
Abstract
Porphyromonas gingivalis, the key etiologic agent of periodontitis, can be classified into six types (I to V and Ib) based on the fimA genes that encode FimA (a subunit of fimbriae). Accumulated evidence indicates that P. gingivalis expressing Type II fimbriae (Pg-II) is the most frequent isolate from severe periodontitis cases and is more virulent than other types of P. gingivalis. However, during the Pg-II infection process, which specific virulence factors play the key role is still unclear. In this study, we examined the capabilities of three Pg-II strains to invade and modulate the inflammatory cytokine expression of human gingival epithelial cells (GECs) compared to two Pg-I strains. P. gingivalis oligo microarrays were used to compare gene expression profiles of Pg-II strains that invade GECs with Pg-I strains. The differential gene expression of Pg-II was confirmed by quantitative reverse transcription–polymerase chain reaction. Our results showed that all of the Pg-II strains could induce interleukin (IL)-1β and IL-6 secretion significantly when compared to Pg-I strains. Thirty-seven genes that were specifically expressed during the pathogenic process of Pg-II were identified by a microarray assay. These findings provide a new insight at the molecular level to explain the specific pathogenic mechanism of Pg-II strains.
Introduction
The epithelial compartment is thought to provide not only a physical but also an initial immune barrier to prevent mucosal penetration by pathogens. Hence, the structural and functional integrity of gingival epithelial cells (GECs) is essential for periodontal tissue to maintain normal metabolism and homeostasis in the oral cavity, which is considered a gathering place and an incubator of diverse pathogens. P. gingivalis, a highly invasive intracellular pathogen (Lamont et al., 1995), can remain resident in the perinuclear region for prolonged periods without causing cell death (Lamont and Jenkinson, 1998), but can throw off the cells' physiological equilibrium through interference with cell signal transduction pathways (Watanabe et al., 2001; Zhou and Amar, 2007) and cell cyloskeletal rearrangements (Yilmaz et al., 2003). In addition, intracellular P. gingivalis can avoid the host immune defense, spread between host cells to extend its infectious range, and disturb various aspects of cellular function, including the synthesis of proinflammatory cytokines (Andrian et al., 2006; Yilmaz et al., 2006). Therefore, the interaction of GECs and P. gingivalis is an effective model for investigating the potentially virulence-specific factors of Pg-II.
Identifying virulent strains of pathogens, their virulence determinants, and their virulence-specific genes are classic tasks of microbial pathogenesis research. Microarray expression profiling and the development of data-mining tools afford an unprecedented opportunity for the genome-wide study of bacterial pathogenicity (Schoolnik, 2002). To date, based on the genome sequence of P. gingivalis strains that have already been published (Nelson et al., 2003; Naito et al., 2008; Watanabe et al., 2011), various researches have been performed to compare the genome content between different P. gingivalis subspecies (Chen et al., 2004; Yuan et al., 2005) and to analyze the gene expression profile of P. gingivalis during interaction with host cells or other bacteria (Hosogi and Duncan, 2005; Rodrigues and Progulske-Fox, 2005; Simionato et al., 2006). These achievements inspired us to confirm our hypothesis that Pg-II may express some specific genes that are closely associated with its pathogenicity during infection, as virulence-specific gene expression may be modulated during the host–pathogen interaction but not in regular culture conditions (Mahan et al., 1993; Rappuoli, 2000).
Due to the probable influence of variations in fimA and FimA on the pathogenicity of P. gingivalis, in the present study we isolated Pg-II and Pg-I strains from periodontal patients or periodontally healthy adult and compared their adhesion, invasion, and induction of proinflammatory cytokines in epithelial cells. Then, depending on this evaluation, the microarrays were adapted to scan for and identify genes potentially specific to Pg-II strains undertaking invasion compared to the control strain ATCC 33277 (Type I fimbriae strain, designated Pg-I) and another Pg-I strain.
