
Abstract
The sperm nuclear basic proteins (SNBPs) of the marine annelid worm Chaetopterus variopedatus have been shown previously to consist of a mixture of two SNBPs: histone H1-like (CvH1) and C.variopedatus protamine-like (CvPL). Here, we report the structural characterization of CvPL. The protein has a molecular weight of 8370.5 Da, a K/R ratio of 0.34, and a secondary structure, which are intermediate between those of protamine (P) and protamine-like (PL) SNBPs. The N-terminal sequence of CvPL shows a high extent of similarity with the arginine-rich C-terminal domain of chordate PL-type SNBPs. Furthermore, the protein binds to DNA in a similar fashion as vertebrate PLs and their own CvH1, but in a way that is different from that of the lysine-rich somatic H1 histones. We have experimentally determined the molar ratio CvH1:CvPL to be ∼1:6 in C. variopedatus sperm. Based on all of these, a model is proposed for the organization of the sperm chromatin by CvH1 and CvPL.
Introduction
However, the little information available to date on the PL protein-mediated organization of chromatin derives mainly from invertebrate organisms (Ausió and van Holde, 1987; Casas et al., 1993). In this sense, the marine worm Chaetopterus variopedatus is the only invertebrate described to date with the sperm chromatin organized by a histone H1-like (CvH1) (Salvati et al., 2008) and a molecule defined as a protamine (De Petrocellis et al., 1983). The latter contains arginine and lysine, and therefore it could now be considered a PL-type protein. However, its arginine+lysine content is higher than 50 mol% (56.1 mol%) with arginine and lysine corresponding to 41.7 mol% and 14.4 mol%, respectively, and it does not contain cysteine. On these bases, it cannot be classified into one of the four basic categories (PL-I, PL-II, PL-III, and PL-IV) described in mollusks. Further, this molecule forms stable oligomers, and we have already shown, by specific amino acid side chain modifications, that the oligomeric structure depends on anion-mediated lysine–arginine interactions (Piscopo et al., 1993).
In this article, we structurally characterize the SNBP protamine component of C. variopedatus. Quite unexpectedly, the partial N-terminal primary structure of the protein reveals a strong similarity to the C-terminal tail of the highly arginine-rich PLs that are present in the sperm of several chordates (Lewis et al., 2004a; Saperas et al., 2006). The secondary structure of this protein is reminiscent of that exhibited by SNBPs of the P-type and short PL proteins (Toniolo et al., 1979; Frehlick et al., 2007).
Materials and Methods
Materials
All chemical reagents were obtained from Sigma-Aldrich. Annelid worms C. variopedatus were kindly provided by the Zoological Station of Naples (Italy). Specimens were collected in the bay of Naples in July and August, when they are sexually active. Sperm cells were obtained by cutting the parapods of the worms as described in De Petrocellis et al. (1983).
Extraction and purification
Sperm cells of individual males were purified by several centrifugation washes in Millipore-filtered sea water. C. variopedatus protamine-like (CvPL) was prepared following the procedure of De Petrocellis et al. (1983) using acid extraction and CM-cellulose column chromatography, and purity was determined according to De Petrocellis et al. (1980) and reverse phase–high-performance liquid chromatography (RP-HPLC).
Extinction coefficient of CvPL and CvH1
The extinction coefficient of CvH1 and CvPL at 230 nm was determined by an amino acid analysis using Norleucine as an internal standard (Frehlick et al., 2007). The values thus obtained were 1.74 and 4.32 cm2 mg−1 for CvPL and CvH1, respectively.
Trypsin digestion
CvPL proteolitic digestion was carried out with trypsin (type III) (Sigma-Aldrich). Aliquots of 333 μL (1 mg/mL) of CvPL in 10 mM Tris-HCl buffer, pH 8.0, were incubated at room temperature with 5 μL of trypsin (40 ng/μL), for a final E/S ratio of 1:5000 (w/w), in presence or in absence of 1 M NaCl. Aliquots of the digestion mixture were collected at different times, immediately frozen in an alcohol and a solid-CO2 bath, and stored in a freezer at −20°C, until used for the electrophoresis study (see below). An overnight digest in the presence of 1 M NaCl was carried out using 280 μg of CvPL, and the enzyme conditions were described above. The reaction was stopped by addition of a twofold (w/w) excess of tosyl-L-lysine chloromethyl ketone (TLCK). After addition of TLCK, the sample was diluted to a final NaCl concentration of 0.3 M and immediately subjected to HPLC fractionation.
Electrophoresis
Protein samples were analyzed on acetic acid–urea polyacrylamide gels (AU-PAGE). Polyacrylamide gels were prepared according to the method of Panyim and Chalkley (1969), but with 15.0% (w/v) acrylamide (acrylamide:bisacrylamide 30:0.8). Gels (83×58×0.75 mm) were run for 50 min at 150 V. High-resolution urea-acetic acid-PAGE (HrAU-PAGE) was carried out according to Frehlick et al. (2007). Proteins were fixed on gel; stained using 0.2% Amido Black, 30% methanol, and 10% acetic acid; and destained with 30% methanol and 10% acetic acid.
