
Abstract
Parkinson's disease (PD) is one of the most common neurodegenerative disorders characterized by decreased levels of the neurotransmitter dopamine (DA) in the striatum of the brain, as a result of degeneration of DA neurons. Levodopa (L-Dopa) crosses the blood–brain barrier and its administration replenishes the loss of DA in dopaminergic neurons in PD patients. Despite the evident beneficial effects, L-Dopa use may cause side effects and its toxicity found in in vitro assays has been attributed to the generation of reactive oxygen species (ROS): L-Dopa is converted to DA and its metabolism and autoxidation gives rise to quinones, semiquinones, and hydrogen peroxide. Despite this evidence, L-Dopa in some in vivo and in vitro experiments showed no toxic effects, or even antioxidant effects. Two major peripheral L-Dopa metabolic pathways, driven by the enzymes Aromatic L-amino acid decarboxylase (AADC) and catechol-O-methyl transferase (COMT), significantly deplete the amount of L-Dopa reaching the brain. The low bioavailability of L-Dopa may cause a wide variation in clinical response between patients. Strategies addressing to improve the bioavailability of L-Dopa include coadministering L-Dopa with carbidopa, a decarboxylase inhibitor, as multiple daily doses. We utilized catecholaminergic human neuroblastoma cells to study DNA damage and ROS production after L-Dopa and carbidopa treatments. Our data lead us to confirm that L-Dopa may have a protective effect on dopaminergic cells especially at certain concentrations, in particular, toward the production of ROS and their toxic effects on DNA. Furthermore in the combined treatment, with induction of ROS following administration of H2O2, carbidopa is effective in reducing the damage caused by reactive oxygen intermediates both alone and in combination with L-Dopa.
Introduction
Levodopa (L-Dopa) is the gold standard drug for the treatment of the motor symptoms of PD (Schapira, 2008), since its administration replenishes the loss of DA in dopaminergic neurons in PD patients (Nyholm, 2006).
L-Dopa crosses the blood–brain barrier and is converted to DA in the dopaminergic cells of the substantia nigra to be utilized as a neurotransmitter for basal ganglia function (Schapira, 2008). Moreover DA in the dopaminergic cells can be converted in neuromelanine, so DA metabolism is the focus of much of the attention relating to the toxicity of L-Dopa (Schapira, 2008).
Despite the evident beneficial effects, L-Dopa use may cause side effects and the toxicity of both L-Dopa and DA found in in vitro assays has been attributed to the generation of reactive oxygen species (ROS) (Jones et al., 2000; Olanow et al., 2004). In fact, L-Dopa and DA metabolism and their autoxidation gives rise to quinones, semiquinones, and hydrogen peroxide, which can be easily reduced in the presence of ferrous iron to hydroxyl radicals. This is of great importance in developing of PD because iron is increased in substantia nigra of patients so that oxidative stress could be seriously enhanced causing neuronal apoptotic death (Nakabeppu et al., 2007).
Two major peripheral L-Dopa metabolic pathways, driven by the enzymes, AADC and COMT, significantly deplete the amount of L-Dopa reaching the brain (Poewe et al., 2010). The short half-life (30–60 min) of L-Dopa is associated with fluctuating L-Dopa plasma levels, which eventually translate into fluctuating levels of synaptic DA derived from exogenous L-Dopa (Poewe et al., 2010).
Strategies that may improve the bioavailability of L-Dopa, include coadministering L-Dopa with carbidopa, a decarboxylase inhibitor, as multiple daily doses (Seeberger and Hauser, 2007). L-Dopa's serum half-life, when coadministered with a decarboxylase inhibitor, is increased approximately to about 90 min (Seeberger and Hauser, 2007). In vivo studies have not provided convincing data on the toxicity of L-Dopa in the treatment of PD: in some cases, L-Dopa seems to have a protective effect on dopaminergic cells (Lyras et al., 2002; Olanow et al., 2004; Prigione et al., 2006; Cornetta et al., 2009; Oli et al., 2010). In vitro studies have, however, shown discordant results, showing an adverse effect on cultured dopaminergic cells (Mytilineou et al., 1993; Ziv et al., 1997; Schapira, 2008).
In light of these conflicting data, we focused our attention on the in vitro effects of various concentrations of L-Dopa and carbidopa, alone and in combination. In particular, we aimed to verify the toxicity of L-Dopa, analysing the induction of DNA damage, and to substantiate whether this effect could appear via oxidative stress. To assess the behavior of L-Dopa in the presence of oxidative stress, we tested its effects in combination with hydrogen peroxide. Due to its important role in PD therapy, we also investigated the effects of carbidopa per sè and in combined treatment with L-Dopa and analyzed the modulation of DNA damage, even under conditions of oxidative stress induced exogenously.
For this purpose, we utilized the catecholaminergic human neuroblastoma cells, SH-SY5Y, employing standard and enzyme-modified alkaline comet assay to study DNA damage and the Dichlorofluorescein cytofluorimetric assay to analyse ROS production.
Materials and Methods
Cell line
SH-SY5Y is a human neuroblastoma cell line, cloned three times by the line SK-N-SH (human neuroblastoma cell undifferentiated) and marketed by ATCC (American Type Culture Collection).
The cells are maintained in monolayer culture in the Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal calf serum, 10 U/mL penicillin, 100 g/mL streptomycin, 2 mM L-Glu, and 0.1% gentamicin.
The cells are kept in an incubator at temperature of 37°C in humidified atmosphere of 5% CO2.
The cells were seeded in 25-cm2 flasks with 5 mL of complete medium.