Materials and Methods
P. gingivalis strains and GEC culture
The following P. gingivalis strains were used in this study: ATCC 33277 (Pg-I); WH1103 (Pg-I); WH4404 (Pg-II); WH5601 (Pg-II); and WH115 (Pg-II). ATCC 33277 was kindly provided by State Key Laboratory of Oral Diseases, China; WH1103 was obtained through the subgingival plaque from a periodontal healthy adult; and three Pg-II strains were all separated from the periodontal pockets of patients. The specific method for isolation and identification of clinical isolates was described in our previous studies (Zhao et al., 2007, 2008) (Supplementary Fig. S1; Supplementary Data are available online at
Healthy human gingival tissue samples were obtained with informed consent from periodontal healthy adults undergoing third-molar extraction at the Oral Surgery Clinic of the West China Hospital of Stomatology, Sichuan University. The cells were prepared for culture as previously described (Krisanaprakornkit et al., 1998). The authenticity of GECs was confirmed histologically by the cell morphology and by immunohistochemistry with a monoclonal antibody against human pankeratin. The isolated GECs were cultured in serum-free keratinocyte medium (K-SFM) supplemented with keratinocyte growth factors, hydrocortisone, bovine insulin, bovine pituitary extract, gentamicin sulfate, amphotericinB, and 0.15 mM CaCl2. The cells were grown to 80% confluence before the treatment with bacteria.
Adhesion and invasion assay
The adhesion to and invasion of GECs by P. gingivalis were quantified using the colony-forming assay and antibiotic protection assay as described by Nakagawa with slight modifications (Nakagawa et al., 2006). P. gingivalis were grown to the mid-log phase, then were harvested by centrifugation at 10,000 g for 10 min, and suspended in K-SFM at 108 colony-forming units (CFU)/mL as determined using a Petroff-Hausser counting chamber. The bacterial suspensions were added to monolayer of GECs (106/well) in six-well culture plates at an MOI of 100 and incubated for 90 min at 37°C in 5% CO2. External nonadherent bacteria were removed by washing the cells three times with phosphate-buffered saline (PBS), and then the cells were disrupted by the addition of distilled water and an incubation at 37°C for 20 min. Serial dilutions of the disrupted mixture were plated on blood agar plates and incubated for 7 days, and the numbers of adherent organisms were determined. To determine the numbers of invasive bacteria, P. gingivalis–infected cells were incubated with a medium containing gentamicin (0.3 mg/mL) and metronidazole (0.2 mg/mL) for 1 h to kill the extracellular bacteria. Control wells were included to verify the effect of antibiotic treatment to kill the extracellular bacteria (Supplementary Fig. S5–6). The cells were washed three times with PBS, and the numbers of internalized bacteria were determined as described above. Adhesion and invasion efficiencies were calculated using the formula: adherent/invasive rate=(the number of colonies on the plate×dilution multiple)/initial inoculums×100%. All of the assays were performed in triplicate. The three multiple wells were used for the statistical analysis of each experiment of the strains.
Cytokine induction assays
The bacterial suspensions with 108 CFU/mL were added to monolayer of GECs with 106 cells/well in six-well culture plates (MOI: 100). After 6 or 24 h incubation at 37°C in 5% CO2, cell-free supernatants were harvested and stored at −80°C until used. The concentrations of interleukin (IL)-1β and IL-6 were measured by commercial enzyme-linked immunosorbent assay (ELISA) kit (BD Opt EIA; BD Biosciences) according to the manufacturer's protocols. The absorbance at 450 nm was read using a microplate reader with wavelength correction set at 550 nm.
Bacterial genomic RNA preparation
P. gingivalis strains were cultivated anaerobically in BHI medium or co-cultured with GECs (MOI: 100) for 18 h as mentioned previously. The co-culture time of 18 h was chosen in accordance with a fluorescence microscopy assay, which demonstrated that the vast majority of P. gingivalis cell could internalize at this time (Belton et al., 1999). After 18 h co-culture, cells were washed three times to remove external organisms and lysed with distilled water. The bacterial cells were harvested and treated with 0.5 mg/mL DNase I and 0.25 mg/mL RNase A in PBS supplemented with 1 mM MgCl2 to remove the epithelial cell DNA and RNA before the total RNA isolation. The total RNA was isolated from P. gingivalis by a Totally RNA kit (Ambion) as described by the manufacturer and stored at −80°C after quantitation (Supplementary Fig. S2).
Microarray analysis
Microarray analysis was performed using a Roche Nimblegen custom-made P. gingivalis W83 whole-genome oligonucleotide chip of 135k (Roche Nimblegen) representing 1907 genes. The polyadenylation-involved oligo-dT priming amplification method was used to deplete samples of ribosomal RNAs and prepare the target cDNA. Then, the labeled cDNA was purified, mixed, and hybridized on the microarrays. The procedure has been previously described (Cao et al., 2010).