Tricine sodium dodecyl sulfate (SDS)-PAGE was carried out as described by Schägger (2006), using the stacking gel at 5%, the spacer gel at 10%, and the separating gel at 16.5% in 6 M urea. Aliquots of 20 μg of proteolytic mixture were incubated in the presence of a sample buffer in a ratio 2:1 (v/v) at 37°C for 15 min. Electrophoresis was carried out in a vertical gel apparatus mini-protean (20×20×0.12 mm) (Biorad); an initial voltage step of 100 V was applied until the sample has completely entered the stacking gel, and a next voltage step of 200 V was applied; at the end of the run, voltage was gradually increased to 300 V to shorten the total time of electrophoresis. Samples analyzed on Tricine SDS-PAGE were silver stained (Oakley et al., 1980).
HPLC analysis
RP-HPLC was carried out using a Vydac C18 4.6×250-mm column. In the case of the trypsin-resistant peptides, the column was eluted using a linear gradient from 0.1% trifluoroacetic acid to 40% acetonitrile in 80 min at a flow rate of 1 mL/min. Under these conditions, the peak corresponding to the trypsin-resistant peptide eluted at 12% acetonitrile. For the fractionation of CvH1 and CvPL, the column was eluted under the same conditions, but using a linear acetonitrile gradient to 60% in 60 min.
Far-ultraviolet circular dichroism spectroscopy
The circular dichroism analysis was carried out using a model J-710 spectropolarimeter (Jasco), which was equipped with a Neslab RTE-110 temperature controller (Neslab Instruments) and calibrated with a standard solution of (+)-10-camphorsulfonic acid. Circular dichroism measurement in the far-UV was performed on samples of CvPL at a protein concentration of 0.1 mg/mL in 50 mM sodium phosphate buffer, pH 7.0, using cuvettes of 0.1-cm optical path length (Hellma). Photomultiplier absorbance did not exceed 500 V in the spectral regions measured. Each spectrum was signal-averaged at least five times and smoothed with circular dichroism (CD) software Spectra Manager Ver. 1.53 (Jasco). All measurements were performed at the temperatures of 20°C under a nitrogen flow (3 L/h).
Mass spectrometry
Positive Reflectron Matrix-Assisted-Laser Desorption/Ionization (MALDI) spectra were recorded on a Voyager DE STR Pro instrument (Applied Biosystems). The Matrix-Assisted-Laser Desorption/Ionization matrices were prepared by dissolving 20 mg of 2,5-Dihydroxybenzoic acid (DHB) or 10 mg of α-cyano (α-cyano-4-hydroxycinnamic acid) in 1 mL of acetonitrile/0.1% trifluoroacetic acid (70:30 v/v). Typically, 1 mL of the matrix was applied to the metallic sample plate, and 1 mL of analyte was then added. Acceleration and reflector voltages were set up as follows: target voltage at 25 kV, first grid at 92% of target voltage, and delayed extraction at 500 ns. The individual spectra were analyzed using Applied Biosystems software.
Sequence analysis
Amino acid sequence determination was performed with the Applied Biosystems Procise 492 automatic sequencer, equipped with on-line detection of phenylthiohydantoin amino acids. Protein sequence analyses were performed using software available on the ExPASy Proteomics Server (
Plasmid DNA gel mobility retardation assay
The reported amounts (see Fig. 4) of C. variopedatus SNBP mixture were added to 600 ng of circular and linearized pGEM3 DNA in 30 μL of 89 mM Tris-borate buffer pH 8.0 and 2 mM ethylenediaminetetraacetic acid (TBE). After 10-min incubation at room temperature, the samples were loaded on 1.8% or 1% agarose gel in TBE. Electrophoretic conditions and visualization of DNA were carried out as described in Piscopo et al. (2010).
Results
CvPL has a molecular weight that is intermediate between that of P- and PL-types
CvPL has an oligomeric structure in solution that depends on the presence of both free lysine amino groups and arginine guanidino groups, as shown when comparing gel filtration patterns of the native to that of the modified molecules (Piscopo et al., 1993). Furthermore, as it is the characteristic of SNBPs of the P- and PL-types, the protein precipitates upon addition of SDS (De Petrocellis et al., 1983), making it difficult to estimate its molecular mass by SDS-PAGE. Therefore, the molecular weight of CvPL was carried out by mass spectrometry.
The molecular weight of 8370.5 Da determined by MALDI for the CvPL (Fig. 1A) is intermediate between those of P- and PL-types (Table 1). Moreover, the K/R ratio of this protein determined from its amino acid analysis composition exhibits a value of 0.34, which is similar to that of the ancestral protamine precursors observed in arginine-rich chordate PL-type SNBPs (Eirín-López and Ausió, 2009) (Table 1).

