-β1) is a ubiquitous and essential cytokine that plays a dual role in the development of cancer: tumor suppressor in the early stage of carcinogenesis and tumor promoter in the later stage of carcinogenesis. To date, several studies have focused on the association between the TGFB1 polymorphisms and risk of gastric cancer. Conflicting results, however, have been reported. We conducted a meta-analysis to analyze more precisely the effects of the TGFB1 C-509T, T869C, and G915C polymorphisms on the risk of gastric cancer. No significant association between the TGFB1 C-509T, T869C, and G915C polymorphisms and risk of gastric cancer was observed in overall analyses and subgroup analyses according to ethnicity.
Introduction
Transforming growth factor-beta (TGF-β), a pluripotent cytokine, plays a biphasic role in cancer development: tumor suppressor in the early stage of carcinogenesis and tumor promoter in the later stage of carcinogenesis (Blobe et al., 2000; de Caestecker et al., 2000; Massague et al., 2000; Derynck et al., 2001). TGF-β exists in 3 isoforms called TGF-β1, TGF-β2, and TGF-β3. Of the three isoforms, TGF-β1 is most frequently upregulated in tumor cells, and thus most studies focus on the role of TGF-β1 in human cancers. The TGFB1 gene is located on the long arm of chromosome 19 (19q13.1) (Yoshie et al., 2007). TGF-β1 plays an essential role in carcinogenesis, and is involved in various cellular processes including proliferation, differentiation, migration, apoptosis, angiogenesis, and immunosurveillance (Elliott and Blobe, 2005; Mishra et al., 2005). In the early stage of cancer development, TGF-β1 functions as a tumor suppressor. In the later stage of cancer development, however, TGF-β1 acts as a tumor promoter by stimulating tumor invasion or suppressing immune response (Blobe et al., 2000; de Caestecker et al., 2000; Massague et al., 2000; Derynck et al., 2001).
Emerging evidence has shown that enhanced serum concentration of TGF-β1 was detected in a wide range of human cancers, including breast cancer (Gorsch et al., 1992), lung cancer (Barthelemy-Brichant et al., 2002), prostate cancer (Ivanovic et al., 1995), colorectal cancer (Shim et al., 1999; Langenskiold et al., 2008), hepatocellular carcinoma (Song et al., 2002), and gastric cancer (Lin et al., 2006; Li et al., 2008b). The high expression of TGF-β1 has also been demonstrated in gastric cancer and in the gastric mucosa of first-degree relatives of patients with gastric cancer (Ebert et al., 2000). Overexpression of TGF-β1 is associated with the invasion, metastasis, and poor prognosis of gastric carcinoma (Maehara et al., 1999; Saito et al., 1999). Moreover, conditional loss of TGF-β signaling resulted in increased susceptibility to gastrointestinal carcinogenesis in mice (Hahm et al., 2002). These findings indicate that TGFB1 may be a candidate gene for the development of gastric cancer. Previously, three common SNPs were identified in the TGFB1 gene: one in the promoter region (C-509T), and the others in the coding region with nonsynonymous amino acid substitution (Leu10Pro, and Arg25Pro, respectively). Several studies have reported that these variations influence the production of TGF-β1 (Awad et al., 1998; Yamada et al., 1998; Grainger et al., 1999.
Many molecular epidemiological studies have been performed to investigate the association between the three polymorphisms and risk of gastric cancer. However, the results were inconclusive. For example, Jin et al. (2007) reported that the −509 CT genotype was associated with a significantly reduced risk of gastric cancer. In contrast, other studies reported that the −509 CT genotype was associated with a significantly increased risk of gastric cancer (Li et al., 2008a; Guo et al., 2011). Because a single study might have lower power to estimate the overall effects, we conducted a meta-analysis by combining data from published case–control studies to provide more analysis of the effects of the TGFB1 polymorphisms on the risk of gastric cancer.