After the hybridization, the microarrays were scanned with a confocal scanner LuxScan™, and the images obtained were then analyzed using LuxScan 3.0 software (CapitalBio). Faint spots were removed from the data if the intensities were lower than the average intensity plus 2 standard deviations of the negative controls, and the data were normalized using a LOWESS intensity-based normalization. To improve the reliability for each sample, the amplifications and hybridizations were performed using the dye-swap strategy. To improve the reliability of gene expression during the invasion process, two biological replicates for each P. gingivalis strain recovered from the GECs were used in microarray analysis, the generated files were imported into Microsoft Excel (Micorsoft Corporation) for subsequent analyses, and the data were expressed as the gene expression ratio. Relative fold changes in gene expression were calculated as the intensity of Cy3/Cy5 or Cy5/Cy3 (Cao et al., 2010). The same magnitude of change was used in this array for the up-regulated and down-regulated genes, and genes were identified as differentially expressed if there was a 2.0-fold difference in their average expression values.
Validation of microarray results using real-time (reverse transcription)–polymerase chain reaction
Quantitative reverse transcription–polymerase chain reaction (RT-PCR) was performed to confirm the differential gene expression observed in the microarrays of the Pg-II strains, including seven genes with higher expression and one gene with a lower expression. Sequences of primer pairs for these genes were acquired through links provides by the GenomeViewer software. The primer sequences were identical to those used by TIGR and are listed in Table 1. RT-PCR was performed in 7900 HT Fast Real-Time PCR system (Applied Biosystems) in 20 μL reaction volumes that contained 10 μL of Power SYBR Green PCR Master Mix (Applied Biosystems), 0.5 μL of each gene-specific primer, 1 μL of cDNA template, and 8 μL of nuclease-Free Water. The cycling conditions consisted of initial 10-min enzyme activation at 95°C, followed by 40 cycles of denaturation at 95°C for 15 s, annealing at 60°C for 1 min, and elongation at 60°C for 1 min. All reactions were repeated three times and melting curve analysis indicated that in each reaction a single product was amplified (Supplementary Fig. S3).
Locus number from TIGR (
F, forward; R, reverse.
Statistical analysis
Statistical analysis was carried out by SPSS 16.0 software (SPSS Inc.) using an independent sample t-test. The significance level was set at p<0.05.
Results
Adherent and invasive abilities of Pg-I and Pg-II strains
The data for adhesion to and invasion of GECs by Pg-I and Pg-II strains are shown in Table 2. According to the standard in Dorn's research (Dorn et al., 2000), although one Pg-II strain exhibited high invasive efficiency rate (1.258%±0.157%), no statistical difference was noted between Pg-I and Pg-II strains regarding degree of adherent/invasive rates (p>0.05). These results suggest that Pg-II strains used in the present study did not show significantly higher adherent and invasive abilities compared to Pg-I strains as a whole.
Values represent means±standard deviations of triplicate experiments.
Invasive rate: >1% indicates high rate; 0.1%–1% indicates moderate rate; <0.1% indicates low rate.
Effect of Pg-I and Pg-II strains on IL-1β and IL-6 production by GECs
As shown in Figure 1, all of the five P. gingivalis strains could stimulate GECs to express IL-1β and IL-6 compared with the control group at both 6 and 24 h (p<0.01 or p<0.05). Notably, co-culturing GECs with Pg-II strains resulted in a significantly increased level of IL-1β and IL-6 compared to cells with Pg-I strains at both time points (p<0.01). It is obvious that the stimulatory effect of P. gingivalis on cytokines production in GECs increased with the extended incubation time from 6 to 24 h.
