
Abstract
We identified extracellular signal-regulated kinase 2 (erk2) from green mud crab, Scylla paramamosain, in this article. It was originally identified from an expressed sequence tag fragment from a normalized gonadal cDNA library. 5′ Rapid amplification of cDNA end (RACE) technique was used to obtain the 5′ untranslated region (UTR). The full-length cDNA of Sp-erk2 is 1516 bp, including a 5′-terminal UTR of 19 bp, an open-reading frame of 1098 bp, and a 3′-terminal UTR of 399 bp. The translated protein is 365 amino acids in length with a predicted molecular weight of 42 kDa, which is the same as other species. It is the first time that the expression of Sp-erk2 in different stages of ovary development of crustacean was analyzed, and the result showed that the expression of Sp-erk2 increased gradually with ovarian development, with a peak in the mature phase. In situ hybridization histochemistry was used to clarify the detail of expression. Positive signals illustrated that Sp-erk2 mRNA is present in follicular cells when the ovary is in early stages, and in both follicular cells and oocytes when it is in mature phases. All above suggest that Sp-erk2 is important for ovarian development.
Introduction
Green mud crab has been studied since the 20th century. The research on green mud crab ovary was mainly focused on the structure and development (Li and Wang, 2001; Tang et al., 2009; Islam et al., 2010), and study on molecular level was very limited. Three normalized cDNA libraries from ovary, testis, and gonad development-related organs of green mud crab were constructed by our research group (Zou et al., 2007, 2009, 2011), and then a total of 5160 high-quality expressed sequence tags (ESTs) were obtained and a home-made cDNA microarray containing 5664 spots was developed from these libraries. The results from EST annotation, cDNA microarray hybridization, and real-time PCR analysis indicated that cell cycle regulatory protein, mitogen-activated protein kinase (MAPK) signal transduction pathway, and ubiquitin proteasome pathway may participate in the ovarian development of green mud crab (Zou et al., 2011). Meanwhile, our group cloned a vasa gene from green mud crab and preliminarily analyzed its expression during gonadal development and gametogenesis, with a result that indicated that vasa gene likely serves as a useful and specific marker for germ cell development (Wang et al., 2011). In addition, some differential expression genes in the thoracic ganglion of green mud crab during ovarian development have been obtained recently from an SSH cDNA library (Zeng et al., 2011).
MAPK signal transduction pathway is a conserved signal transduction pathway in eukaryotes that exists from yeast to human and is widely involved in cell growth, differentiation, reproduction, apoptosis, antistress, and other physiological processes (Schaeffer and Weber, 1999; Widmann et al., 1999). It transduces signals from outside the cell to the cytoplasm or nucleus by a conserved three-kinase cascade. This cascade consists of a MAPK, which is activated via phosphorylation by a MAPK kinase, which in turn is phosphorylated by a MAPK kinase kinase, phosphorylating intracellular substrate proteins or transcription factors to regulate physiological processes (Schaeffer and Weber, 1999; Chang and Karin, 2001; Orton et al., 2005). At least four subfamilies of MAPK—extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinases, p38 MAPK (p38), and ERK5—have been defined in mammalian cells (Cowan and Storey, 2003; Qi and Elion, 2005). As the first discovered MAPK pathway, ERK signal transduction pathway is activated by a variety of mitogenic stimuli, such as growth factors, cytokines, and phoebe, which regulate proliferation, differentiation, meiosis, fertilization, embryonic development, metamorphosis, learning and memory, cell survival, and so on (Kumano et al., 2001; Lambert and Nagy, 2003; Akanuma and Nishida, 2004; Sharma and Carew, 2004; Feld et al., 2005; Chambon et al., 2007; Junttila et al., 2008).
As is known, meiosis is necessary for producing haploid gametes and ERK pathway is required for this process. Function of ERK on meiosis of oocyte has been studied for decades in certain species, which include at least four aspects in starting the meiosis resumption, spindle organization, meiosis I–meiosis II (M I–M II) transition, and M II arrest as a component of cytostatic factor (Schmitt and Nebreda, 2002; Fan and Sun, 2004).
ERK2—also termed ERK, MAPK, or MAPK1 in other articles—is the main molecule in ERK signal transduction pathway. Since the first erk gene was cloned by Boulton et al. in 1991 (Boulton et al., 1991), ERK transduction pathway has been well studied in many species, especially the regulation mechanism during oocyte development and ovarian pathology in mammals as a pivotal signal transduction pathway (Fan and Sun, 2004; Steinmetz et al., 2004). Although erks have been cloned from a great number of vertebrates, the information is inadequate in invertebrates. Up to date, in aquatic invertebrates, erks have been cloned from Strongylocentrotus purpuratus, Ciona intestinalis, Marthasterias glacialis, Aplysia californica, Marsupenaeus japonicus, and partial sequence from Littorina littorea (Iakovleva et al., 2006). Moreover, research on the action of ERK signal transduction pathway in ovarian development in crab is unavailable. In this article, the Sp-erk2 full-length cDNA from S. paramamosain was cloned and its differential expression in developing ovary was also described. These results may allow us to gain more insight into the function of erk2 in ovarian development of crustaceans.