Structural characterization of CvPL.
The values relative to Cv PL and Cv H1+Cv PL are in bold.
Number of lysine residues; #number of arginine residues; aCAC94904.1; bNP_999714.1; cAAI22614.1; dNP_001038138.1; eNP_005316.1; fAAH00145.1; gNP_006017.1; hNP_005314.2; iAAP32169.1; jP83214.1; kAAC50486.1; lNP_038665.1; mAAA30741.1; nB02669; oP69011.1; pTYTUZ2; qQ86QH9.2; rQ86QI0.1; sABC18201.1; tABC18200.1; uB3EWJ1; vThis publication; w(Lewis et al., 2004a); x(Lewis et al., 2004a); yAQ01227.1; zABY89927.1; αQ08GK9.2.
CvPL has a CD spectrum similar to that observed for P- and PL-types
The purified CvPL was structurally characterized by circular dichroism. The CvPL spectrum carried out under pH and ionic-strength buffer conditions close to physiological values (Fig. 1B) showed similarity with spectra obtained for SNPBs from other PL- and P-types, including human protamines (Ausió and Subirana, 1982a, 1982b; Gatewood et al., 1990). With a negative band at about 200 nm, this spectrum is very close to those that have long been observed for charged polypeptides presumed to be in the so-called random coil conformation (Johnson, 1988). The secondary structure estimation using the method described by Yang (Chen and Yang, 1971) indicated the presence of a high content in random (64.6%) and β-turn (28.0%) structures, the absence of α-helix, and a slight content in β-sheet (7.4%). Generally, in solution, most protamines have CD spectra similar to the CD presumed to correspond to denatured proteins (Toniolo et al., 1979). Although these proteins could have relatively well-defined topologies, they do not have the regular repeating pattern or defined geometry as those observed in any of the canonical secondary structure features found in other proteins (Janes and Wallace, 2009).
Therefore, like most SNBPs of P- and PL-types, CvPL appears to belong to the family of intrinsically disordered proteins (Hansen et al., 2006) that, while exhibiting a low secondary structure in solution, they may acquire an ordered secondary structure conformation upon interacting with DNA as it has been shown for the N- and C-terminal tails of H1 histone (Vila et al., 2001; Roque et al., 2005).
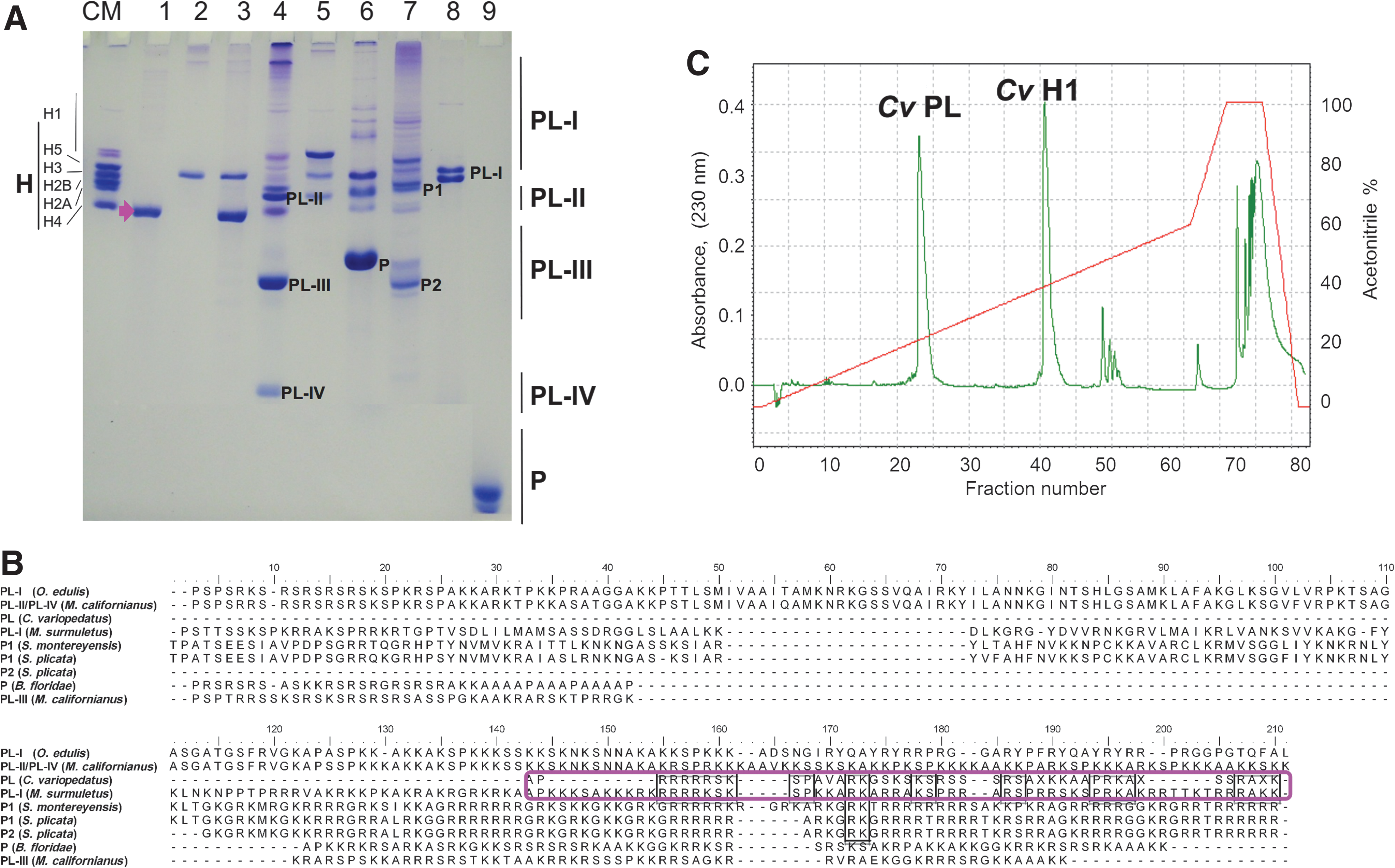