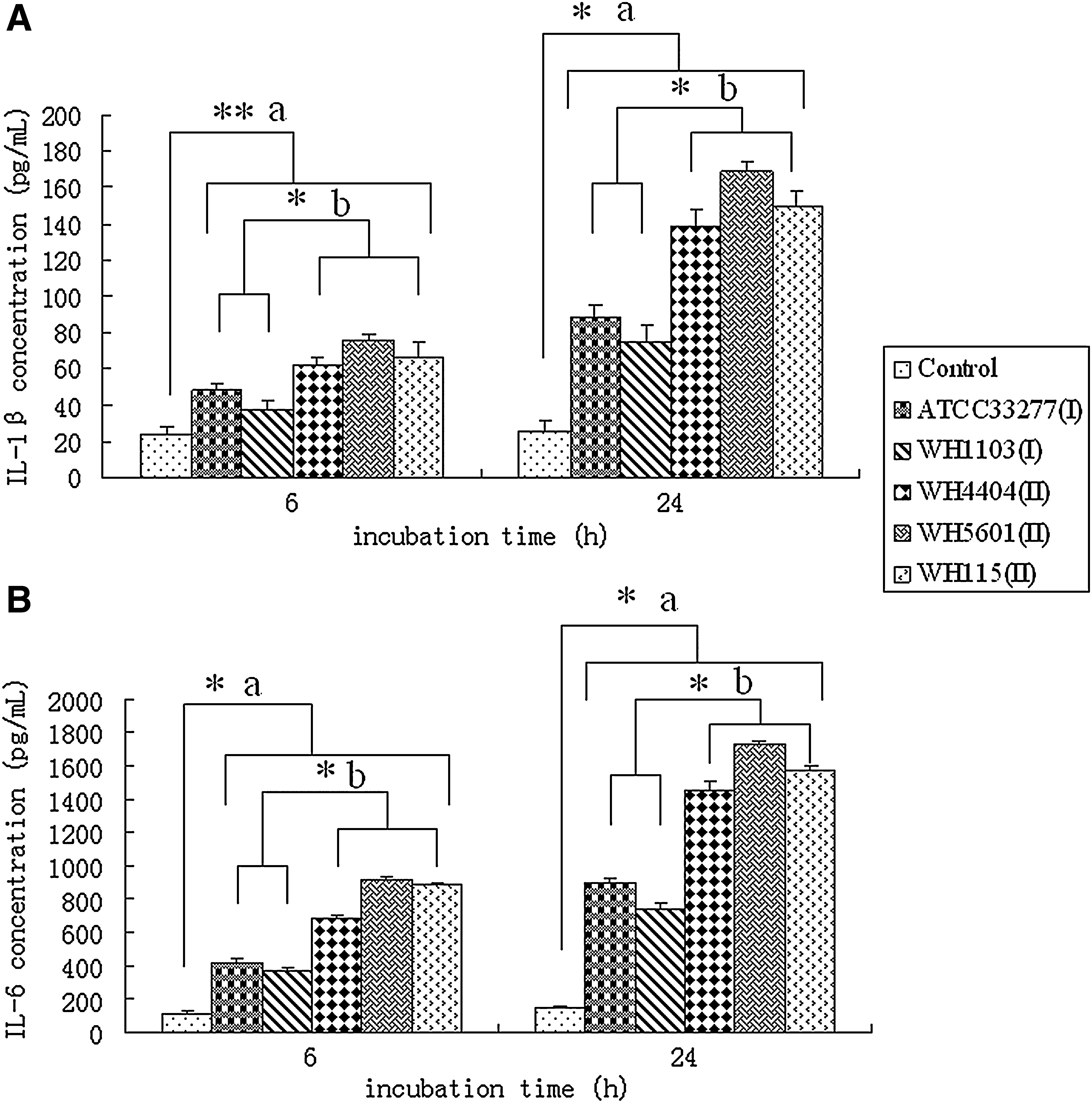
IL-1β
Genomotyping analysis by microarray assays
So far, the reported whole genomic sequences of P. gingivalis are only available for strain W83 [GenBank: AE015924], ATCC 33277 [GenBank: AP009380] and a clinical isolate TDC 60 [GenBank: AP012203] (Nelson et al., 2003; Naito et al., 2008; Watanabe et al., 2011). W83, a commonly accepted virulent strain compared to the avirulent ATCC 33277, is the reference strain of P. gingivalis that has been widely used for the characterization of bacterial features. One global comparative analysis confirmed that 7% of predicted genes showed very low or no signals in ATCC 33277 in spite of the 93% chromosomal similarity between W83 and ATCC 33277 (Chen et al., 2004). This result suggests that some pathogenic genes may be absent from ATCC 33277, and W83 may possess more virulent genes. Moreover, ATCC 33277 was assessed as an avirulent control, along with another Pg-I clinical isolate. Thus, it is more objective and reliable to design the chip in accordance with W83 ORFs.