Levodopa and carbidopa
Levodopa (3.4 dihydroxy-L-phenylalanine) (Sigma-Aldrich) was dissolved in a hydrochloric acid solution (HCl) 0.5 M, to obtain a stock solution with a concentration 50 mM.
The final concentrations used for the different treatments are 10, 15, 20, 50, 100, and 150 μM.
Treatments, lasting 30 min, 1, 2, or 24 h, were carried out directly in a complete medium in which cells were grown in subconfluence.
Carbidopa (Sigma-Aldrich) was dissolved in dimethylsulfoxide (DMSO) to obtain a stock solution of 20 mM (4.52 mg/mL). It is used in treatments with a 4:1 ratio L-Dopa/carbidopa, based on the quantity of the drug Sinemet®. The concentrations used for the treatments were 12.5 (for the combined treatment with L-Dopa 50 μM), 25 μM (for the combined treatment with L-Dopa 100 μM), and 37.5 μM (for the combined treatment with L-Dopa 150 μM).
The treatments, lasting 30 min, 1, 2, or 24 h, were carried out directly in the complete medium with subconfluent cells.
Citotoxicity assay
Forty thousand adherent cells were incubated for 24 h at 37°C with L-Dopa (10, 15, 20, 50, 100, and 150 μM) and with carbidopa (12.5, 25, and 37.5 μM) in the DMEM 10% FCS medium. Cells were detached with the trypsin/EDTA solution. 40 μL of cellular suspension were mixed with 5 μL trypan blue for 3 min. Cell suspension was pipetted onto a glass microscope slide and analyzed under a phase-contrast microscope, for calculating the proportion of nonstaining cells.
Standard comet assay
The alkaline comet assay was performed as described by Festa et al. (2003). Twenty microliters of cellular suspension was mixed with 180 μL 0.7% low melting point agarose in phosphate-buffered saline (PBS) (Ca and Mg free) at 37°C, and immediately pipetted onto a frosted glass microscope slide precoated with a layer of 1% normal melting point agarose, similarly prepared in PBS. Two slides were prepared for each experimental point. The agarose was allowed to set at +4°C for the necessary time, and then slides were incubated in a lysis solution (2.5 M NaCl, 10 mM Tris–HCl, 100 mM Na2EDTA, NaOH to pH=10, 1% Triton, 10% DMSO) for 50 min. After lysis, slides were placed onto a horizontal electrophoresis unit containing a fresh solution (1 mM Na2EDTA, 300 mM NaOH, pH=13) for 20 min to allow DNA unwinding. Electrophoresis was conducted for 15 min (25 V, 300 mA) at +4°C.
Subsequently, slides were gently washed in a neutralization buffer solution for 5 min (0.4 M Tris–HCl, pH=7.5), fixed in 100% fresh methanol for 3 min, and stained with ethidium bromide (2 μg/mL). Slides were analyzed using a fluorescence microscope (Leica) equipped with a camera. Fifty comets on each slide, coded and blindly scored, were acquired using I.A.S. software automatic image analysis system purchased from Delta Sistemi. To quantify the induced DNA damage, we used the% Tail DNA, which is a measure of the percentage of migrated DNA in the tail (Collins, 2004).
Formamidopyrimidine-DNA-glycosylase-modified comet assay
We used formamidopyrimidine-DNA-glycosylase (FPG)-modified comet assay to evaluate oxidative DNA damage. This test uses the FPG enzyme. FPG is a glycolase that recognizes and specifically cuts the oxidized bases principally 8-oxoguanine from DNA, producing apurinic sites converted in breaks by the associated AP-endonuclease activity. Therefore, these breaks can be detected by comet assay and give a measure of oxidative DNA damage, enabling us to detect moderate, but still appreciable damage. We followed the procedure of Collins et al. (1993), with minor modifications. The analysis of modified DNA bases was performed using a FPG FLARE™ module (Trevigen). Within the module, the manufacturer provided all the reagents used. The comet assay was carried out as described above, with the exception that after lysis, slides were washed three times for 15 min with the FLARE™ buffer (1×, pH=8). After this time, slides were incubated with 100 μL of FPG (1:100 in REC dilution buffer). Enzyme dilution was prepared right before use. Control slides were treated with 100 μL of the REC dilution buffer only. Slides were placed horizontally in the humidity chamber at 37°C for 45 min. DNA unwinding and electrophoresis were then completed as described above.
Intracellular ROS Detection
Dichlorofluorescein-diacetate (DCFH-DA) cytofluorimetric test was performed to analyze the production of ROS after L-Dopa treatment. Briefly, 200,000 adherent cells were loaded for 30 min at 37°C with 10 μM DCFH-DA/DMSO solution in the DMEM 10% FCS medium. Cells, twice washed, were incubated for further 2 hours with L-Dopa (50–100–150 μM) in 10% FCS medium. Cells were detached with the trypsin/EDTA solution and analyzed by a flow cytometer. Green fluorescence was quantify by a mean fluorescence intensity on the FL1 channel and its variation was represented as a fold increase versus untreated sample. Hydrogen peroxide treatment (10−4 M) was used as a positive control.