Materials and Methods
Selection of published studies
We searched for articles using the following terms (“TGF-beta” or “TGF-β” or “transforming growth factor-beta” or “transforming growth factor-β”) and (“polymorphism”) and (“gastric cancer”) in the PubMed, Embase, China National Knowledge Infrastructure (CNKI), and Chinese Biological Medicine (CBM) databases (last search: July 13, 2011). No language restrictions were used. The inclusion criteria were: (1) articles investigating the association between TGFB1 C-509T (rs1800469), T869C (rs1800470; rs1982073; T29C; Leu10Pro), and G915C (R25P; rs1800471) polymorphisms and gastric cancer risk; (2) case–control studies; and (3) providing enough data for computing odds ratios (ORs) with 95% confidence intervals (CIs). We excluded the studies that were repeatedly published in other languages by the same research group (Zhou et al., 2007; Guo et al., 2010). Thus, a total of eight articles were included in this meta-analysis (Fig. 1). Of the eight studies, six studies investigated the association between the TGFB1 C-509T polymorphism and risk of gastric cancer, six studies investigated the association between the TGFB1 T869C polymorphism and risk of gastric cancer, and four studies investigated the association between the TGFB1 G915C polymorphism and risk of gastric cancer.
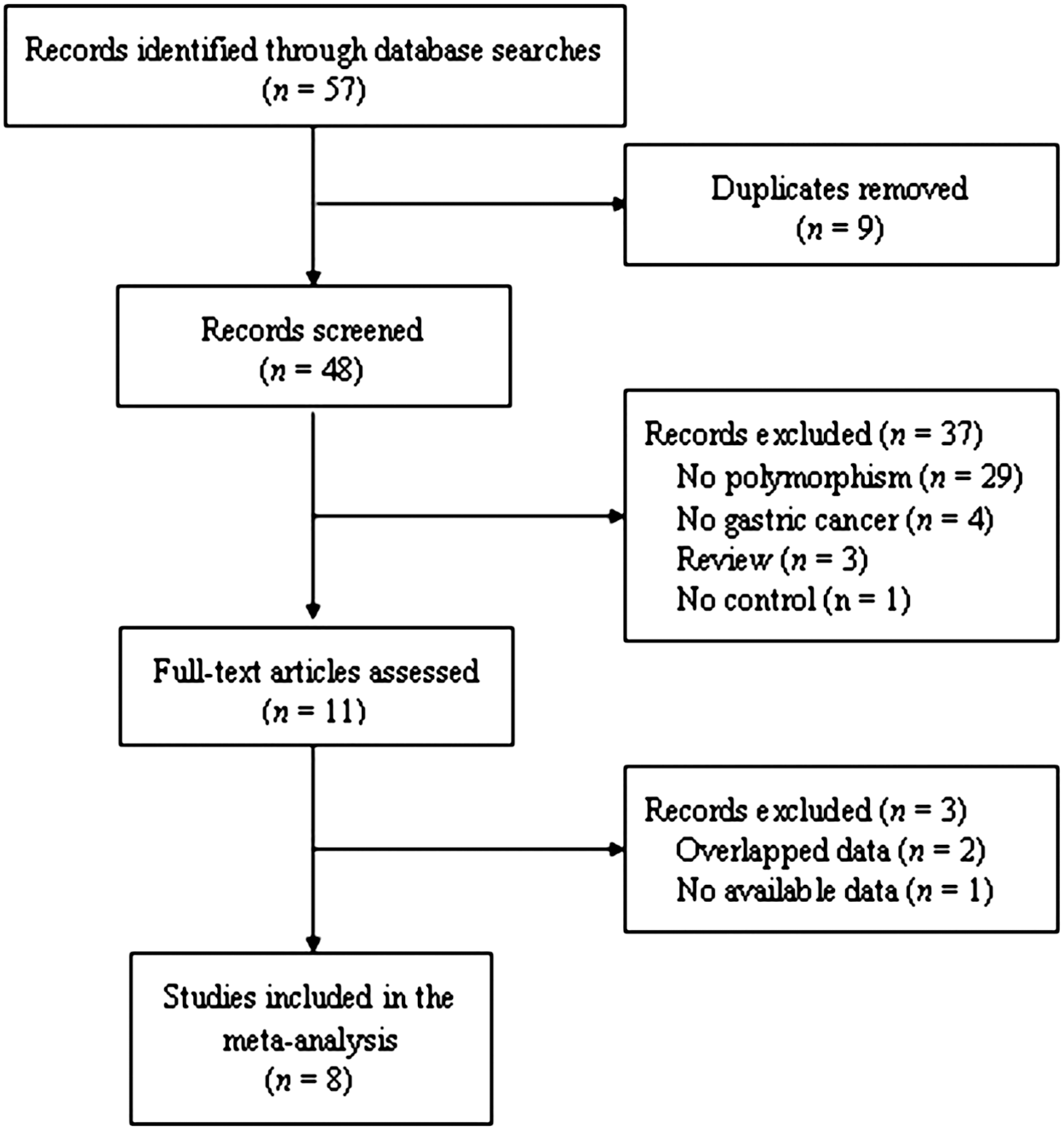
Flow diagram of the selection of eligible studies.