Materials and Methods
Animals' and materials' preparation
Green mud crabs were bought mainly from a crab farm in Zhangpu, Zhangzhou, P.R. China. Tissues from ovary, muscle, gill, heart, brain, stomach, intestine, and hepatopancreas were rapidly dissected, snap-frozen in liquid nitrogen, and stored at −80°C for later experiments. In addition, ovaries for in situ hybridization histochemistry were immersed into 4% paraformaldehyde in phosphate-buffered saline (pH 7.4) overnight at 4°C, treated with gradient methanol, and finally stored in 100% methanol at −80°C for use. The gonad somatic index (GSI=gonad weight/body weight)—the female crab was classified into six stages: stage 1 (GSI=0.7±0.3), stage 2 (GSI=2.0±0.1), stage 3 (GSI=3.5±0.2), stage 4 (GSI=5.2±0.2), stage 5 (GSI=7.6±0.6), and stage 6 (GSI=10.3±0.3)—described in several publications (Lo et al., 2007; Wonglapsuwan et al., 2010; Zhang et al., 2010) was adapted in this study.
RNA isolation and cloning of the full-length cDNA of Sp-erk2
RNA isolation and reverse transcription PCR
Total RNA from tissues obtained previously was isolated with home-made trizol reagent according to methods described previously (Zhang et al., 2003), and the concentration of the total RNA was calculated by measuring the absorbance at 260 nm. RQ1 RNase-Free DNase I (Promega) was used to eliminate the contaminating DNA. Three micrograms of total RNA of each sample was reverse transcribed using random primers and M-MLV reverse transcriptase (Promega). Lastly, the PCR products were 10-fold diluted in ultrapurified water for final use.
Cloning the 5′ untranslated region of Sp-erk2 cDNA
A partial sequence of 1416 bp that was identified as a 3′ sequence homologous to erk2 was obtained from a normalized gonadal cDNA library of green mud crab (Zou et al., 2011). 5′ Rapid amplification of cDNA end (RACE) method was used to amplify the 5′ end of Sp-erk2, and it was carried out by using SMART™-RACE cDNA amplification kit (Clontech) according to the manufacturer's instructions. Sp-erk2-out and Sp-erk2-inner primers (Table 1) were designed based on the selected fragments. With Sp-erk2-out and universal primer mixture (UPM), the PCR programs were carried out at 94°C for 3 min, followed by 5 cycles of 94°C for 30 s and 72°C for 2 min; 5 cycles of 94°C for 30 s, 69°C for 30 s, and 72°C for 2 min; 25 cycles of 94°C for 30 s, 67°C for 30 s, and 72°C for 2 min; and an extension step at 72°C for 10 min. Then, a nested PCR was performed using the primer Sp-erk2-inner and the nested universal primer (NUP), using a program at 94°C for 3 min, followed by 35 cycles of 94°C for 30 s, 68°C for 30 s, and 72°C for 2 min, and a final extension step at 72°C for 10 min. Both UPM and NUP primers were provided by the kit. PCR products were purified with a Qiaquick PCR Purification Kit (Qiagen) and inserted into pMD18-T (TaKaRa) and sequenced. To check the sequence, a pair of full-length primers (Sp-erk2-head and Sp-erk2-toe; Table 1) were designed and head-to-toe PCR was carried out with three different cDNA templates.
Bioinformatic analysis
Nucleotide and predicted amino acid sequence data were compiled and aligned with sequences in GenBank databases using BLASTn and BLASTx with default parameters (
Real-time quantitative PCR
The expression levels of Sp-erk2 in crab tissues and different stages of ovaries were analyzed by real-time quantitative PCR. Primers for Sp-erk2 (Sp-erk2-F and Sp-erk2-R; Table 1) and 18S rRNA (18S rRNA-F and 18S rRNA-R; Table 1) were designed using Primer Premier 5 software and tested to ensure amplification of single discrete bands with no primer dimers. An aliquot of 3 μg of RNA pretreated with DNase I was used as template for cDNA synthesis in 20 μL reactions with random hexamers according to the M-MLV RT user information (Promega). The 20 μL reaction volume of quantitative real-time PCR contained 8 μL of 100-fold diluted original cDNA, 10 μL of 2× SYBR Green Master Mix (Applied Biosystems), and 1 μL of each primer (10 pM/μL) and analyzed in the ABI 7500 Real-Time System. The original cDNAs were 20,000-fold diluted for preparing the templates for 18S rRNA. The cycling program for both Sp-erk2 and 18S rRNA was as follows: 1 min at 95°C, followed by 40 cycles of 15 s at 95°C and 1 min at 60°C. Melting curves were also plotted (60°C–90°C) to make sure that a single PCR product was amplified by each pair of primers. PCR products for Sp-erk2 and 18S rRNA were ligated into plasmids and transformed into Escherichia coli JM109–competent cells. Minipreps of isolated plasmid DNA were then prepared for sequencing to check the sequence of the real-time PCR products. The comparative threshold cycle (CT) method (user Bulletin#2, the ABI PrismR 7500 Sequence detector; PE Applied Biosystems) was used to calculate the relative concentrations. This method involves obtaining CT values for the Sp-erk2; normalized to the housekeeping gene, 18S rRNA; and compared with the relative expression level. Experiments were performed routinely with at least three crabs each stage with values presented as 2−ΔΔCT for the expression levels of Sp-erk2 normalized with 18S rRNA (ΔCT=CT of Sp-erk2−CT of 18S rRNA, ΔΔCT=ΔCT of challenged sample−ΔCT of calibrator sample). Data were expressed as mean and standard error of the mean unless otherwise stated. Statistical analysis of the normalized CT values was performed with one-way analysis of variance and Student's t-test using SPSS 18.0 program. Differences were considered significant at p<0.05 (two-tailed test). In the analysis of gene expression pattern in crab developing ovary, stage 1 ovary was chosen as the calibrator.