Statistical analysis
To assess DNA damage fluctuation, a nonparametric analysis of variance was applied. The comparison between treated and not treated cells was performed by the Mann–Whitney U test. The statistical analyses were performed using the GraphPad InStat version 3.00 (GraphPad Software). The comparison among different types of treatment (single and combined ones) was also performed through the Kolmogorov–Smirnov test (
Results
Effects of L-Dopa and carbidopa treatment: cytotoxicity assay, comet assay and DCFH analysis
In Figure 1, the measurement of L-Dopa and carbidopa effects on SH-SY5Y according to trypan blue staining is shown. Cells were treated with different concentrations of L-Dopa (10, 15, 20, 50, 100, and 150 μM) or carbidopa (12.5, 25, and 37.5 μM) for 24 h. A 30-min treatment with H2O2 was also performed. After incubation, nonstaining cells were counted with trypan blue staining. Neither L-Dopa nor carbidopa treatment induced cytotoxicity; in combined treatments, the cytotoxicity induced by H2O2 treatment was completely recovered.

Measurement of SH-SY5Y cell viability according to trypan blue staining.
In Figure 2A, we show the results of a short-time (2 h) treatment with several doses of L-Dopa. No increase of DNA damage (expressed as percentage of DNA in the Tail) was observed. In addition a longer time treatment (24 h) was performed with the three highest doses of L-Dopa obtaining a significant increase of damage with 150 μM (p<0.001).
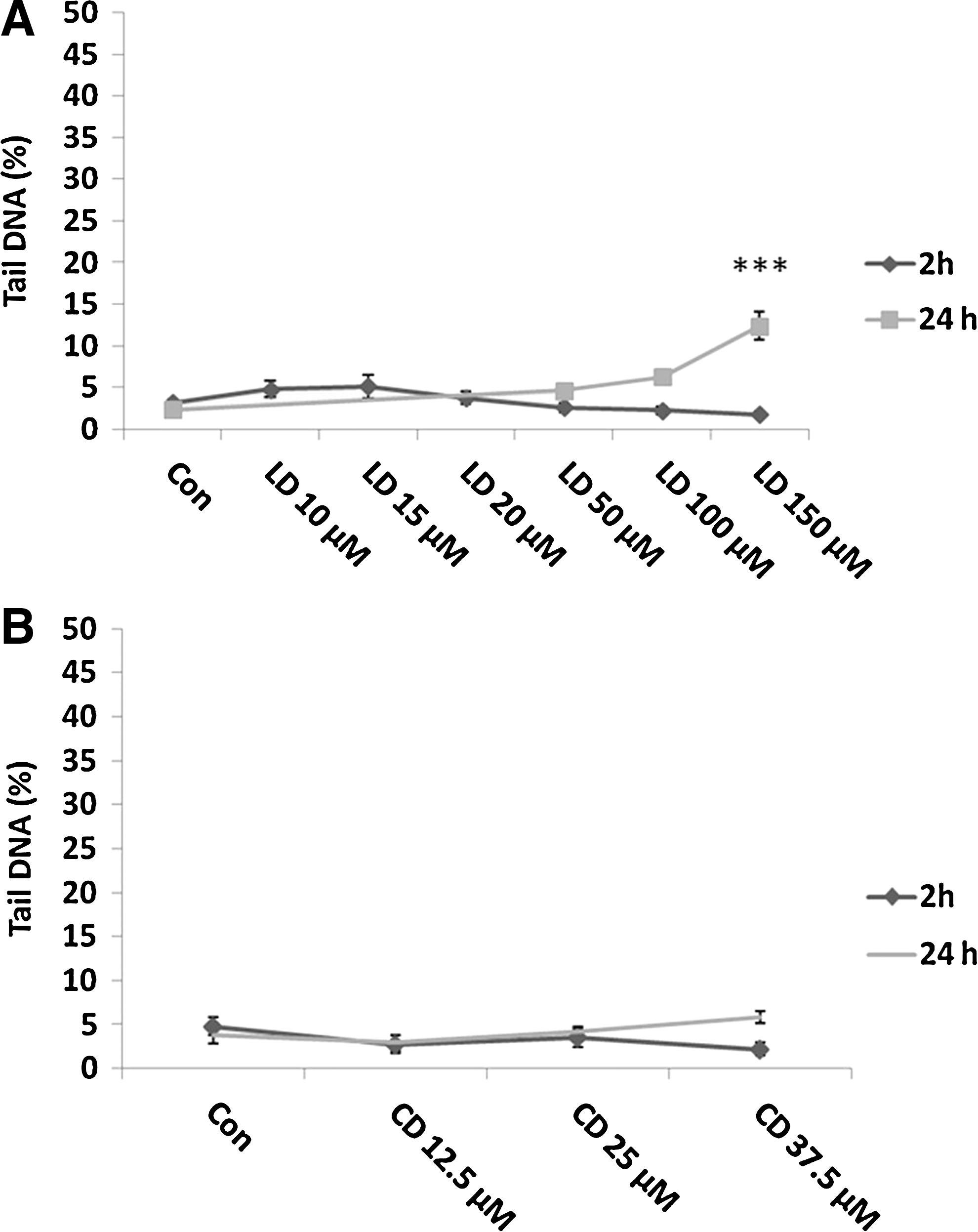
On the contrary, no effect was observed after 2- and 24-h carbidopa treatments (Fig. 2B).