Data extraction
Two investigators worked independently to extract the following information from each study: the first author's name, the year of publication, country of origin, the year of sample collection, ethnicity, the number of cases and controls, genotyping methods, matching criteria, and Hardy–Weinberg equilibrium (HWE; Table 1).
PCR-RFLP, polymerase chain reaction–restriction fragment length polymorphism; HWE, Hardy–Weinberg equilibrium; PIRA-PCR, primer-introduced restriction analysis–polymerase chain reaction.
Statistical analysis
The pooled ORs and corresponding 95% CIs were performed to evaluate the association between the three polymorphisms in TGFB1 gene and gastric cancer risk. Four different ORs were calculated: (a) heterozygote comparison (C-509T: CC vs. CT; T869C: TT vs. TC; and G915C: GG vs. GC); (b) homozygote comparison (C-509T: CC vs. TT; T869C: TT vs. CC; and G915C: GG vs. CC); (c) dominant model (C-509T: CC/CT vs. CC; T869C: TC/CC vs. TT; and G915C: GC/CC vs. GG); and (d) recessive model (C-509T: TT vs. CC/CT; T869C: CC vs. TT/TC; and G915C: CC vs. GG/GC). The Q-test and I 2 statistics were used to assess the between-study heterogeneity (Higgins and Thompson, 2002). If the p-value for Q-test ≤0.10 or heterogeneity index I 2>50%, indicating obvious heterogeneity between studies, then a random-effects (the DerSimonian and Laird method) model was used to compute the pooled ORs (DerSimonian and Laird, 1986). In contrast, if the p-value for Q-test > 0.10 and heterogeneity index I 2≤50%, indicating the absence of heterogeneity between studies, then a fixed-effects (the Mantel-Haenszel method) model was used (Mantel and Haenszel, 1959). Logistic meta-regression analysis was conducted to explore potential reason for the between-study heterogeneity. Publication bias was carried out using a Begg's funnel plot (Egger et al., 1997). All the statistical analyses were done using STATA software, version 10.0 (STATA Corp., College Station, TX).
Results
A flow diagram of the selection of eligible studies is shown in Figure 1. In all, 57 records were identified through screening the PubMed, Embase, CNKI, and CBM databases. Forty-eight records remained after removing duplicates. We excluded 37 records because they were review articles (n=3) or where absence of polymorphism (n=29), gastric cancer (n=4), and control (n=1) was noted. Then, full-text articles were assessed for eligibility, and 3 records were excluded for overlapped data (n=2) or lacking available data (n=1). Finally, 8 studies, including 6 studies of the TGFB1 C-509T polymorphism (1936 cases and 2322 controls), 6 studies of the TGFB1 T869C polymorphism (1998 cases and 2562 controls), and 4 studies of the TGFB1 G915C polymorphism (861 cases and 1381 controls) were included in this meta-analysis.
The main characteristics of the eligible studies are shown in Table 1. Of the eight studies, six were of Asians, and two were of Caucasians. All the gastric cancer cases in the included studies were pathologically confirmed. Most of the studies (87.5%) performed polymerase chain reaction–restriction fragment length polymorphism analysis for genotyping. Several quality control methods were used to avoid genotyping error, including positive and negative control, internal control, random repeat, blindness to the case–control status, and different genotyping assays for validation. Three studies were found with significant deviation from HWE (Glinsky, 2008; Guan et al., 2009; Guo et al., 2011).