In situ hybridization histochemistry
Sp-erk2 digoxigenin-labeled RNA probe synthesis
A pair of primers (Sp-erk2-ISH-F and Sp-erk2-ISH-R; Table 1) were designed for Sp-erk2 DIG-labeled RNA probe synthesis. A common PCR with an annealing temperature at 60°C was conducted and the products were cloned into the pGEM-T Easy Vector (Promega), and then the ligation mixture was transformed into E. coli JM109 competent followed by sequencing. Then, the plasmids were isolated and linearized with T7 and SP6 primers. After purification, the cDNA was subjected to in vitro transcription using SP6/T7 RNA polymerase and DIG RNA Labeling Mix (Roche Applied Science) following the user's manual and quantitated via spectrophotometer. The DIG-labeled probe was subpackaged and stored at −80°C.
Tissue preparation and in situ hybridization
Tissues preserved in methanol were translocated in turn into ethanol, xylene, and four grades of hot paraffin to complete paraffin embedding. After cooling, the wax block was sliced to 4 μm and transferred to slides (pretreated with polylysine). Tissue sections were bathed in xylene and then in gradient ethanol to rehydrate just before hybridization. Prehybridization, hybridization, and posthybridization were performed according to the general protocol of DIG RNA labeling kit (Roche). The concentration of both sense and antisense DIG-labeled RNA probe was 1 ng/μL for hybridization. An anti-DIG antibody was used to react with the DIG-labeled RNA probe. NBT/BCIP was used to detect the signal by dyeing the substrates. The hybridized sections were observed and photographed under an automatic digital microscope (Motic BA600Mot-7.5) and the pictures were retouched with Motic VM V1 Viewer as well as Adobe Photoshop CS5.
Results
Cloning and sequence analysis of Sp-erk2
Full-length cDNA of Sp-erk2 was cloned by 5′ RACE based on the EST sequence in the cDNA library and was verified through head-to-toe PCR. The full-length cDNA of Sp-erk2 is 1516 bp, with an 18 bp polyA tail. The ORF contains 1098 bp that encodes a protein with 365 amino acids. Its 5′ untranslated region (UTR) and 3′ UTR contain 19 and 399 bp, respectively. The GenBank accession number is GQ847862 (Fig. 1). The predicted molecular mass of ERK2 is 42 kDa, with a theoretical pI of 5.98. It is a nonsecretary protein since the SingalP 3.0 server showed that the ERK2 lacks an amino-terminal signal peptide. There are 22 predicted phosphorylation sites in the amino acid sequence with 7 serines, 8 threonines, and 7 tyrosines. Conserved domain finder suggested that 17–329 amino acid residues are the catalytic domain of ERK1 and ERK2-like serine/threonine kinases, including an active site, an ATP binding site, a substrate binding site, an activation loop (A-loop), and a kinase interaction motif docking site that appears to be conserved in all MAPKs. Moreover, there is a TEY motif located in the A-loop, in which Thr183 and Tyr185 are dual-phosphorylation sites (Fig. 1).

The full-length cDNA of Sp-erk2 and its deduced amino acid sequences. Amino acid residues in ovals are phosphorylation sites; sequence in shadow is the catalytic domain of extracellular signal-regulated kinase 1 and 2-like serine/threonine kinases; amino acid residues underlined with dotted line are the activation loop (A-loop); the black T and Y are the dual-phosphorylation sites in the TEY motif. erk2, extracellular signal-regulated kinase 2.
A phylogenetic tree was constructed by MEGA 4.0 software using NJ method to investigate the relationship of ERK2 of several species from human to aquatic invertebrates (Fig. 2). The phylogenetic tree shows that mammalian, avian, amphibian, and fish form one cluster; arthropod forms another cluster; and these two clusters together finally. Chordate was relatively distant and echinodermata forms a branch independently. Protein multiple-alignment results (Fig. 3) demonstrate that it is very highly conserved among ERK2 proteins. Sequence of M. japonicus that belongs to crustacea shows the highest similarity to Sp-erk2 (94% identity).