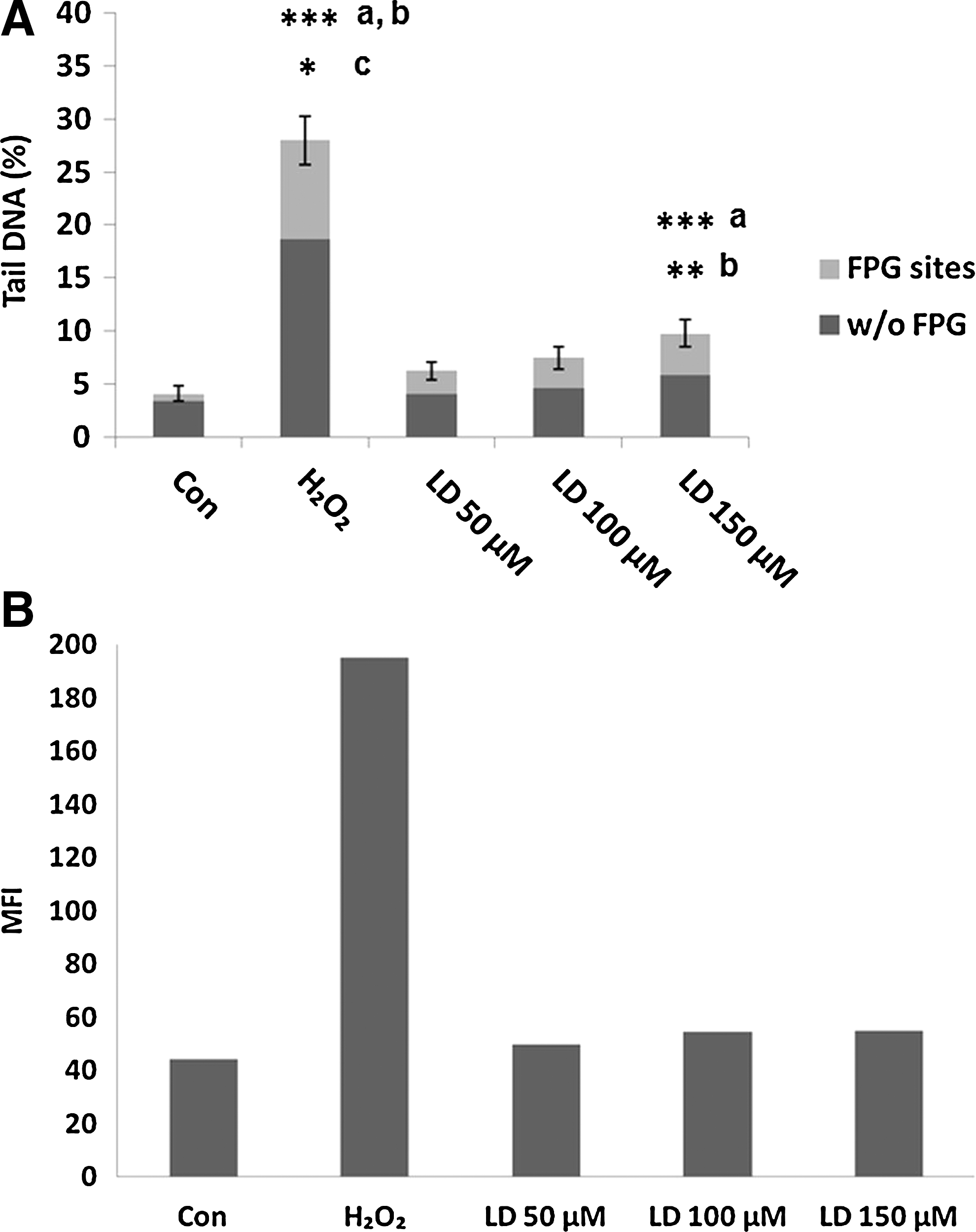
To analyze the nature of the DNA damage induced by the highest doses of L-Dopa, we performed also the FPG-modified version of comet assay to visualize the presence of oxidative lesions. In Figure 3A, we show the results obtained after 24-h treatment with 50–100–150 μM L-Dopa with both comet assay versions. H2O2 treatment was performed as a positive control. Only 150 μM of L-Dopa induced a significant increase in DNA damage. However, the increase in FPG sites registered with the modified version of comet assay is not significant when compared to the percentage of DNA damage tested with the standard comet assay. L-Dopa does not induce oxidative damage as also confirmed by data obtained with DCFH analysis: in Figure 3B, the production of intracellular ROS after 2-h treatment with L-Dopa is shown.
Protective effects of L-Dopa and carbidopa single treatment against exogenous oxidative stress
To assess the protective effects of L-Dopa and carbidopa against oxidative stress, we performed combined treatments with hydrogen peroxide (H2O2).
For this purpose, SH-SY5Y cells were treated with several doses of L-Dopa (from 10 to 150 μM) for 30 min and 1, 2, and 24 h adding H2O2 (100 μM) to the culture medium in the last 30 min.
Our results (Fig. 4A) showed that the lowest doses of L-Dopa (10 and 15 μM) induce a slight, but significant decrease in DNA damage (p<0,05) after 1-h treatment: this effect is exhausted after 2-h treatment. 20 μM L-Dopa shows greater effectiveness in reducing DNA damage, especially after 1-h treatment (p<0.001). This effect is exhausted after 2-h treatment. When the experiments with higher doses were performed, a longer treatment time (24 h) was added. In these experiments, we observed a very significant decrease (p<0.001) of H2O2-induced DNA damage with all used doses at earlier times: the protective effects persisted at a longer time increasing L-Dopa dose. In fact, while 50 μM was effective only after 30 min and 1 h, 100 μM decreased the damage also at 2-h treatment time. The highest dose (150 μM) showed its protective effect not only during the shorter treatment times, but the effect persisted also after 24-h treatment.