CvPL has an electrophoretic mobility and an amino acid sequence similar to PL-type and consists of discrete short domains that are resistant to trypsin digestion under high ionic strength conditions
Figure 2A shows a HrAU-PAGE of CvPL in comparison to other SNBPs from invertebrates and chordates. Under these electrophoretic conditions, CvPL migrates in the gel in the region between that of PL-II and PL-III types of bivalve mollusks (Ausió, 1986), and far from the region of P-type proteins, suggesting its closeness to SNBPs of the PL-type family. In agreement with this, the N-terminal sequence determined for CvPL (which comprises ∼55% of its whole sequence) (Fig. 2B) did not show similarity with the known P-type sequences (data not shown), while it appeared to have similarity with the C-terminal domain of some PL-type proteins (Fig. 2B). In particular, and quite unexpectedly, the closest similarity was with those from the PLs of the tunicates Styela plicata and S. montereyensis (Lewis et al., 2004a) and the fish Mullus surmuletus (Saperas et al., 2006). All of these suggest a potential structural relatedness of CvPL to the PL-type of invertebrate and chordate SNBPs.
CvPL N-terminal is related to the C-terminal end of arginine-rich PL-type SNBPs.
As shown in Figure 3A, the CvPL protein is quite unique in that, in the presence of 1 M NaCl, it exhibits the presence of short protein domains, with an average molecular weight of 850 Da (Fig. 3B), that are resistant to trypsin digestion. The protein is completely digested under the same conditions in the absence of NaCl. Mass spectrometry of the HPLC-purified-resistant peptide reveals the presence of a complex mixture of peptides of molecular weight at 820–870 Da as well as other smaller components at 650 Da. The presence of these small salt-dependent trypsin-resistant fragments, in this highly basic protein, is intriguing, but is possibly related to the lysine–arginine-mediated oligomerization exhibited by CvPL (Piscopo et al., 1993) that upon association probably results in small segments (6–8 amino acids) that are refractory to trypsin digestion.
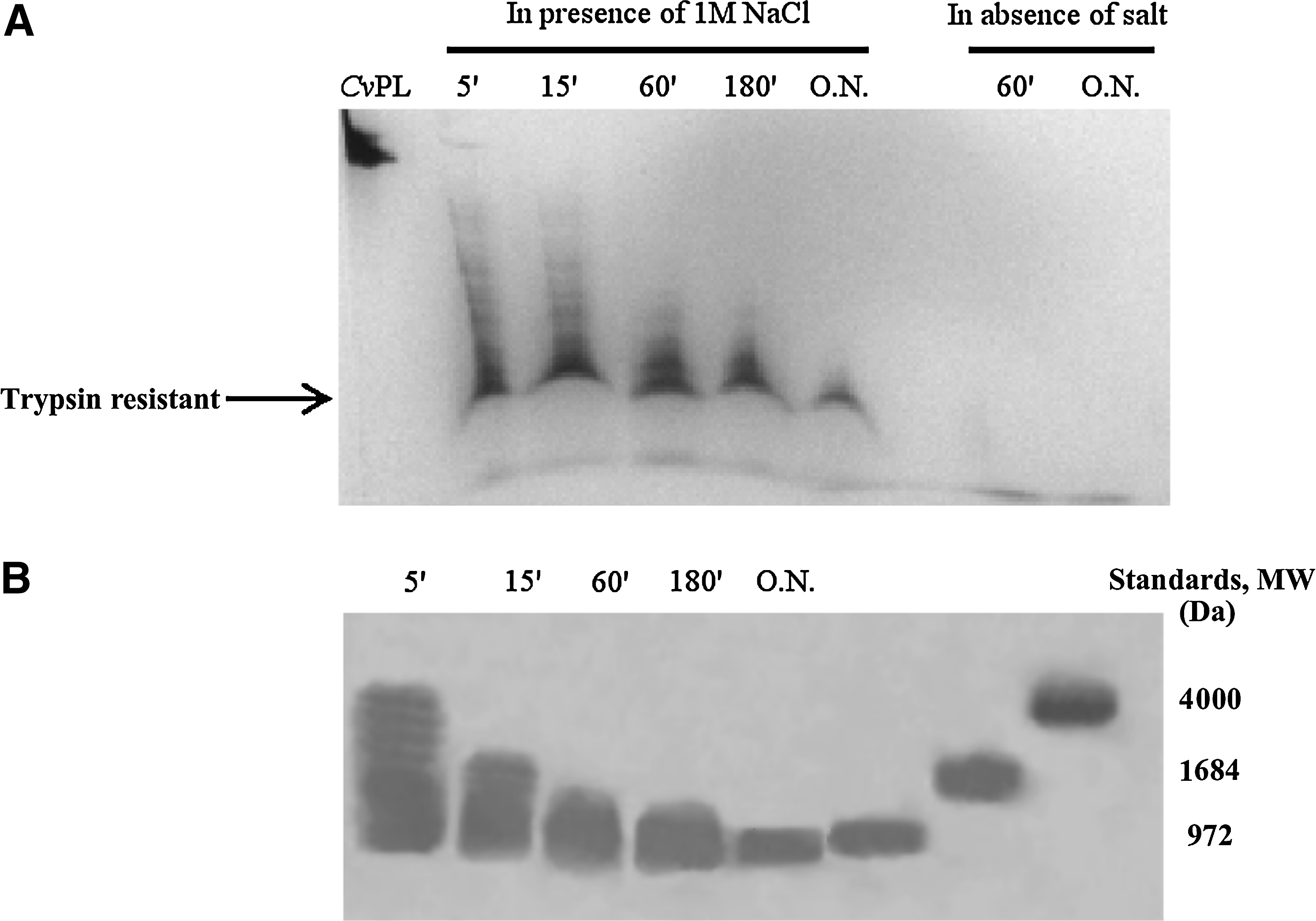