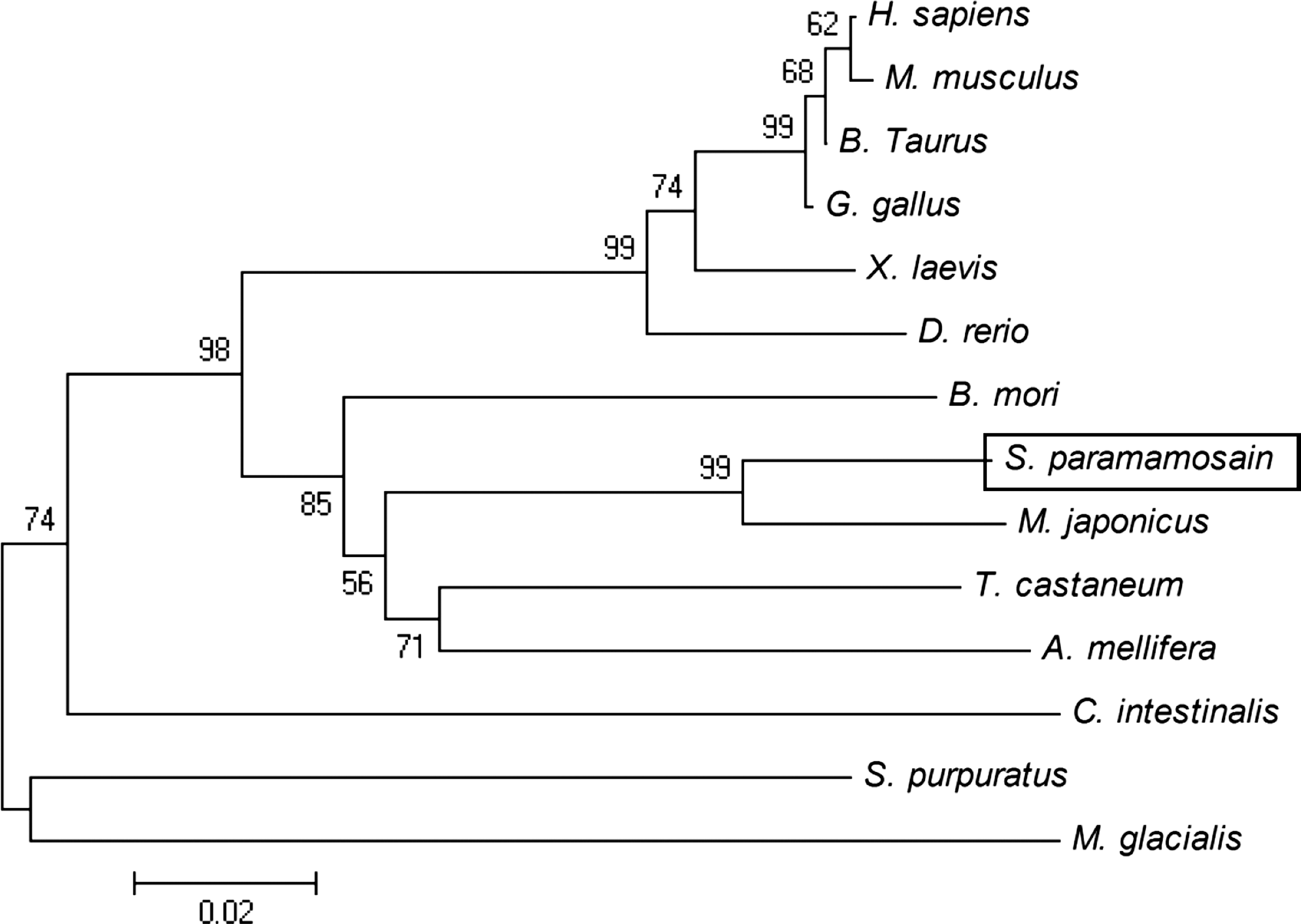
Phylogenetic analysis of Sp-ERK2. A phylogenetic tree was structured based on the ERK2 amino acid sequence of several species from human to aquatic invertebrates as follows: Scylla paramamosain, ACX32460.1; Marsupenaeus japonicus, BAH86598.1; Tribolium castaneum, XP_966833.1; Strongylocentrotus purpuratus, NP_999813.1; Apis mellifera, XP_393029.1; Bombyx mori, NP_001036921.1; Xenopus laevis, NP_001083548.1; Bos taurus, NP_786987.1; Homo sapiens, NP_620407.1; Gallus gallus, NP_989481.1; Mus musculus, EDK97436.1; Danio rerio, AAH65868.1; Ciona intestinalis, NP_001071697.1; Marthasterias glacialis, CAD60453.1. The number near node represents bootstrap values.
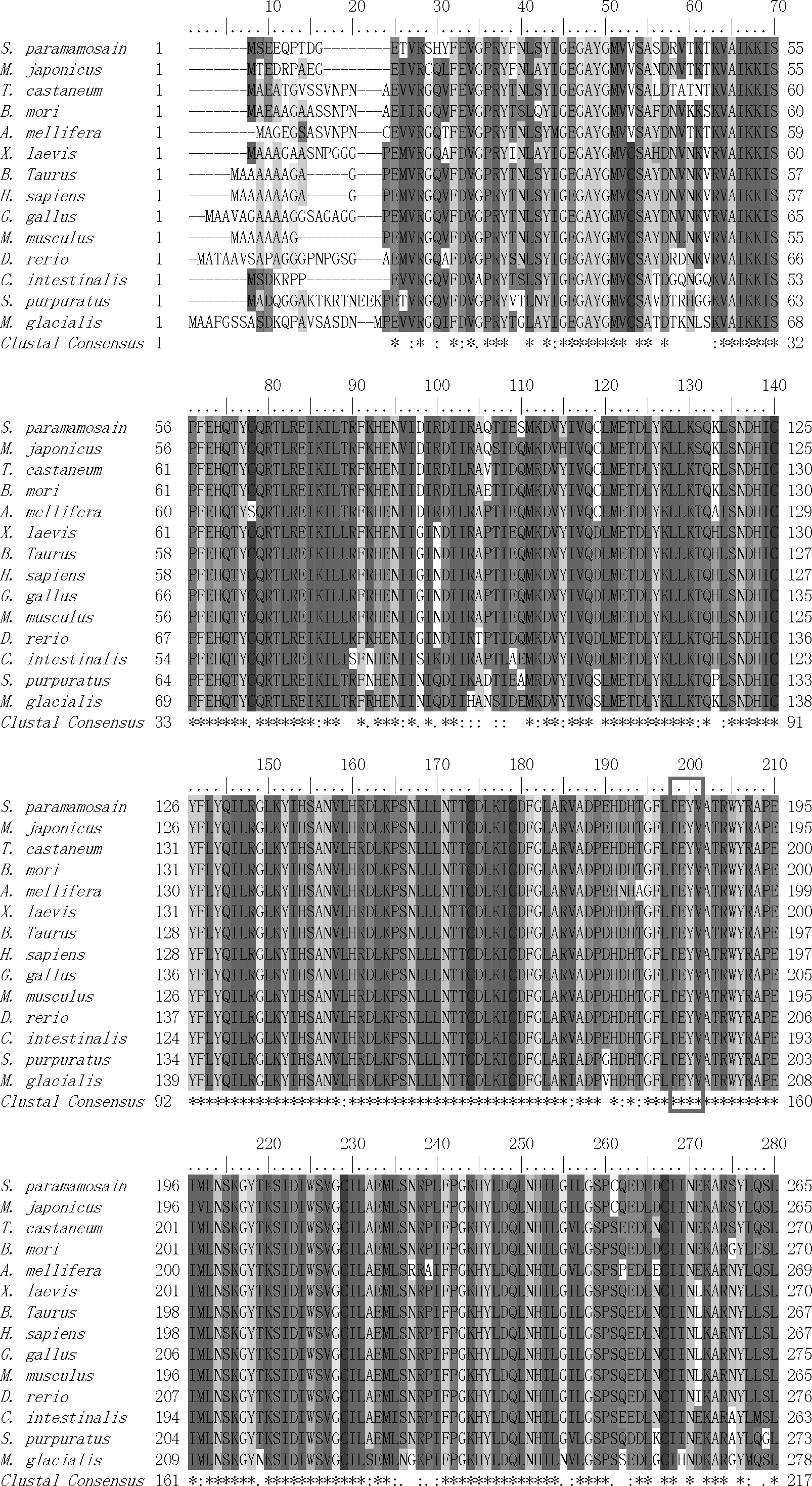
Protein multiple alignments of the ERK2 amino acid sequence between S. paramamosain and other species. The amino acid residues in red frame are the characteristic motif TEY of ERK. For species names see Figure 2.