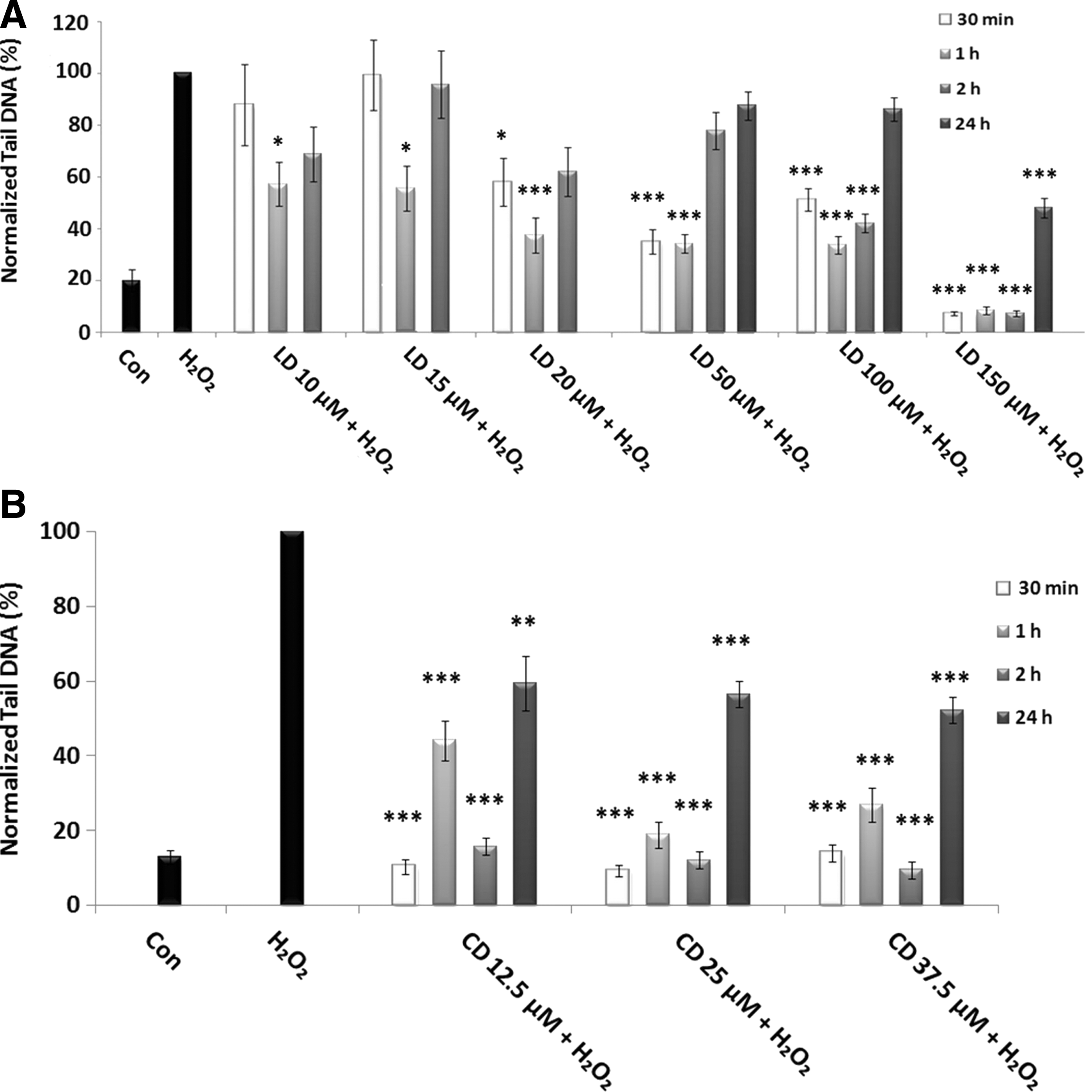
Protective effects of LD and CD in SH-SY5Y cells evaluated by alkaline comet assay.
As far as carbidopa is concerned, the treatment was always very efficient in reducing H2O2-induced DNA damage, with all used doses and at all times of treatment (Fig. 4B).However, at 24 h, the effect of reduction is less pronounced.
Protective effects of L-Dopa and carbidopa combined treatments against exogenous oxidative stress
As previously mentioned, PD therapy involves the use of a combination of L-Dopa with a Dopa-decarboxylase inhibitor. For example, the Sinemet® drug consists in a combination of L-Dopa/carbidopa in a 4:1 ratio. For this reason, to test the effect of combined treatment, L-Dopa/carbidopa on neuroblastoma cells, we decided to treat SHSY-5Y cells with 12.5, 25, or 37.5 μM carbidopa combined respectively with 50, 100, or 150 μM L-Dopa: these doses of L-Dopa, in fact, have shown greater efficacy in the reduction of DNA damage and had an effect that persisted longer than 10, 15, and 20 μM, allowing us to analyze the long-term effects of combined treatments. In the last 30 min, H2O2 was added.
The results shown in Figure 5 revealed a significant reduction of H2O2-induced DNA damage in all experimental conditions, except for the lowest doses at the shortest time (50 μM L-Dopa+12.5 μM carbidopa at 30 min of treatment). However, the most striking results concern the combination of highest doses (150 μM/37.5 μM). In fact, after 2-h treatment, the combined effect is not higher than the effect of single treatments (see also Fig. 4); in contrast, at the longest treatment time (24 h), the combined treatment reverted the H2O2-induced DNA damage to control level, so revealing a much more efficient action than single ones.