CvPL contains salt-dependent trypsin-resistant domains.
SNBPs in C. variopedatus consist of a mixture of approximately six molecules of CvPL per each molecule of CvH1
The relationship of CvPL to PL-types is intriguing, especially in regard to the potential chromatin organization resulting from its interaction and that of CvH1 with DNA in the sperm of C. variopedatus. An insight into this question can be gained from the determination of the molar ratio between these two proteins. We used two different methods for such determination. In the first approach, the ratio was determined by scanning densitometry of the two electrophoretic bands corresponding to these two proteins (see Fig. 2A, lane 3). The analysis was performed using an electrophoretic titration with different amounts of the purified CvPL and CvH1 (Fig. 2A, lanes 1 and 2). A value of 5.6 molecules of CvPl per molecule of CvH1 was obtained in this way. In a second approach, the areas of the peaks corresponding to each of the proteins in the HPLC chromatogram (shown in Figure 2C) were determined. These values were then used in conjunction with the molecular weights (CvPL: 8370 Da; CvH1 20,270 Da) of the proteins and their corresponding extinction coefficients at 230 nm (determined as indicated in section 2.3) to calculate a molar ratio of 5.8 molecules of CvPL per molecule of CvH1. Therefore, it appears from these two independent methods that ∼6 molecules of CvPL coexist with one molecule of CvH1 in the sperm chromatin of C. variopedatus.
C. variopedatus SNBPs bind to DNA in the same way as CvH1 and differently from somatic histone H1
We have previously studied the interaction of CvH1 with linear and circular DNA (Piscopo et al., 2010), and we have shown that CvH1 binds in a way that is different from canonical lysine-rich histone H1 (such as calf thymus H1) (Cole, 1987). We have also shown that this differential binding is dependent on the arginine presence. If CvH1 and CvPL are structurally related to commonly organize the C. variopedatus sperm chromatin, one would expect that these two proteins would bind to DNA in a similar fashion. Therefore, we decided to study the interaction of the C. variopedatus SNBP mixture (see Fig. 2A, lane 3).