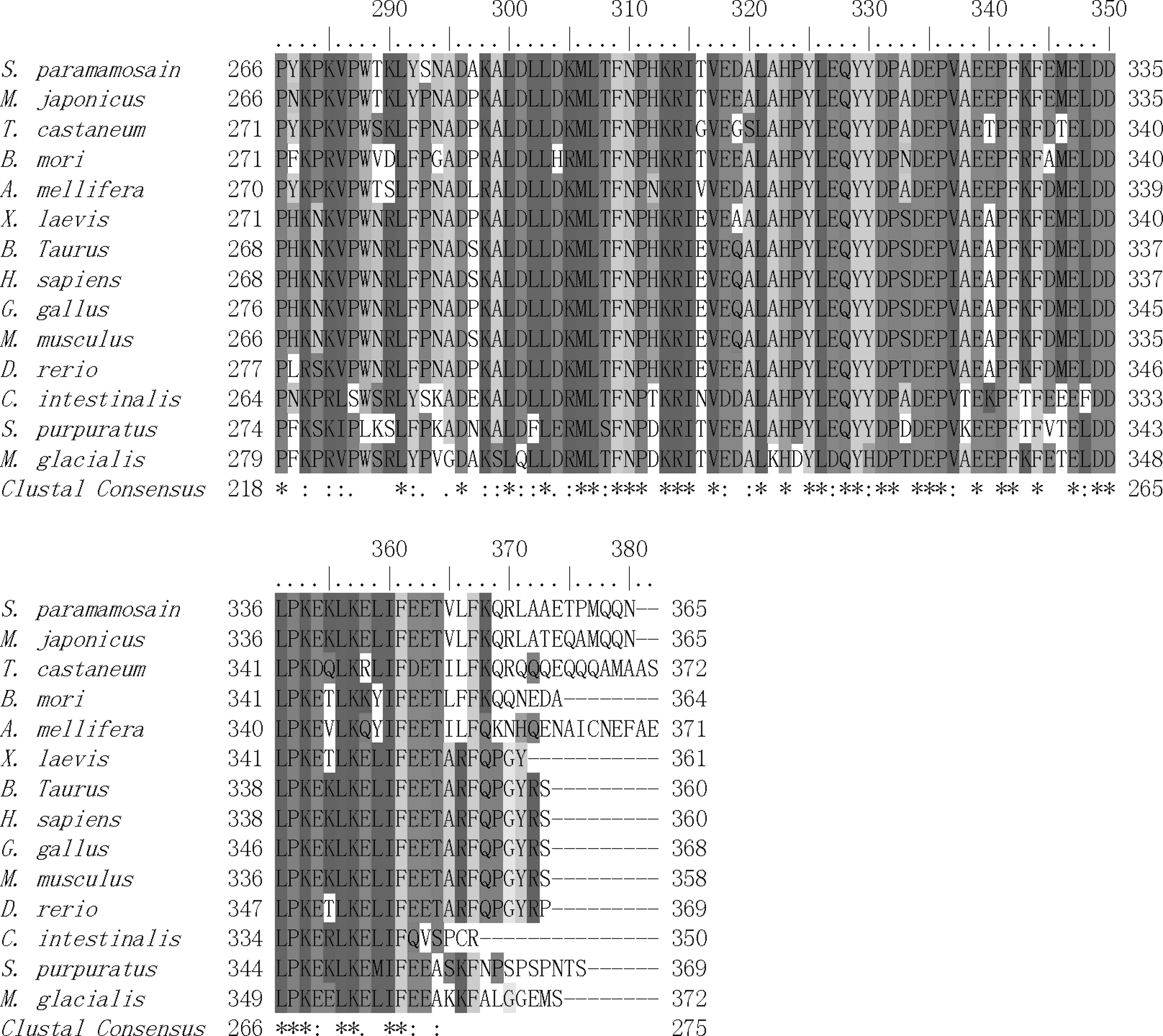
Expression of Sp-erk2 in different tissues
Real-time quantitative PCR results (Fig. 4) showed that Sp-erk2 was expressed in all the tissues chosen. However, compared with heart (H), muscle (M), gill (G), brain (B), stomach (ST), hepatopancreas (HP), and intestines (I), ovary (O) had a significant high expression level (p<0.05), and it led us to further research on the expression levels of different developing stages of ovaries.