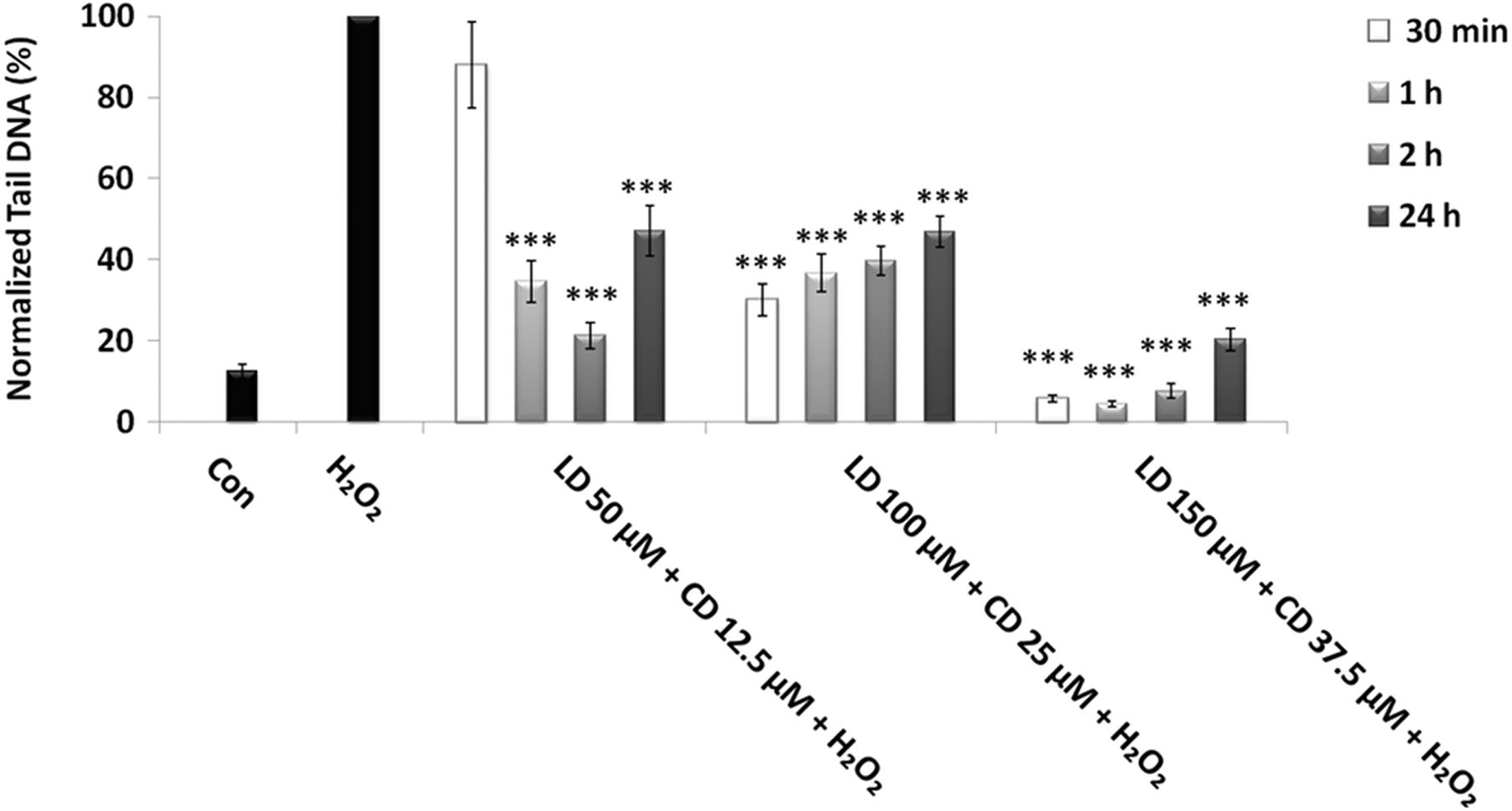
Protective effects of combined treatment LD/CD in a 4:1 ratio in SH-SY5Y cells evaluated by alkaline comet assay. The protective effect in SH-SY5Y cells after 30-min-, 1-, 2-, and 24-h treatments with various concentrations of LD/CD in the presence of H2O2 is evaluated in terms of reduction of H2O2-induced DNA damage. Values of Tail DNA are normalized with respect to the percentage of DNA damage induced by H2O2 single treatment (100%) and represent the mean results of three experiments±SE. H2O2: 100 μM; 30-min treatment. ***p<0.001 (at Mann–Whitney U test) treated cells versus H2O2-treated ones.
To better evaluate these results, we performed also a more detailed statistical analysis based on the distribution pattern of damage values. In Figure 6, we show the two most informative situations, that are the single and combinated treatments with the highest doses of both molecules after 2 and 24 h with H2O2, in addition to the effect of H2O2 alone. After 2-h treatment (Fig. 6A), no statistical difference (at the Kolmogorov–Smirnov test) was observed between the DNA damage distributions comparing L-Dopa treatment or carbidopa treatment with a combined one. Indeed, as we can see from the Figure 6A, the three curves representing different treatments follow a pattern that overlaps almost completely. In other words, the single treatments cause a reversal of DNA oxidative damage comparable to that of a combined one.