Figure 4 shows the interaction of C. variopedatus SNBP mixture with linear and circular pGEM3 DNA, in comparison to CvH1 and calf thymus H1 (Piscopo et al., 2010). As it can be seen in Fig. 4C, the mixture of C. variopedatus SNBPs binds to DNA in a way that is almost indistinguishable from that of CvH1, reinforcing the hypothesis that these proteins are probably related. Interestingly, the linear DNA-binding saturation takes place at a (w/w) protein/DNA ratio of 1.2 for both CvH1 and C. variopedatus SNBP mixture. The molecular mass per positive charge of CvH1 and of C. variopedatus SNBPs (CvH1-CvPL considered as a joint molecule) determined from their amino acid analysis composition (De Petrocellis et al., 1983) was 288.5 and 242, respectively. Since the complete shift takes place at a 1.2 protein/DNA (w/w) ratio, this corresponds very closely to charge saturation [1 mole positive charge/1 mole of nucleotide (negative charge)] as it has been observed with Xenopus laevis SNBPs (Frehlick et al., 2007).

Interaction of C. variopedatus SNBPs with DNA.
Discussion
CvPL has similarity to arginine-rich C-terminal of chordate PL-types
Annelida and Mollusca derive from a common protostome ancestor. The Polychates, among the Annelida phylum classes, have many typical features of ancestral annelids. Then, the general opinion is that the worms are closer to the Mollusca phylum. The nuclei of the sperm of the marine bivalve mollusks contain a complex protein mixture in which PL-type and histone-like proteins coexist with a complete set of somatic histones (Ausió, 1986; Ausió et al., 1987). The nuclear protein composition consists of about 20% histones and about 80% sperm-specific protein component. The persistence of a small amount of the somatic-type histones seems to be a common feature of sperm of the class Bivalvia (Ausió, 1986) that contains only PL-type proteins. Evidence also exists for the coexistence of P-type proteins and histones in some mammals (Ausió et al., 2011), including humans (Hammoud et al., 2009). The sperm chromatin of C. variopedatus does not contain somatic-type histones, but only CvPL and a sperm histone H1 (De Petrocellis et al., 1983). However, the coexistence or not with remnant somatic histones at the end of spermiogenesis is not a good criterion for the classification of SNBPs; the CvPL is highly likely a PL-type SNBP. Moreover, the electrophoretic mobility in HrAU-PAGE of CvPL (see Fig. 2A) as well as its primary sequence (Fig. 2B) suggests that this protein shares features with PL-type proteins. In fact, the mobility of the CvPL is between those of PL-II and PL-III subtypes of bivalve mollusks.
There are basically two main subtypes of PLs: those whose structure consists of a winged-helix fold domain (WHD) (Ramakrishnan et al., 1993), like PL-I from the surf clam Spisula solidissima (mollusk) (Ausió and Subirana, 1982a; Lewis et al., 2004b), and those that lack this domain, like Mytilus (a bivalve mollusk) PL-III (Eirín-López et al., 2008). Although CvPL is certainly a PL-type protein, its high arginine content (see Table 1) reveals some common features with P-type proteins such as it happens in some chordate PLs (Ausió et al., 1997; Lewis et al., 2004a; Saperas et al., 2006). In fact, based on the compositional amino acid analysis, it was hypothesized by Subirana et al. (1973) that P-type had evolved from a primitive somatic-like histone precursor via a protamine-like intermediate through a mechanism of vertical evolution. This is supported by the observation that these organisms that replace histones with P-type in the mature sperm are always found at the furthermost tips of the evolutionary branches (Ausió, 1999), whereas sperm-specific germinal histones are found in the sperm of more primitive organisms such as the sponge Neofibularia (Ausió et al., 1997) and the sea urchin Paracentrotus lividus (De Petrocellis et al., 1980). Recent evidence has identified histone H1 as the putative precursor of protamines (Lewis et al., 2004a; Eirín-López and Ausió, 2009). A detailed systematic analysis of the distribution of SNBPs in fish provided additional support for the vertical evolution hypothesis of SNBPs by revealing that the sporadic distribution of P-type was not random and could be traced phylogenetically (Saperas et al., 1994). A recent publication has provided an important insight into potential molecular mechanisms by which specialized lysine-rich PLs of low-molecular weight could have evolved into the arginine-rich SNBPs of the P-type (Lewis et al., 2004a).
What is really surprising is the similarity observed between CvPL and the C-terminal of the arginine-rich PL of some chordates (Fig. 2B), both in sequence and in electrophoretic mobility (Fig. 2A, compare lanes 1, 6, and 8). The potential implications from this are that the lysine-to-arginine transition observed in the PL-types in deuterostomes (Eirín-López and Ausió, 2009) has also taken place in a similar fashion in protostomes likely as a result of a parallel evolution.
