
Abstract
The myocyte enhancer factor 2A (MEF2A) gene encodes a member of the myocyte enhancer factor 2 (MEF2) protein family that is involved in vertebrate skeletal, cardiac, and smooth muscle development and differentiation during myogenesis. According to recent studies, MEF2 genes might be major regulators of postnatal skeletal muscle growth; thus, they are considered to be important, novel candidates for muscle development and body growth in farm animals. The aim of the present study was to search for polymorphisms in the bovine MEF2A gene and analyze their effect on the MEF2A mRNA expression level in the longissimus dorsi muscle of Polish Holstein-Fresian cattle. In total, 4094 bp of the whole coding sequence and the promoter region of MEF2A were re-sequenced in 30 animals, resulting in the detection of 6 novel variants as well as one previously reported SNP. Three linked mutations in the promoter region (-780T/G, g.-768T/G, and g.-222A/G) and only two genotypes were identified in two Polish breeds (TTA/TTA and TTA/GGG). Three SNPs in the coding region [g.1599G/A (421aa), g.1626G/A (429aa), and g.1641G/A (434aa)] appeared to be silent substitutions and segregated as two intragene haplotypes: GGG and AAA. Expression analysis showed that the mutations in the promoter region are highly associated with the MEF2A mRNA level in the longissimus dorsi muscle of bulls carrying two different genotypes. The higher MEF2A mRNA level was estimated in the muscle of bulls carrying the TTA/TTA (p<0.01) genotype as compared with those with TTA/GGG. The results obtained suggest that the nucleotide sequence mutation in MEF2A might be useful marker for body growth traits in cattle.
Introduction
In the present study, genetic variations in the promoter and coding regions of the MEF2A gene were investigated in 200 bulls representing four cattle breeds using multitemperature single strand chain polymorphism (MSSCP) and DNA sequencing methods. In addition, the promoter variant effect on MEF2A mRNA expression was evaluated in the longissimus dorsi muscle of Polish Holstein-Fresian (HF) bulls.
Materials and Methods
Blood, tissue sampling, DNA and RNA isolation, and cDNA preparation
The blood samples were obtained from 200 unrelated bulls of four cattle breeds including Polish HF (n=130), Polish Red (PR, n=30), Hereford (HER, n=20), and Limousine (LIM, n=20). DNA was extracted from blood using Wizard® Genomic DNA Purification Kit (Promega) and stored at −20°C. Longissimus dorsi muscle samples for quantitative polymerase chain reaction (qPCR) expression analysis in promoter region were taken from 20 Polish HF bulls of known MEF2A genotypes. Animals were maintained at the Polish Academy of Science Experimental Farm, Jastrzębiec, and kept under the same housing and feeding conditions. The bulls were housed in a tie-stall and fed silage, hay, and concentrate ad libitum with constant access to water. After 24 h fattening, all bulls were slaughtered at the age of 12 months, with body weight about 370 kg. All tissues were harvested and immediately frozen in liquid nitrogen and then stored at −80°C. Total RNA was extracted from tissues using a Qiagen RNeasy® Fibrous Tissue Mini Kit (Qiagen) according to the manufacturer's instructions, afterward a DNase treatment was performed. The quality and quantity of RNA was examined by spectrophotometric NanoDrop measurements and by 2% agarose gel electrophoresis. Reverse transcriptions were performed on 1 μg of total RNA using Transcription First Strand cDNA Synthesis Kit with oligo primers (Roche), according to manufacturer's instructions. The resulting cDNA was stored at −20°C until use. All procedures carried out on animals were approved by the Local Ethics Commission, permission No. 29/2007.
Identification and analysis of the polymorphism
The genomic organization of the MEF2A was performed using the ScanGen (
PCR products representing different MSSCP patterns were sequenced using a 3130xl Genetic Analyzer (Applied Biosystems Applera). Sequence alignments and identification of SNPs were performed using Clustal.W (
Real-time analysis
Ten Polish HF bulls from each genotype group—TTA/TTA and TTA/GGG—were used to measure the effect of SNPs in the promoter region on MEF2A mRNA expression level in the longissimus dorsi muscle. Real-time PCR amplification was done in triplicate using an SYBR Green detection and the Roche Light Cycler 2.0 system (Roche). Real-time PCR primers were designed to anneal to adjacent exons or exon–exon junctions (Supplementary Table S1). Raw results were normalized relative to the geometric mean of mRNA detected from three reference genes SF3AI, EEFIA2, and TBP genes. The amplification efficiencies of the genes were determined using Light Cycler 3.5 Software (Simpson et al., 2000). The results were calculated using the mathematical formula for relative mRNA quantification in real-time PCR given by Pfaffl (2001). The statistical analysis between the MEF2A genotypes and mRNA expression level was performed with the Duncan's test.