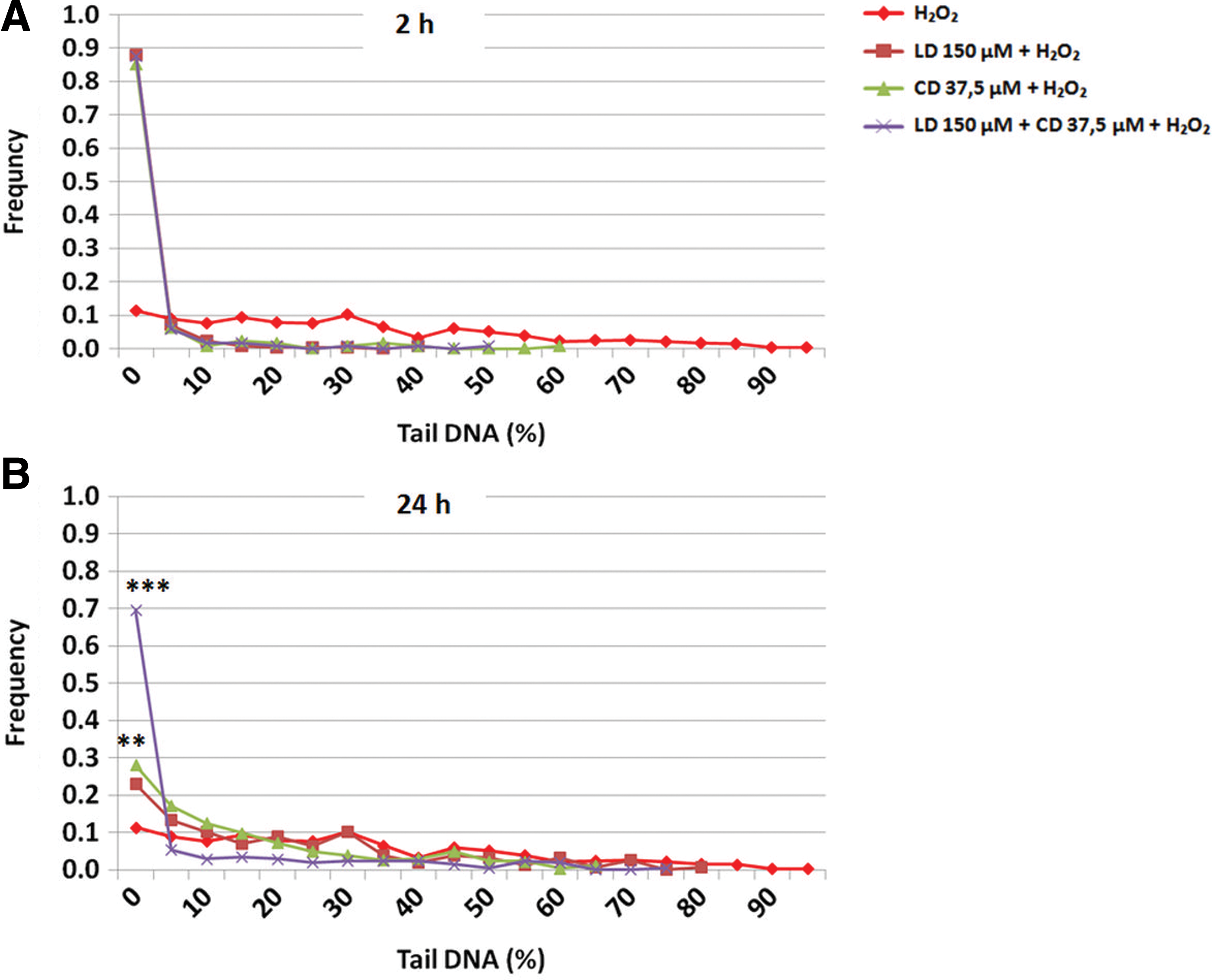
DNA damage distribution in combined treatments LD/CD in a 4:1 ratio compared with LD and CD single treatments in the presence of H2O2 in SH-SY5Y cells: on Y axis frequency of cells; on X axis classes of DNA damage expressed as Tail DNA.
In contrast, after 24 h (Fig. 6B), all comparisons (combined treatment vs. single L-Dopa one p << 0.001; combined treatment vs. single carbidopa one p << 0.001) gave highly significant results, the combined treatment being much more effective in reducing DNA damage than single treatments as evidenced by the presence of 70% of cells in the less damaged cells category. Furthermore, carbidopa treatment seems also more effective than L-Dopa one as evidenced by the presence of more cells in the less damaged cells categories (single carbidopa treatment vs. L-Dopa one p<0.01).
Discussion
PD is caused by the degeneration of dopaminergic neurons of the substantia nigra pars compacta. The etiology is still unclear, but several lines of evidence support the involvement of DA (Spina and Cohen, 1989; Jiang et al., 2008). DA toxicity may be caused by autoxidation and enzymatic oxidation of AADC, COMT, and monoamine oxidase. Previous studies demonstrated that DA-induced cytotoxicity in neuronal cells is mediated by ROS and mitochondrial inhibition (Miyazaki and Asanuma, 2008).
Although L-Dopa has been shown to be an effective medication, there are still disputes about the possibility that it can be toxic to DA neurons and can generate ROS through the metabolism of DA, thus enhancing the conditions of oxidative stress already present in patients with PD. In vivo studies have not provided extensive evidence to suggest toxicity of treatment with L-Dopa in Parkinsonian patients, and in some cases, in animal models, L-Dopa administration seems to exert a protective effect on dopaminergic cells (Lyras et al., 2002; Olanow et al., 2004; Prigione et al., 2006; Cornetta et al., 2009; Oli et al., 2010). In vitro studies have revealed rather some discordant data showing that L-Dopa may have adverse effects in cultures of dopaminergic neurons (Mytilineou et al., 1993; Ziv et al., 1997; Schapira, 2008) due to mechanisms that mediate the formation of free radicals (Hoyt et al., 1997; Mytilineou et al., 2003; Hattoria et al., 2009). Conversely, some results show that L-Dopa has a protective effect on cultures of dopaminergic neurons (Mena et al., 1997; Shi et al., 2002) even after exposure to pro-oxidants (Han et al., 1996).
To investigate the effect of L-Dopa, we conducted in vitro studies on neuroblastoma cells SH-SY5Y. These cells are an excellent model because they have the biochemical characteristics of dopaminergic neurons (Xie et al., 2010).