Microarray-based competitive hybridizations with labeled cDNAs from two Pg-I stains or three Pg-II isolates were performed for a total of 15 slides. Three slides were utilized for each strain to screen genes that could be regulated during the invasion of GECs: one for P. gingivalis cultivated in vitro and two for cells recovered from GECs. Fold changes in gene expression between GEC-invasive bacteria and bacteria cultures alone were calculated as mentioned in the Materials and Methods section. The genes that were differentially expressed in all of the three Pg-II strains were compared to the differentially expressed genes in the two Pg-I strains, and the specific genes of the Pg-II strains within GECs were acquired.
Analysis of the microarray images from the two Pg-I strains showed that a total of 483 common genes were differentially regulated. Of these genes, 249 were up-regulated and 234 were down-regulated after internalization of GECs, compared with those growing in broth culture. At the same time, 408 genes that were differentially expressed in all of the three Pg-II strains were obtained, which included 215 up-regulated genes and 193 down-regulated genes. According to the comparison of commonly exchanged genes from Pg-I and Pg-II strains, 37 genes (19 up-regulated and 18 down-regulated genes) were selected as potentially Pg-II-specific genes and are shown in Tables 3 and 4.
Locus number and putative identification are according to the TIGR Porphyromonas gingivalis genome database.
Relative fold indicates expression of that gene of Pg-II within GECs versus in BHI; it is the average of three strains of Pg-II.
GEC, gingival epithelial cell; BHI, brain heart infusion.
Locus number and putative identification are according to the TIGR P. gingivalis genome database.
Relative fold indicates expression of that gene of Pg-II in BHI versus within GECs; it is the average of three strains of Pg-II.
Quantitative RT-PCR validation of differentially expressed genes
Seven of the up-regulated genes (PG0001, PG0625, PG0758, PG1129, PG1175, PG1676, and PG1858) and one down-regulated gene (PG0444) of the Pg-II strains were random selected to confirm the accuracy of microarray data by RT-PCR. The same RNA samples were used for RT-PCR as well as microarray assay. As shown in Figure 2, all of the tested genes demonstrated an alteration in consistent with the microarray.

Quantitative RT-PCR analysis was performed to confirm the microarray data. The seven genes (PG0001, PG0625, PG0758, PG1129, PG1175, PG1676, and PG1858) from the up-regulated group and one gene (PG0444) from down-regulated group demonstrated an alteration in consistent with microarray. The quantitative RT-PCR data represent as the means±standard deviations of triplicate determinations. RT-PCR, reverse transcription–polymerase chain reaction.
Discussion
Our previous epidemiological survey demonstrated that a majority of periodontal patients were found to carry Pg-II, while Pg-I was widely distributed among the periodontally healthy people. Moreover, Pg-II isolates were more frequently detected in deep pockets and co-existed with Aggregatibacter actinomycetemcomitans and Tannerella forsythia (Zhao et al., 2007). Thus, we speculated that Pg-II organisms may be disease-associated strains that are closely associated with the initiation and progression of chronic periodontitis in Chinese adults. In the current study, we isolated Pg-II and Pg-I strains from periodontal patients or periodontally healthy adult and compared their ability to invade GECs and modulate inflammatory cytokine expression. The results showed that Pg-II strains did not have significantly higher adherent and invasive abilities compared to Pg-I strains. These results are similar to the data obtained by Umeda (Umeda et al., 2006) and Inaba (Inaba et al., 2008), which showed that fimA genotypes were not related to the adherent and invasive abilities of P. gingivalis strains, and invasive efficiency may be related to the activities of secreted gingipains. However, it should be noted that adherent/invasive ability is not the only determinant correlated with bacterial virulence. It is well known that strain W83 exhibits a much lower invasive ability compared with ATCC 33277 (Pg-I), but it is considered more virulent because of genes involved in capsular polysaccharide synthesis and other ORFs obtained by horizontal gene transfer (Chen et al., 2004). Our finding that all of the Pg-II strains could stimulate GECs to express high levels of IL-1β and IL-6 also supports this view. In another assay of macrophage activation, Pg-II strain OMZ314 and purified type II fimbriae from this strain were more potent than Pg-I in inducing proinflammatory cytokine (Wang et al., 2009). Furthermore, through measuring the serum sialic acid concentration and other inflammatory parameters in a mouse abscess model, studies found that most Pg-II strains could cause severe inflammation compared to ATCC 33277 (Nakano et al., 2004; Inaba et al., 2008). These lines of evidence indicate that there must be some potential virulence strategies that are responsible for the increased prevalence and high virulence of Pg-II during its interaction with the mammalian host.