Results
In total, 4094 bp were re-sequenced in 30 animals, resulting in the detection of 6 novel variants in the bovine MEF2A, specifically, three SNPs (g.-780T/G, g.-768T/G, and g.-222A/G) in the promoter (Supplementary Fig. S1), 1 InDel (g.167747C/-) in the intron 9, and 2 synonymous SNPs g.1599G/A (421aa) and g.1626G/A (429aa) in the exon 11 (Supplementary Fig. S2). Moreover, we detected a previously described SNP by Chen et al. (2010) in the exon 11, namely the synonymous g.1641G/A (434aa) substitution. The nucleotide sequences and polymorphism information for bovine MEF2A have been deposited in the GenBank database under accession No. GU211003, JN944535, and JN944536. The distribution of genotypes and allele frequencies was studied in 200 animals representing four cattle breeds (Supplementary Table S2). The distribution of genotypes showed that variations in the promoter region segregated as two intragene haplotypes TTA and GGG in the animals studied. Genotyping performed for g.-780T/G, g.-768T/G, and g.-222A/G SNPs revealed two genotypes for each locus in the HF and PR breeds, only. All studied HER and LIM animals showed variant TTA. No GGG/GGG genotype was found in all cattle breeds tested. The frequencies of the haplotype TTA and GGG for HF and PR were 0.829, 0.108 and 0.950, 0.050, respectively.
Three SNPs within exon 11 segregated as two haplotypes GGG and AAA, with higher AAA haplotype frequencies in all examined breeds (Supplementary Table S2). All genotypes for the tested SNPs distributed according to the Hardy–Weinberg equilibrium. The g.167747C/- InDel polymorphism in the intron 9 was not genotyped.
In silico analysis of the promoter SNPs using TESS software revealed that allele T at the g.-780T/G SNP creates putative binding sites for HiNF-A, SGF-1, and POU1F1a transcription factors, which are abrogated by the G allele, respectively. The SNP g.-768T/G was located within the potential sites for c-Ets-2, PU.1, and H-2RIIBP, which are abrogated or introduced by the T allele, respectively. While the g.-222A/G SNP changes the binding sites for IHF transcription factors (Supplementary Fig. S3).
The in silico transcription factor binding site analysis is in the line with the real-time PCR results that showed a genotype-dependent MEF2A mRNA expression level (p<0.01) in the longissimus dorsi muscle. The mRNA expression of MEF2A was higher in animals carrying homozygous TTA/TTA genotype, than in those with heterozygous TTA/GGG variant (Fig. 1).