Expression of Sp-erk2 in different tissues. G, gill; H, heart; M, muscle; O, ovary; B, brain; ST, stomach; I, intestine; HP, hepatopancreas. Data are presented as means±standard error of the mean (SEM) of at least three separate individuals. Significant differences were indicated: *p<0.05.
Expression of Sp-erk2 in different developing stages of ovaries
Sp-erk2 expression pattern in developing ovaries is also carried out by real-time quantitative PCR. At least five crabs of each stage were used in the experiment. Data collected (Fig. 5) show that the expression level of Sp-erk2 raised gradually as the ovarian stage increased from stage 1 to stage 6 (O1–O6), reaching the highest expression peak at stage 6. Expression level of stage 6 ovary is significantly different from the first four stages (p<0.05), while there is no significant difference between stage 6 and stage 5.

Expression of Sp-erk2 in different stages of developing ovaries. O1, stage 1 (GSI=0.7±0.3); O2, stage 2 (GSI=2.0±0.1); O3, stage 3 (GSI=3.5±0.2); O4, stage 4 (GSI=5.2±0.2); O5, stage 5 (GSI=7.6±0.6); O6, stage 6 (GSI=10.3±0.3). Data are presented as means±SEM of at least three separate individuals. Significant differences were indicated: a and b were significantly different (p<0.05); a/b and ab had no significant differences (p>0.05). GSI, gonad somatic index.
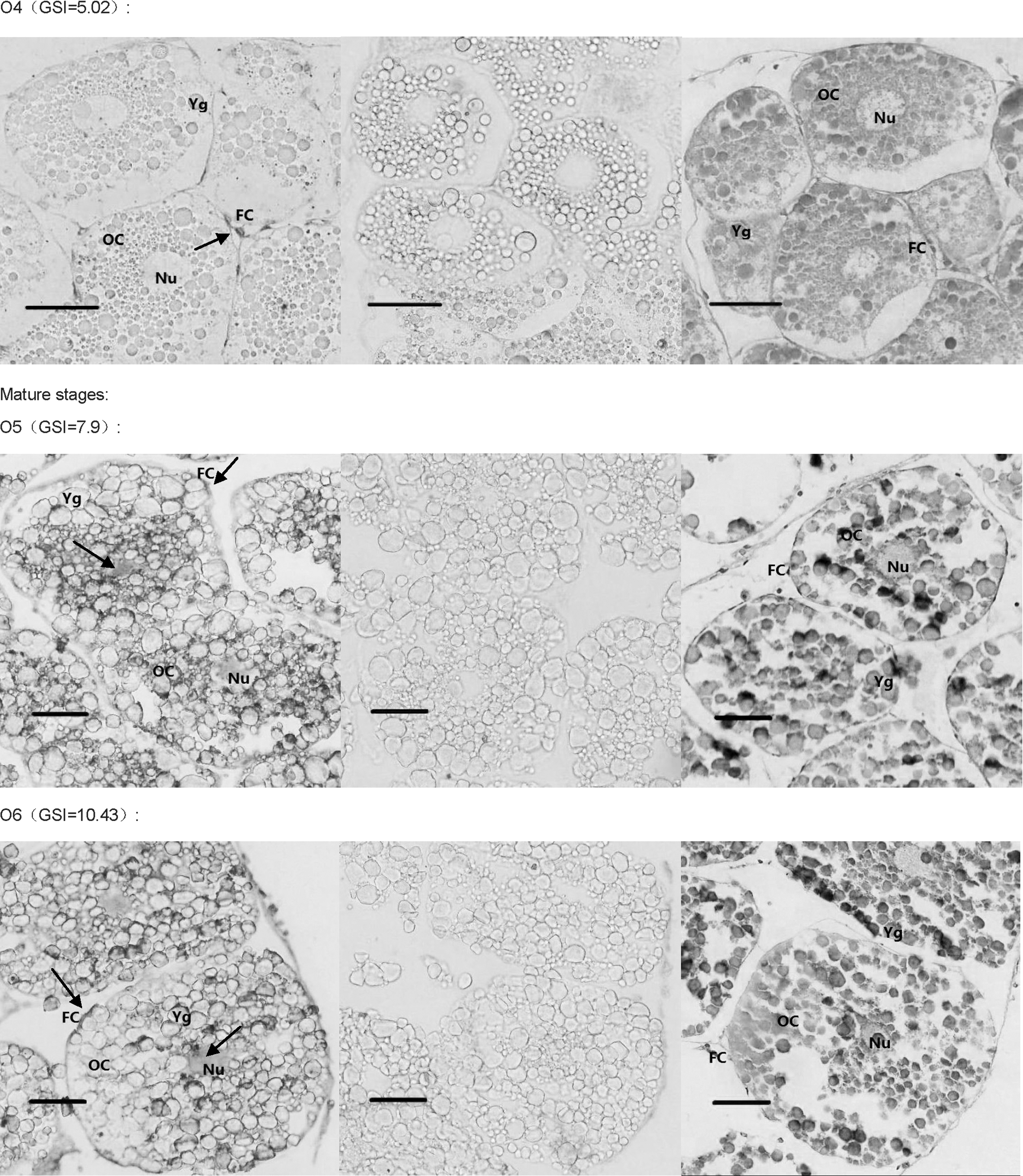
The DIG-labeled RNA probe for Sp-erk2 was obtained using Sp-erk2-ISH-F and Sp-erk2-ISH-R. In situ hybridization histochemistry was performed to clarify and analyze the detail of the expression. Positive signals in early developing ovary were detected in follicular cells around the oocytes (O1–O4), though there seems to have a subtle expression in oocytes in O3 and O4 stages. In relatively mature and fully mature ovarian tissues of GSI 7.9 and 10.43, Sp-erk2 mRNA was distributed in both follicular cells and oocytes (Fig. 6).