CvPL/CvH1 and the structure of the sperm chromatin
How can a histone H1 and a relatively short PL protein (Fig. 2A, lane 3) organize the sperm chromatin of C. variopedatus? The presence of only two proteins with a low secondary structure poses the problem as to how these can contribute to the organization of the chromatin in the sperm of C. variopedatus. Another important related question is: Are these SNBPs deposited on DNA sequentially or independently during spermiogenesis? A similar situation to that observed in C. variopedatus is observed in the mussel (Mytilus sp.) in which a histone H1-like (PL-II/PL-IV) coexists with a PL-III of smaller molecular weight (see Fig. 2A, lane 4). PL-III is encoded by its own gene (Ruiz-Lara et al., 1993; Eirín-López et al., 2006a), and it is sequentially incorporated onto the sperm chromatin after initial deposition PL-II/PL-IV during spermiogenesis (Bloch, 1966). In other instances, a single arginine-rich histone H1-like related protein (PL-I) consisting of the characteristic WHD is present in the sperm as is the case of the fish M. surmuletus (Fig. 2A, lane 8) (Saperas et al., 2006). This protein, in some instances, is post-translationally cleaved (Fig. 2B), presumably after deposition onto DNA into two fractions: one consisting of the WHD and another one corresponding to the highly arginine-rich C-terminal domain as in the tunicates from the genus Styela (Fig. 2A, lane 7) (Saperas et al., 1992). The need for the presence of an SNBP component with a WHD in all these organisms is not clear, but it is presumably the result of the mechanism involved in the displacement of histones by these SNBPs during spermiogenesis. The presence of a WHD may result from the need of targeting them to the DNA exit and entry sites of the nucleosomes whose histones are being displaced in order to maintain the DNA superhelicity (Ausió, 1995). Indirect evidence for this comes from the experimentally observed chromatin organization in the sperm of the PL-containing fish M. surmuletus (Saperas et al., 2006). Therefore, it is likely that C. variopedatus SNBPs, consisting of an H1 and a PL protein, organize the sperm chromatin in a similar fashion as other PL-containing sperm. The high similarity of CvPL to the C-terminal of M. surmuletus PL-I comes in support of this notion. Furthermore, both CvH1 and CvPL bind to DNA in a way that is very similar to that of vertebrate SNBPs (Frehlick et al., 2007) and very different from that of somatic H1 histone (Fig. 4) (Piscopo et al., 2010). The reasons for this observed difference could reflect the higher ability of arginine versus lysine (or their low K/R ratio [Piscopo et al., 2010], see Table 1) to form hydrogen bonding with the DNA (Ausió, 1995) in addition to the electrostatic interactions (Ichimura et al., 1982; Vila et al., 2001), achieving a more efficient chromatin packing favored by the sperm of highly evolved organisms with internal fertilization (Kasinsky et al., 1985) in both the protostome and deuterostome branches. Another contributing factor could be the observed potential of CvPL that could also be able to form aggregated structures. Indeed, we have reported that the oligomeric state of CvPL depends on the presence of both lysine amino groups and arginine guanidine groups (Piscopo et al., 1993) that also determine the DNA-binding mode (Piscopo et al., 2010).
The answer to the two questions posed at the beginning of this section will ultimately depend on whether CvH1 and CvPL share a precursor relationship (Fig. 5A) as that observed in tunicates or whether they are the product of two independent genes as in the case of PL-II/PL-IV and PL-III in Mytilus.
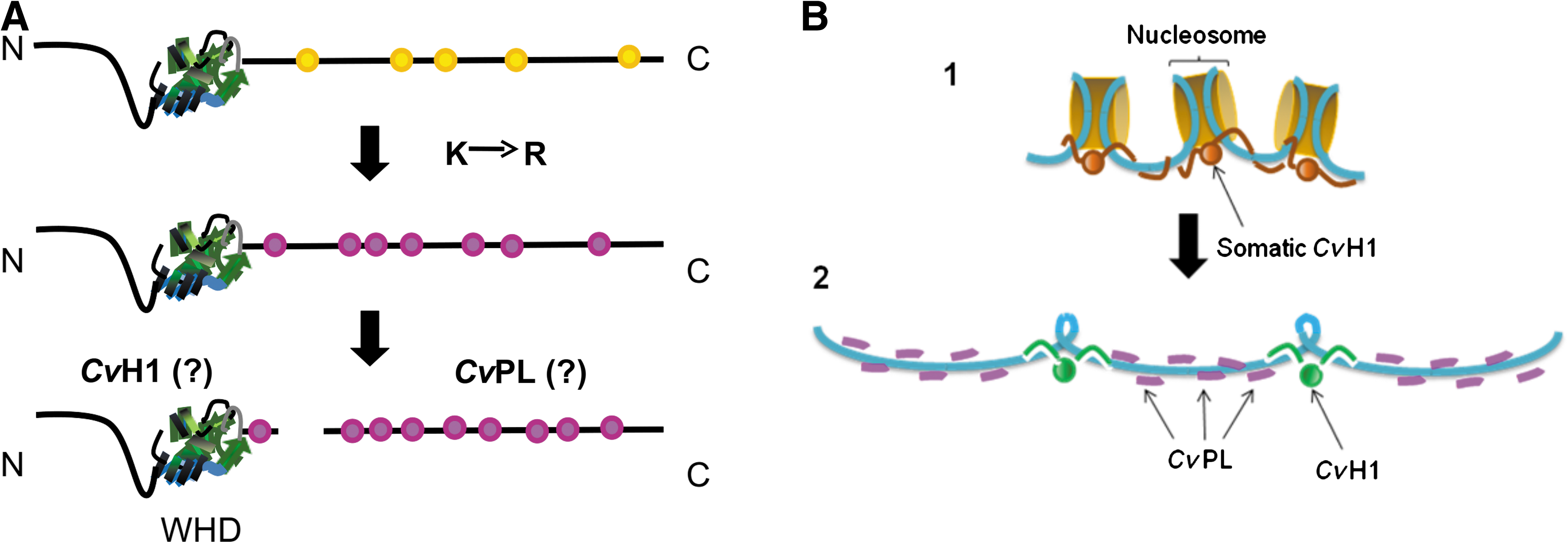
Two hypothetical models to explain the relationship between CvPl and CvH1 and their chromatin deposition.