To test this hypothesis, mircoarray-based comparative transcriptome profiling has been applied to compare the gene modulation of five P. gingivalis strains in the invasion process and identify the specific genes that encoded the potential virulence factors of the Pg-II strains. The results showed that 370 genes were commonly modulated in all of the five P. gingivalis strains within GECs: 196 up-regulated and 174 down-regulated genes (data not shown) (Supplementary Table S1). The up-regulation of many genes associated with bacteria adhesion, invasion, evasion, the stress response, and the acquisition of peptides have already been identified and characterized in previous studies (Nelson et al., 2003; Park et al., 2004). The comparison of our microarray results with the differential display RT-PCR analysis of altered genes that are differentially regulated in the invasion process of P. gingivalis revealed that almost all up-regulated genes identified in this research were included in the up-regulated genes observed in our study, such as PG0159 (endopeptidase PepO), PG0165 (Heat shock protein 15), and PG1642 (cation-transporting ATPase) (Park et al., 2004). However, there are few overlaps between our result and other microarray assays that were used to identify genes of P. gingivalis differentially expressed during interactions with Hep-2 cells or human endothelial cells (Hosogi and Duncan, 2005; Rodrigues and Progulske-Fox, 2005). Different host cells and the co-culture time used in the research may cause the observed differences between our assay and others, indicating that host cell activities play a major role in influencing the gene expression of pathogens. Significantly, of these few overlapped genes, some may be involved in the attachment and coaggregation of P. gingivalis with epithelial cell membranes, such as PG1683, which encodes a conserved hypothetical protein that has homology to α-amylases; some are correlated with pathogenesis or protein stabilization, such as PG0506 (Arg-gingipain B gene, rgpB) and PG0593 (Heat-induced serine protease, htrA). These consistent results may provide some useful information for studying the potential pathogenicity of P. gingivalis during contact with host cells.
Furthermore, according to mircoarray assay profiling, 37 specifically expressed genes of the Pg-II strains were further identified, including 19 up-regulated genes. These genes have been suggested to be involved in the biosynthesis of cofactors, prosthetic groups, and carriers; cell envelope; DNA metabolism; energy metabolism; protein fate; protein synthesis; and purines, pyrimidines, nucleosides, and nucleotides. There are also genes with unknown functions and those encoding hypothetical proteins in this group. The specific functions of these genes will be discussed in detail subsequently.
Of these genes, there were two up-regulated genes involved in the biosynthesis of cofactors: PG0625 (folE) encoding GTP cyclohydrolase I, which catalyzes the formation of folic acid from GTP in the first step, and PG0211 (cbiGF) encoding the cobalamin biosynthesis protein CbiG, which regulates cobalamin synthesis. In many types of bacteria, folic acid can promote the proliferation of microorganisms and therefore play a role in pathogenesis. It has also been shown that a perturbation of the bacterial folate-mediated one-carbon metabolism can reduce the growth and virulence of Staphylococcus aureus (Kao et al., 2010). Cobalamin is required for the degradation of mainly amino acids and peptides as energy sources in P. gingivalis and other bacteria (Roper et al., 2000; Huang et al., 2007). Therefore, the up-regulation of PG0625 and PG0211 may suggest an increased utilization of folic acid and cobalamin in the pathogenesis process of Pg-II strains.
The cell envelope structures, such as lipopolysaccharides (LPS), peptidoglycan, surface polysaccharides, and murein sacculus, are all important virulence factors of P. gingivalis. A total of 18 genes involved in cell envelope were up-regulated in the process of the invasion of epithelial cells and 3 were specifically expressed in Pg-II strains. PG0578 (murD) encoding UDP-N-acetylmuramoylalanine-D-glutamate, which catalyzes the attachment of D-glutamate to a cytoplasmic peptidoglycan precursor, is involved in cell wall and peptidoglycan biosynthesis. Properly constructed peptidoglycan networks in the bacteria cell wall are necessary for bacteria metabolism and protect bacteria against high internal osmotic pressure and sometimes harsh external environment (Walsh et al., 1999). It has also been found that peptidoglycans purified from P. gingivalis can induce cell death, stimulate the production of IL-6 from mouse peritoneal macrophages, and act as an immune stimulant to cause inflammation in the host, which may account for the high IL-6 induction by the Pg-II strains in our experiments (Ishii et al., 2010). The important role of LPS in the pathogenic process of P. gingivalis has been well recognized. PG2223 encodes a glycosyltransferase that is involved in the synthesis of O-antigen side chains of the LPS component of a gram-negative outer membrane (Chen et al., 2000). The knockout of a gene that is homologous with genes encoding glycosyltransferase in P. gingivalis resulted in a complete loss of surface-associated gingipain proteinases, which is also a major virulence factor of P. gingivalis. Another up-regulated gene, PG1711, encodes α-1, 2-mannosidase, which plays an important role in protein glycosylation and pathogen–host interaction (Zhou et al., 2009). In addition, PG2223 and PG1711 can both help in the evasion of P. gingivalis from the host immune response.