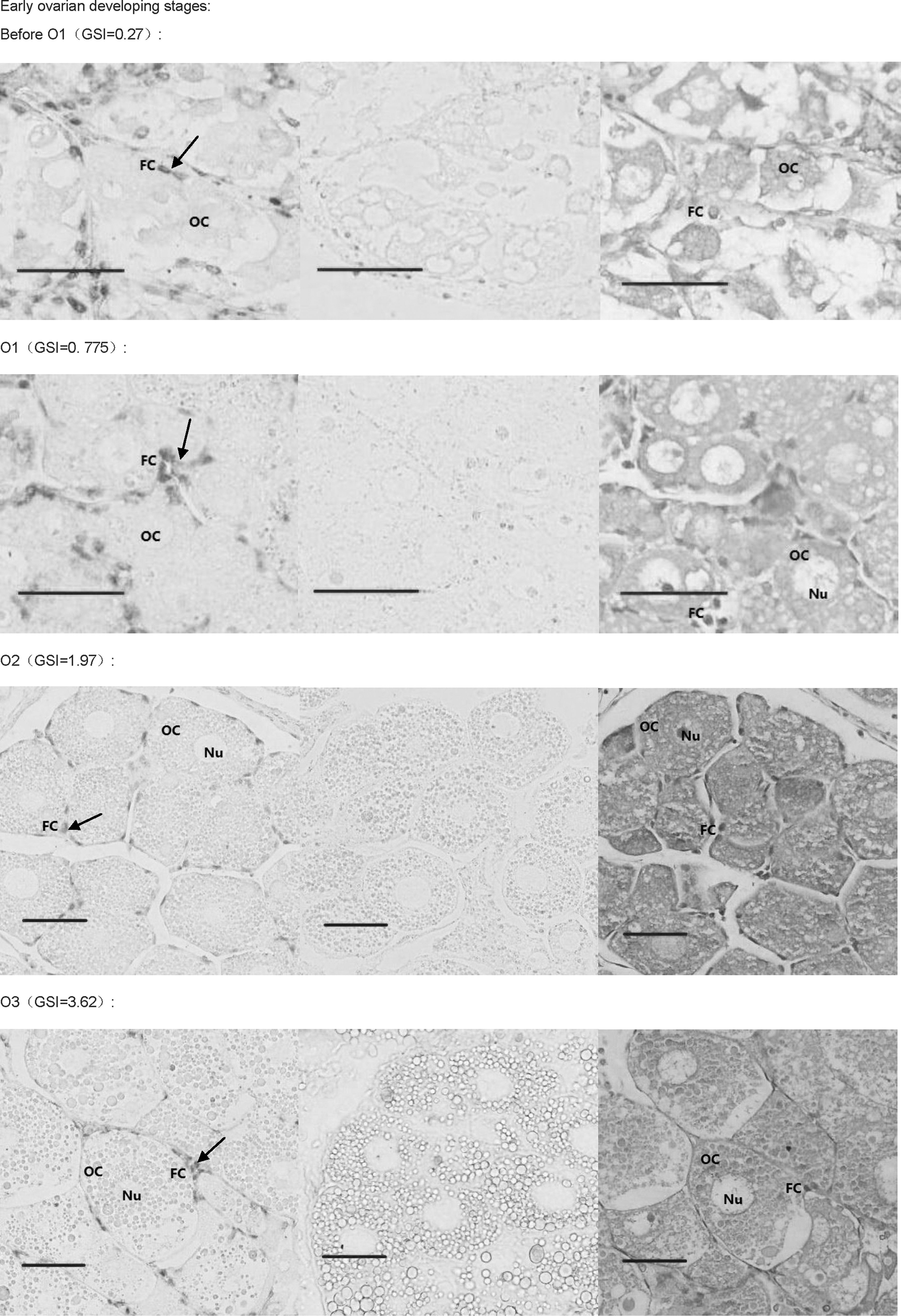
In situ hybridization results. The first picture of each group is the positive result, the second one is negative control, and the third is HE control. Mature stages' group showed that Sp-erk2 was distributed in both follicular cells and oocytes, while in follicular cells only, in early stages' group. The red bars stand for 50 μm. The black arrows indicate the positive signals. OC, oocyte; FC, follicular cell; Nu, nucleus; Yg, yolk globules.
Discussion
MAPK signal transduction pathway is considered to be highly conserved in all the eukaryotes. The key molecules of this pathway show extremely high sequence similarity as evident in the protein multiple alignments. Our results demonstrate that Sp-ERK2 shares 94% identity to the counterpart from M. japonicus, which is the only one identified in crustacean, and has above 80% identity to the other ERKs from almost all the species examined so far. Our data suggest that the structure and function of ERK2 may be somewhat different between marine chordate/echinodermata animals and other species. In addition, ERK signal transduction pathway is famous for the dual-phosphorylation of ERK on threonine and tyrosine separated by a glutamic acid (Widmann et al., 1999), which makes the cascade reaction more effective and precise (Farooq and Zhou, 2004). In Sp-ERK2, the dual-phosphorylation sites, Thr183 and Tyr185, are also found through bioinformatic analysis.