In the first scenario (Fig. 5A), CvH1 and CvPL will be the products of the protein precursor after a putative post-translational cleavage as it has been observed for the PL-II and PL-IV in the mussel Mytilus (Carlos et al., 1993) or for P1 and P2 in the tunicates of the genus Styela (Fig. 2A, lane 7) (Lewis et al., 2004a). It is important to notice that in this latter group, a transition leading from the characteristic lysine-rich nature of the histone H1 C-terminal tail to arginine was observed. Such transition has been ascribed to one of the potential molecular mechanisms involved in the transition from PL- to P-types starting from a primitive H1 histone precursor (Lewis et al., 2004a). Given the aforementioned similarities between the C-terminal tails of vertebrate PLs (Fig. 2B) and those of CvPL, it is possible that a similar mechanism may have occurred in lower invertebrates.
Alternatively (Fig. 5B), CvH1 and CvPL could be the result of two independent genes related to an ancestral H1 histone gene as it is the case of PL-II/PL-IV and PL-III of Mytilus (Fig. 2A, lane 4) (Erín-López et al., 2006a). In an attempt to elucidate whether this could be the case, the molar ratio between these two proteins, as they are obtained from acid extraction of the sperm chromatin, was determined. The analysis indicated that there are ∼6 molecules of CvPL per each molecule of CvH1. This result clearly supports this second alternative and suggests a sperm chromatin organization pattern such as the one shown in Fig. 5B. In this model, the interaction of these proteins with DNA will be mainly electrostatic as supported by their in vitro interaction analysis (Fig. 4). Taking into consideration their molecular mass as well as their amino acid composition (De Petrocellis et al., 1983), it is possible to estimate that such arrangement would correspond to ∼340 positively charged amino acids for every nucleosome. Assuming an average value of 200 bp of DNA per nucleosome in the somatic chromatin at the onset of spermiogenesis, this corresponds to a value of 85% charge neutralization, a value that is very close to that observed for other P- and PL-type sperm chromatin.
The sequence of the genes for CvPL and CvH1 and how these two proteins are sequentially deposited onto DNA during the replacement of somatic histones are the objectives of future research in our lab.
Conclusion
We report evidence of a peculiar sperm protein from the annelid C. variopedatus. The electrophoretic mobility in HrAU-PAGE of CvPL as well as its primary structure suggests that this protein shares features with PL-type proteins. Because of this evidence, we could classify CvPL as a PL-type protein. However, its high arginine content led us to suppose that this protein shares some common features with P-type proteins, such as it happens in some chordate PLs.
It is now well established that P-type proteins had evolved from a primitive somatic-like histone precursor via a PL-type intermediate through a mechanism of vertical evolution. The similarity observed between CvPL and the C-terminal of the arginine-rich PL proteins of some chordates, both in sequence and in electrophoretic mobility, is the possible evolutionary evidence that the lysine-to-arginine transition observed in the PL-type in deuterostomes has also taken place in a similar way in protostomes as a result of parallel evolution.
Both CvH1 and CvPL bind to DNA in a way that is very similar to that of vertebrate SNBPs and different from that of somatic histones H1 (Fig. 4). The reasons for this observed difference could reflect the higher ability of arginine versus lysine to form hydrogen bonding with the DNA, in addition to the electrostatic interactions. In addition, a particular chromatin organization must be present in the sperm of C. variopedatus that is not found in other organisms. A histone H1 and a relatively short PL protein are involved (Fig. 5). It is likely that C. variopedatus SNBPs organize the sperm chromatin in a similar fashion as other PL-containing sperm (Lewis and Ausió, 2002).
Footnotes
Acknowledgments
We thank Dr. Maria Malanga, prematurely passed away, in 2011 November, for providing the peptides used in the Tricine SDS-PAGE as a molecular weight marker.
Disclosure Statement
No competing financial interests exist.