PG0001 encodes a chromosomal replication initiation protein that plays an important role in the initiation and regulation of chromosomal replication and can affect the transcription of multiple genes, including itself. Therefore, the up-regulation of PG0001 suggests an enhanced DNA replication in the pathogenic process of Pg-II strains, and PG0001 may participate in pathogenesis by regulating DNA replication.
Recently, the bacterial metabolism during infection has been appreciated to contribute to persistence as much as their virulence properties. In our study, the energy metabolism-associated genes PG1858 and PG1676 (pckA) were highly up-regulated with an average change of 10.536-fold and 4.416-fold, respectively, indicating the role of genes in the physiological adaptation of Pg-II strains to an intracellular environment. Flavodoxin (PG1858) is an electron-transfer protein involved in a variety of reactions and metabolic routes (Sancho, 2006). This protein is essential for the survival of some human pathogens and is being investigated as potential drug target for the treatment of bacterial infections, such as infections by Helicobacter pylori (Cremades et al., 2005). A genome analysis of Helicobacter suis (H. suis) also found that flavodoxin and its complex may play a role in the gastric colonization by H. suis and the MALT lymphoma development in H. suis–infected people (Vermoote et al., 2011). The high up-regulation of PG1858 indicates that flavodoxin may participate in the pathogenesis of Pg-II strains and provides a new perspective for drug therapy of periodontitis. PG1676 encodes phosphoenolpyruvate carboxykinase (PEPCK), which catalyzes the first committed step in gluconeogenesis. Gluconeogenesis is required for virulence in microbes with an array of pathogenic lifestyles (Liu et al., 2003; Naderer et al., 2006). A defect in gluconeogenesis has been shown in many organisms to result in a significant reduction in the fitness within the host, deceased plaque formation, and a defect in the intracellular growth or cell-to-cell spread (Gore and Payne, 2010). A study of Mycobacterium tuberculosis indicated that PEPCK is essential for the growth of bacteria on fatty acids, and it catalyzes carbon flow from tricarboxylic acid cycle-derived metabolites to gluconeogenic intermediates, which is required for bacterial growth. The bacteria lacking PEPCK not only failed to replicate in mouse lungs but also failed to survive, and a PEPCK depletion resulted in mycobacterial clearance during the chronic phase of infection and a lower bacterial population in IFN-γ-deficient mice; as a result, this enzyme represents an attractive target for chemotherapy (Marrero et al., 2010). However, there are few studies on the gluconeogenesis and PEPCK levels in P. gingivalis, and the results of our study suggest that research on glucose metabolism may shed light on the specific virulence of Pg-II strains and provide a new chemotherapy target for periodontitis.
Several genes involved in protein fate and synthesis were identified as being associated with Pg-II strain virulence. PG2000 encodes a signal peptidase-related protein, which participates in the protein export pathway and plays a role in the invasion of bacteria. Both PG0758 (dcp-1) and PG1789 (dcp-2) encode peptidyl-dipeptidase and have been identified as virulence agents and are involved in the acquisition of peptides in P. gingivalis W83 (Nelson et al., 2003). PG1004 has also been identified as a virulence agent in P. gingivalis W83, and it encodes a type of serine endopeptidase that can mediate invasion by triggering an increase in Ca2+ concentration (Coetzer et al., 2008). All of these genes belong to the peptidase family, and participate in the biological process of proteolysis. Proteolytic enzymes have been implicated in the invasion, tissue destruction, and evasion of host antibacterial defenses by P. gingivalis, and they not only can degrade various host peptides and cause inflammation, but also can provide bacteria with nutrients in the form of small peptides and amino acids (Potempa et al., 2000), indicating their critical role in the virulence of P. gingivalis. PG0969 encodes S-adenosylmethionine, which is a tRNA ribosyltransferase-isomerase involved in tRNA modification and tRNA-queuosine biosynthesis. This factor participates in protein synthesis, and its role in the pathogenic process of Pg-II strains is not clear.