Although erk2 has been investigated extensively in many different species, the expression patterns of this gene in crab ovarian development have not been systematically examined yet. In this study, the real-time quantitative PCR consequences of expression of Sp-erk2 in different green mud crab tissues showed that the expression level in ovaries (relative mature stages) is significantly higher than other tissues. This indicated that Sp-erk2 may play a crucial role in the development and function of crab ovaries. With this hypothesis, we checked the expression pattern in developing ovaries. As expected, the expression levels increase along with the development of ovaries, indicating the function of ERK2 was related to oocyte maturation and ovarian development. Furthermore, the in situ hybridization histochemistry results showed that Sp-erk2 mRNA was detected in crab follicular cells but not in oocytes in early developing stages, while in relatively mature phases positive signals were found in both follicular cells and oocytes, indicating that ERK2 in follicular cells may involve in early steps of oocyte meiosis and ERK2 in oocytes may participate in later events.
Xenopus oocytes were the earliest model for investigation on the interactive mechanism between the ERK pathway and meiotic resumption. In Xenopus oocytes, evidence has revealed that ERK activation is essential for progesterone-induced maturation promoting factor (MPF) activation and germinal vesicle breakdown (GVBD). For instance, ERK activation was shown to occur just before GVBD and was requisite (Posada and Cooper, 1992; Fabian et al., 1993). Microinjection of an anti-Xenopus MEK (ERK kinase) antibody into immature oocytes prevented progesterone-induced ERK activation and GVBD was also inhibited (Kosako et al., 1994; Gotoh and Nishida, 1995). Moreover, microinjection of constitutively activated thiophosphorylated ERK into resting Xenopus oocytes was able to activate MPF and led to GVBD (Haccard et al., 1995). White et al. (2005) found that androgen-mediated maturation and activation of ERK was significantly and specifically reduced by RNA interference of Xenopus androgen receptors. Although all above suggested that activation of ERK in Xenopus oocytes took an important part in meiosis maturation of oocytes, ERK activation could not be detected in mammalian oocytes before meiosis resumption (Liang et al., 2007). More and more reports presented that activation of ERK in follicular cells was probably necessary for meiosis resumption in mammalian oocytes (Zhang et al., 2007; Fan et al., 2009). Studies on mouse and porcine oocytes show that ERK activation in cumulus-enclosed oocytes (CEOs) is initiated before GVBD and is essential for gonadotropin-induced GVBD (Sun et al., 2002; Li et al., 2008; Kolesarova et al., 2010). To observe more clearly, Liang et al. (2005) used ultracentrifugation method to move the opaque lipid droplets aside the porcine oocyte and found that activation of ERK in cumulus cells, but not in oocytes, was essential for GVBD in CEOs, while phosphorylation of ERK in oocytes occurred after GVBD. Similar process was also found in sheep (Cecconi et al., 2008). In our experiments, Sp-erk2 mRNA was found in follicular cells only in early stages of ovarian development, which is similar to the research of mammal cited previously. Thus, we conjectured that it may be related to GVBD in a way similar to that reported in mammals, although further experiments need to be done to prove it.
Gross et al. (2000) found that U0126-treated Xenopus oocytes fail to form metaphase I spindles and appear to enter S-phase instead of M II, indicating that ERK2 activation is required for suppression of S-phase and entry into M II. In mammalian oocytes, it is reported that ERK is indispensable for the post-GVBD events, for instance, spindle organization; M I–M II transition; and M II arrest (Fan and Sun, 2004; Liang et al., 2007). Mature mammalian oocytes are arrested at metaphase of M II and ovulated for fertilization (Liang et al., 2007; Duggavathi and Murphy, 2009). However, there are some differences in several invertebrates such as crab where the fertilization happens at M I arrest when oocytes are ovulated, which means that crab oocytes are ovulated just at M I arrest (Chen et al., 2004). In starfish, which also spawn M I arrest oocytes, M I pause was maintained by ERK as well as low intracellular pH (∼7.0) as recorded by Usui et al. (2008). Moreover, the ERK transduction pathway was reported for involving in M I arrest (Usui et al., 2008), M I–M II transition (Tachibana et al., 2000), pronucleus stage arrest, and the eventual fate decision of starfish oocytes (Kishimoto, 2004; Sadler et al., 2004; Mori et al., 2006). Based on the information just outlined and on our experiments, we deduce that ERK in crab oocytes may participate in events before or around M I since the Sp-erk2 mRNA was strongly distributed in oocytes in relatively mature and mature ovarian tissues of GSI 7.9 and 10.43. To obtain thorough information on the role of ERK signal transduction pathway during crab ovarian development, an ERK knockdown or inhibition experiment should be taken to see whether the development of ovary is blocked or influenced.
To summarize, we have cloned and characterized the Sp-erk2 from S. paramamosain, which is the first such cloning in crustaceans to our knowledge. Our experiments indicated that ERK2 may play a crucial role on crab ovary development. According to our experiments, we concluded that the function of ERK in crab ovarian development is similar to mammalian oocytes rather than to Xenopus oocytes. The results could provide essential information on the regulatory mechanism of crab or even crustacean ovarian development.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 30571430 and 31072200) and Innovation Team foundation of Jimei University (2010A001). The authors thank Mr. Ion Beldorth (Department of Chemistry & Biochemistry, Texas State University, San Marcos, TX) for critical reading of the article.
Disclosure Statement
No competing financial interests exist.