The main results of this meta-analysis are summarized in Tables 2 –4. No significant associa tion between the TGFB1 C-509T, T869C, and G915C polymorphisms and the risk of gastric cancer was observed in overall analyses and subgroup analyses according to ethnicity. Obvious heterogeneity was found in the effect of TGFB1 C-509T and T869C polymorphisms on the risk of gastric cancer. To explore the source of the heterogeneity, we assessed all comparison models by HWE, ethnicity, and sample size (≤500 and > 500 subjects) with meta-regression analysis. We found that the variable HWE could explain the heterogeneity (TGF-β1 T869C: TT vs. TC, and TT vs. CC). The results of the meta-analysis were not altered after exclusion of studies of Hardy–Weinberg disequilibrium (HWD).
Number of comparisons.
Random-effects model was used when p-value for heterogeneity test <0.10.
OR, odds ratio; CI, confidence interval.
Number of comparisons.
Random-effects model was used when p-value for heterogeneity test <0.10.
Number of comparisons.
Begg's funnel plot was used to evaluate the publication bias, and no evidence of publication bias was detected in this meta-analysis.
Discussion
In this study, we failed to find any association between the TGFB1 C-509T, T869C, and G915C polymorphisms and risk of gastric cancer in overall analyses. After subgroup analyses according to ethnicity, we did not find any association between the three polymorphisms in TGFB1 gene and gastric cancer in both Asians and Caucasians. Moreover, three out of eight studies showed significant deviation from HWE. When we excluded these studies of HWD, the results of the meta-analysis were not modified. These findings indicate that the TGFB1 polymorphisms may not be risk factors for the development of gastric cancer.
Several common functional polymorphisms in the TGFB1 gene have been reported. One is the C-509T polymorphism located in the promoter region of the TGFB1 gene. The −509 T allele was reported to be associated with increasing levels of TGF-β1 compared with the −509 C allele (Grainger et al., 1999). Moreover, the variation could alter TGFB1 transcription activity and the interaction with the transcription factor Yin Yang 1 (Silverman et al., 2004). Two additional polymorphisms, located in codon 10 (Leu→Pro) and codon 25 (Arg→Pro) of the TGFB1 gene were reported to be associated with interindividual variation in levels of TGF-β1 production (Awad et al., 1998; Yamada et al., 1998; Grainger et al., 1999). Given the functional relevance, several epidemiological studies have focused on the association between the TGFB1 polymorphisms and risk of gastric cancer.
In the last decades, evidence of the relationship between TGF-β1 and gastric cancer has been provided: in patients with gastric cancer, elevated expression of TGF-β1 was detected (Lin et al., 2006; Li et al., 2008b), and the high expression was related to the progression and invasion of tumor (Maehara et al., 1999; Saito et al., 1999); in mice model, loss of TGF-β activity led to increased susceptibility to gastrointestinal tumorigenesis (Hahm et al., 2002). However, no significant association between the TGFB1 polymorphisms and risk of gastric cancer was observed in this meta-analysis. This finding was confirmed by two recent genome-wide association studies that did not identify TGFB1 as the susceptibility gene of gastric cancer (Sakamoto et al., 2008; Abnet et al., 2010).
Obvious between-study heterogeneity was detected in this meta-analysis. Logistic meta-regression analysis, therefore, was performed to explore possible reasons for the heterogeneity. After taking HWE, ethnicity, and sample size into consideration, we found that the variable HWE but not ethnicity, and sample size could explain the heterogeneity. To check whether HWD could influence the results of this meta-analysis, we recomputed the summary ORs omitting these studies (Glinsky, 2008; Guan et al., 2009; Guo et al., 2011). No significant association was found, suggesting the results were robust.
Several limitations of this study need to be addressed. First, obvious heterogeneity among studies was observed in overall analyses and also subgroup analyses, which is likely to result in misleading results. Second, the relatively small sample size included in this meta-analysis might not provide sufficient power to assess the relationship between the TGFB1 polymorphisms and gastric cancer risk. Finally, further evaluation was prevented due to lack of available data on the effect of gene–gene, gene–environment interactions on gastric cancer.
In conclusion, this meta-analysis provides evidence that TGFB1 C-509T, T869C, and G915C polymorphisms are not associated with the development of gastric cancer. Well-designed studies with thousands of individuals are warranted to confirm these findings. Further studies investigating the effect of gene–gene and gene–environment interactions on gastric cancer risk would also be valuable.
Footnotes
Disclosure Statement
Niu H and Chen ZL designed the study and wrote the article. Niu Z, Zhang XL, and Chen ZL collected the data, performed the statistical analysis, and interpreted the results. All authors read and approved the final article.