The purpose of the first series of experiments was to evaluate the effect of L-Dopa in terms of DNA damage in neuroblastoma cells.
L-Dopa treatment induced a slight increase in DNA damage only with the highest dose, as shown in other studies (Ziv et al., 1997; Schapira, 2008). However, we excluded this increase, which might be due to oxidative damage, since neither FPG-modified comet assay which recognizes DNA bases modified by oxidation (Collins et al., 1996) nor DCFH analysis gave significant results. Anyway, it should be taken into account that many of the studies demonstrating L-Dopa toxicity used relatively high concentrations (>50 μM), while peak plasma concentrations in patients are in the range of 10–20 μM and only 12% of an oral dose appears in the cerebrospinal fluid (Olanow et al., 1991).
The second aspect to be taken into account is the protective effect exerted by L-Dopa against H2O2-induced DNA damage. Here we reported a significant reducing effect with all used doses, particularly evident with the highest ones. This result, in agreement with previous one (Shi et al., 2002), shows for the first time a close relationship between L-Dopa concentrations and treatment times.
One of the therapies now considered more effective in treating symptoms of PD is L-Dopa in association with a Dopa-decarboxylase inhibitor, such as carbidopa (Hadjiconstantinou and Neff, 2008; Muller, 2009; Solla et al., 2010; Tayarani-Binazir et al., 2010). The current formulation of the drug Sinemet® consists of a L-Dopa/carbidopa 4:1, which can inhibit the activity of Dopa-decarboxylase to increase the half-life of L-Dopa (Seeberger and Hauser, 2007; Goole and Amighi, 2009).
In this regard, we first investigate the effect of carbidopa on neuroblastoma cells, both in a single treatment and in treatments involving induction of oxidative stress with hydrogen peroxide.
We show that carbidopa per sè does not induce DNA damage, but rather is very effective in reducing the damage caused by reactive oxygen intermediates. In combination with L-Dopa in the proportions used in pharmacological therapy (Goole and Amighi, 2009), the reducing effect is even more evident. Furthermore, it is very important to emphasize the effect of the highest dose in the combined treatment. Even if 150 μM L-Dopa seems to exert a slight DNA damage on neuroblastoma cells, in the combined treatment with carbidopa a highly protective effect in the presence of exogenously induced oxidative stress has been revealed, reverting the percentage of DNA damage almost to control level. This result allows us to assert that carbidopa acts decisively both in prolonging L-Dopa half-life, as previously shown (Seeberger and Hauser, 2007; Goole and Amighi, 2009), and further protecting cells against oxidizing effects of metabolic intermediates of L-Dopa.
It has been previously demonstrated in an in vivo perfusion system that the concomitant administration of carbidopa and L-Dopa blocks the generation of OH• due to the autoxidation of DA (Obata and Yamanaka, 1996). So, carbidopa probably exerts its antioxidant effect also by inhibiting the formation of intermediates of DA oxidation.
The combination of literature data with ours leads us to emphasize the determinant contribution of carbidopa, which allows, in combination treatment with L-Dopa, to lower the dose useful to preserve L-Dopa protective effect also at shorter times.
The doses we used in these experiments are higher than those in plasma (and brain) after an oral dose intake. In fact, the concentration peak reached in the plasma of patients is between 10 and 20 μM after each administration for a maximum of five doses per day (Stocchi et al., 2005; Kuoppamaki et al., 2009). However, after the first year of therapy, patients need to increase the daily dose to limit the worsening of motor symptoms. In this context, here we show that the combination of carbidopa with the highest dose of L-Dopa eliminates the pro-oxidant action of L-Dopa and extends the antioxidant effectiveness of the treatment up to 24 h.
Conclusions
The results obtained from previous experiments carried out to assess the effects of L-Dopa in the treatment of PD are often conflicting (Shi et al., 2002; Olanow et al., 2004; Schapira, 2008). The most difficult aspect of experimental analysis is related to the individual responses of each patient, both regarding the effectiveness of therapy and its side effects. The discrepancy of the results obtained in recent years, probably, is due to the intrinsic nature of the molecule of L-Dopa, which has both pro-oxidant and antioxidant nature depending on concentrations used. The presence of conflicting data led us to evaluate the effect of L-Dopa at different concentrations and at various times of treatment in dopaminergic cells. In our hands, the concentrations of L-Dopa, even much higher than physiological ones, have not proven damaging to DNA by themselves, but rather protective against exogenous oxidative damage.
It is also very important the additional assessment of other molecules used in combination in the treatment of parkinsonian patients. In particular, here we show that carbidopa has an antioxidant effect by itself, confirming the validity of its use in the treatment of PD. The scientific debate on the function of carbidopa in pharmacological therapy is still open (Tayarani-Binazir et al., 2010) and our data contribute to clarify the issue. In fact, we show that the presence of carbidopa makes the therapy much more protective extending the action of L-Dopa over time and possibly allowing to reduce the frequency of daily dosing. Since these data arise from an in vitro analysis, it should be important to consider the large number of physiopathological variables involved in in vivo metabolic pathways, although the cell line we used is a good model for the study of PD. However, we believe that data obtained by this in vitro investigation may provide important clues for the evaluation of the effects of the individual substances and their combination, to improve pharmacological therapy.
Footnotes
Acknowledgments
We thank Francesco Di Domenico for his contribution in statistical analysis of our data.
Research Grants from Don Carlo Gnocchi Foundation, Milan, Italy.
Disclosure Statement
No competing financial interests exist.