PG1129 encodes ribonucleotide reductase (RNR), which provides the precursors necessary for DNA synthesis and catalyzes the biosynthesis of deoxyribonucleotides from the corresponding ribonucleotides. RNRs have been identified as virulence determinants in several organisms (Kirdis et al., 2007; Gill et al., 2010); they can help the bacteria to grow anaerobically and ensure the normal replication and growth of pathogens. Deletion of some types of RNRs leads to a reduced virulence or even avirulence of the pathogen, and the study of RNRs may provide a new direction for the antibacterial treatment of periodontitis.
ABC transporters constitute one of the largest and most highly conserved superfamilies and are found in large numbers in all organisms. The important role of ABC transporters in cell-to-surface and cell-to-cell interactions and biofilm development has been reported in several bacteria (Matthysse et al., 1996; Hinsa et al., 2003). In the genome of P. gingivalis W83, a total of 39 different ABC transporter genes have been identified. It has been shown that ABC transporters are required for an optimal entry of P. gingivalis into epithelial cells (Park et al., 2004) and are involved in the LPS biosynthesis and biofilm formation of P. gingivalis (Toyoda et al., 2010). In this study, two ABC transporters (PG0685 and PG1176) were up-regulated in both the Pg-I and Pg-II strains, while PG1175, the inner membrane component of an ABC transporter complex with an N-terminal transmembrane domain and a C-terminal ABC ATPase domain, was specifically up-regulated in the Pg-II strains. PG1175 has been detected to be up-regulated in a monospecific biofilm of P. gingivalis W50 (Lo et al., 2009), suggesting that this gene may contribute to the high virulence of Pg-II strains by promoting biofilm formation or by other possible mechanisms. However, several ABC transporter genes were down-regulated in both the Pg-I and Pg-II strains (PG0682, PG1663, PG1664, and PG1665), and the role of these genes in the interaction of P. gingivalis and epithelial cells remains to be determined.
Some of the up-regulated genes in the Pg-II strains found inside of cells were of unknown or poorly characterized function or encode hypothetical proteins. PG1553 encodes a CobN/magnesium chelatase family protein, with an average change of 16.407-fold. A comparison between P. gingivalis with high virulence and low virulence showed that a putative CobN/magnesium chelatase was involved in heme uptake and might be a relevant virulence determinant of P. gingivalis (Brunner et al., 2010). PG0980, PG1649, and PG1179 encode hypothetical proteins, and PG1179 was highly up-regulated with an average change of 9.514-fold. These genes and their products may be the potential virulence factors of Pg-II strains and their functions require further study.
In contrast to genes up-regulated, 18 genes were down-regulated in the Pg-II strains during the invasion process. Among these genes, seven genes encode hypothetical proteins, and some were significantly down-regulated. The down-regulation of three genes (PG0446, PG2109, and PG2110) involved in the thiamine biosynthetic process may indicate a decrease utilization of thiamine in the pathogenesis of Pg-II strains. PG0194 and PG0293 both contribute to the invasion of P. gingivalis, and PG0443 plays a role in the adhesion. The down-regulation of these genes may account for the moderate adherent and invasive abilities of Pg-II strains compared to Pg-I strains. PG0444 and PG1606 are involved in transport and binding proteins, and PG1606 participates in voltage-gated potassium channel activity, suggesting a change in potassium ion flow as a part of pathogenesis of Pg-II strains. PG0396 encodes a Crp/FNR family transcriptional regulator, which can regulate the transcription of many genes and is a regulator of the virulence of bacteria (Akif et al., 2006). The down-regulation of this gene suggests that it may be a negative regulator of the virulence of Pg-II strains.
This is the first report of a global genomic expression profile that identifies the specific genetic differences of Pg-II during invasion of GEC cultures. In the experiments reported here, 37 genes specifically expressed during the pathogenic process of Pg-II stains were identified, and the potential functions of these genes were further analyzed in detail. Although these specific genes from Pg-II are only beginning to be understood, and their mechanisms remain to be elucidated, the results presented here may provide new insights at the molecular level to explain the high virulence of Pg-II and may also help to better understand the pathogenic mechanism of P. gingivalis, which may lead a new treatment for periodontitis.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (30801295) and (30872872) and Science Foundation for The Excellent Youth Scholars of Ministry of Education of China (200806101107).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.