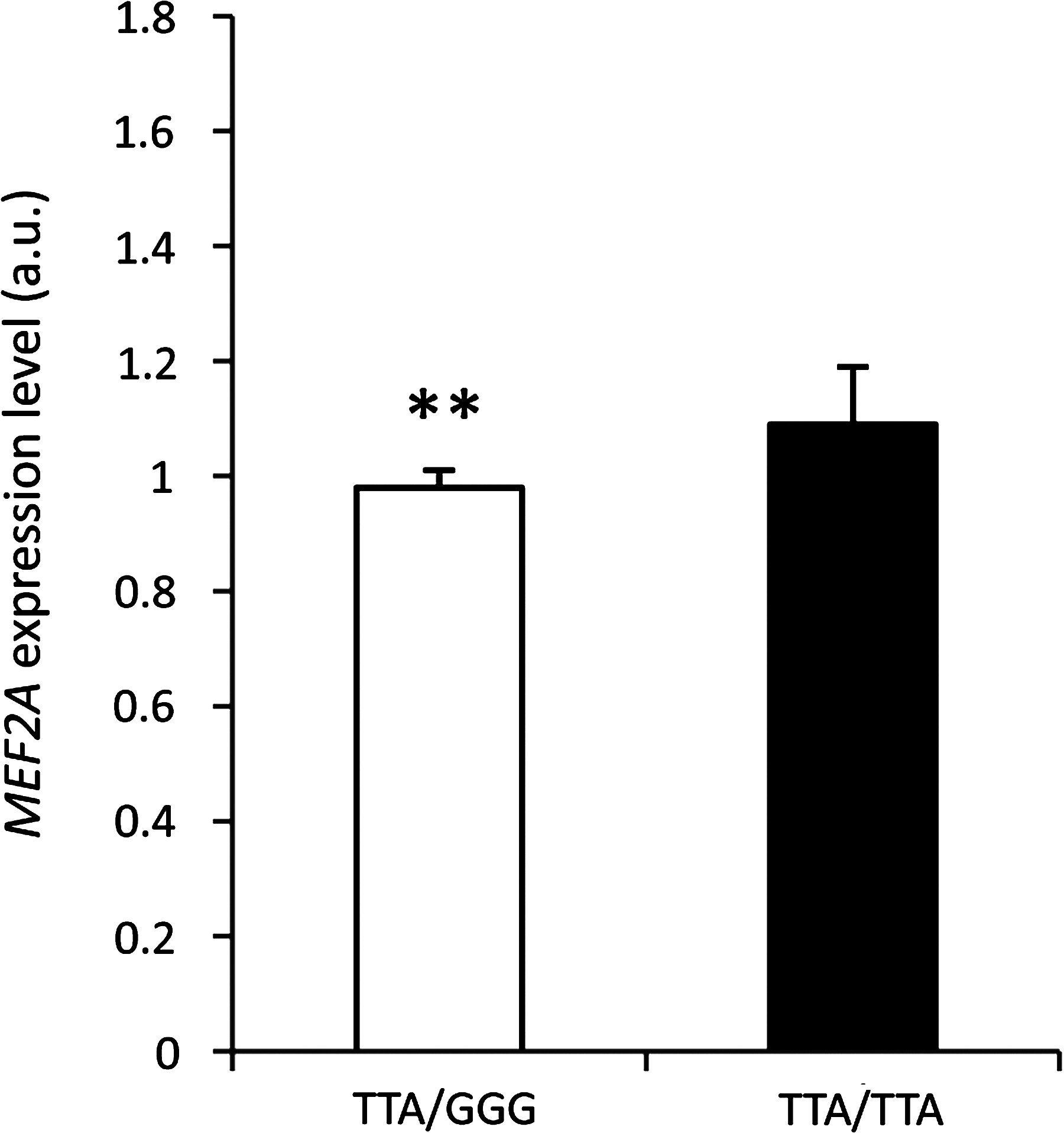
The effect of SNPs at position -780T/G, -768T/G, and -222A/G in the promoter region on the MEF2A expression level in longissimus dorsi muscle; **p<0.01.
Discussion
Members of the MEF2 family of transcription factors are upregulated during skeletal muscle differentiation and cooperate with the MyoD family of myogenic basic helix-loop-helix (bHLH) transcription factors to control the expression of muscle-specific genes (Molkentin and Markham, 1993; Black et al., 1998).
Recently, a number of published studies have clearly shown that MEF2 factors are involved in the postnatal regulation of skeletal muscle development, growth, and homeostasis (Knapp et al., 2005; Hennebry et al., 2009). Zhao et al. (2011) confirmed that muscle regulatory factor (MRF) and MEF2 families are critical for the phenotypic differences between two pig breeds and proposed a novel model myogenesis. According to these authors, MyoD and MEF2A control the balance between intermuscular adiopogenesis and myogenesis by regulating the CCAAT/enhancer-binding protein (CEBP) family, while MEF2C and Myf5 are important during the whole myogenesis process and MEF2D affects muscle growth and maturation. The expression level of MEF2 family genes is downregulated after birth but increases during skeletal muscle regeneration (Parker et al., 2003), while Musaro et al. (1995) showed that MEF2C was expressed at lower levels in newborn and young mice but increased conspicuously in adults and remained high in senile mice. The increased expression of the MEF2C in adult mice was associated with increased slow myosin isotype, indicating the possible role of MEF2C in the induction of the myogenic pattern specific for type I fibers in mature muscles (Senna et al., 2004; Hennebry et al., 2009).
Several studies have established that MEF2 factors govern the intricate process of cardiogenesis by regulating cardiac-specific gene expression (Black et al., 1998). In addition, MEF2 transcription factors are involved critically in the regulation of inducible gene expression during myocardial cell hypertrophy, inasmuch as the MEF2-binding site within the MLC2 promoter is required during PE-mediated and ET-1–mediated hypertrophy (Zhu et al., 1991) and MEF2 DNA-binding activity is increased in the hearts of rats subjected to pressure or volume overloads (Molkentin and Markham, 1993).
Several SNPs identified in the human MEF2A gene are associated with hypertrophic cardiomyopathy and coronary artery disease (CAD). Several missense mutations of the MEF2A gene were identified in CAD patients, which reduced the transcriptional activation activity of MEF2A and could increase the risk of suffering a myocardial infarction (Bhagavatula et al., 2004; Weng et al., 2005; González et al., 2007; Elhawari et al., 2010). Coto et al. (2009) reported that mutation CAG-repeat in exon 12 of the human MEF2A gene could be a risk factor for left ventricular hypertrophy.
In the current study, six novel SNPs of the MEF2A gene were identified, suggesting that the bovine MEF2A gene is lowly polymorphic and evolutionarily conserved. In addition, three SNPs in the promoter region appeared to be breed specific for the HF and PR cattle breeds. Recently, Zhou et al. (2010) described three SNPs in 5′-UTR, exon 4, and intron of the chicken MEF2A gene. In our previous study, we found in the bovine Mef2C gene two substitutions and two InDels upstream from the putative transcription start of exon 1 that were deposited in the GenBank database (Juszczuk-Kubiak et al., 2011).
In the current study, the promoter variants are associated with the MEF2A mRNA expression level in the longissimus dorsi muscle in 12-month-old HF bulls. The SNPs in the MEF2A promoter region are localized within the binding sites for transcription factors, namely HiNF-A, SGF-1, c-Ets-2, PU.1, H-2RIIBP, IHF, and POU1F1a. Salvatori et al. (2002) reported that the GHRHR promoter is regulated by POU1F1 and that promoter mutations impairing POU1F1 binding can reduce the expression of GHRHR. In addition, POU1F1 regulates GH secretion in the pituitary gland and plays crucial roles in the development and growth of mammals (Gil-Puig et al., 2005). This suggests that the transcription factor might also be involved in the cis-regulation of the MEF2A transcription in muscles. Similar effects have been noted previously in other bovine genes where mutations localized in the promoter region changed the affinity of transcription factors to the promoter sequence and acted as cis-regulators in the expression of the target gene (Flisikowski et al., 2004; Adamowicz et al., 2006). Additionally, these SNPs might be in linkage disequilibrium with the variation in other regions of the gene with functional or structural significance. So far, the potential effect of any polymorphisms in the regulatory region of the MEF2A gene on its expression in the muscles of cattle has been not reported in any available study.
It is known that MEF2 proteins act as major transducers of Ca2+ signaling events and that these events have a central role in the hypertrophic growth and remodeling of adult skeletal muscle in response to mechanical loads (Olson and Williams, 2000). Additionally, increased MEF2 expression is induced by exercise in response to acute endurance exercise, which is correlated with the increased proportion of oxidative fibers without promoting fast to slow fiber transformation (Vissing et al., 2008). It is also possible that MEF2A promoter variants could have a potential effect on MEF2A transcription levels in human muscles and might influence the homeostasis of muscles and their performance during exercise. Therefore, it is possible that postnatal skeletal muscle growth depends more on Ca2+ signaling and MEF2 proteins than on myogenic bHLH factors (Knapp et al., 2005).
Two SNPs, g.1599G/A (P421P) and g.1626G/A (P429P) in exon 11, could not change amino acids but might have caused codon alternations, which probably affected translation efficiency and the efficiency of cotranslational protein folding (Kimchi-Sarfaty et al., 2007). Additionally, proline residue confers unique structural constraints on peptide chains and markedly influences the speed of protein folding (Wedemeyer et al., 2002).
The bovine MEF2A gene is localized on BTA21 and contains 11 exons, encoding for 492 amino acids, and has a genomic structure similar to that in humans and pigs (Wu et al., 2011), and recent studies have shown that polymorphisms in the MEF2A gene are associated with body weight in chicken and cattle (Chen et al., 2010; Zhou et al., 2010). Chromosome 21 harbors 2 QTLs associated with birth weight. Davis et al. (1998) and Casas et al. (2003) detected a QTL for birth weight in the centromeric region (located at 4 centimorgans) of this chromosome, but Kim et al. (2003) found a QTL for birth weight at 62 centimorgans from the beginning of the linkage group. These implied that MEF2A might have an important role in postnatal muscle development, and is considered to be a potential molecular marker for carcass quality traits in farm animals. This information on genotype-dependent MEF2A mRNA expression can be utilized for future marker selection in association studies of carcass traits in cattle and for more detail in in vitro functional analyses.
Footnotes
Acknowledgments
This study was funded by the Ministry of Scientific Research and Information Technology, grant No. NN311034834 and by IGAB PAS project S.V.9.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.