
Abstract
Directing differentiation of neural stem/progenitor cells (NPCs) to produce functional neurons is a promising remedy for neural pathological conditions. The major challenge, however, lies in the effective and efficient generation of a sizable population of neurons. A potential strategy is to incorporate RNA interference (RNAi) during directed stem cell differentiation to recapitulate the complex cell-signaling cascades that often occurs during the process. In this study, in vitro silencing of RE1-silencing transcription factor (REST) was carried out using small-interfering RNAs (siRNAs) to evaluate the efficacy of combining REST knockdown with conventional differentiation approaches to enhance neurogenesis. While earlier studies have demonstrated enhanced neuronal lineage commitment from embryonic stem cells and mesenchymal stem cells upon REST knockdown, the effects of REST silencing during other stages of neural development have not been extensively evaluated. We hypothesize that REST knockdown would enhance NPC development to mature neurons and that induced REST silencing can serve as a potential biochemical approach to direct cell fate. Under nonspecific induction conditions, REST knockdown induced eightfold higher Tuj1 mRNA expression at day 14 compared with untransfected cells and cells subjected to scrambled-siRNA treatment (controls). Immunostaining also revealed greater percentage of Tuj1 positive cells with REST knockdown. Combined with neuronal induction, REST silencing enhanced the kinetics of neuronal differentiation and the rate of maturation of committed neuronal cells. Specifically, upregulation of MAP2 occurred as early as 3 days after induction with REST silencing and the expression was comparable to the controls at day 14. Likewise, downregulation of REST generated more than twice the percentage of Tuj1 and MAP2 positive cells compared with controls at day 5 (p<0.05). Morphologically, REST-silencing enhanced the number and length of neurite extensions from Tuj1 positive cells (p<0.05), which was not evaluated in previous differentiation studies with REST knockdown. Taken together, these results demonstrate the efficacy of combining REST silencing during directed NPC differentiation to enhance the rate of differentiation and subsequent maturation of NPCs. This study also highlights the potential of RNAi as a biomedical strategy for guided stem cell differentiation.
Introduction
RNAi is a post-transcriptional gene silencing technique which has been extensively employed in studies to address fundamental biological questions, while emerging in recent years as a potential therapeutic treatment for cancer (Dharmapuri et al., 2011) and various human diseases (Singer et al., 2005; Hong et al., 2006; Davidson and McCray, 2011). The basic premise underlying such broad utility of RNAi lies in its capacity to virtually target any gene of interest. Although current applications in stem cells remain largely restricted to studies on molecular pathways and signaling, RNAi can be expanded as a biomedical strategy to direct lineage-specific differentiation of stem cells for therapeutic purposes. One key factor that can possibly be adapted into the RNAi strategy for directing neuronal differentiation of neural stem cells is the repressor element 1 (RE-1) silencing transcription factor (REST). In most differentiated non-neuronal cells and uncommitted neural stem cells, REST functions as a transcriptional repressor for a myriad of neuronal specific genes such as ion channels, synaptic vesicles proteins, and neurotransmitter receptors (Chong et al., 1995; Schoenherr and Anderson, 1995), by binding to a highly conserved DNA sequence known as RE-1. During neurogenesis, REST is rapidly downregulated in embryonic stem cells (ESCs) and neural stem cells upon differentiation into neurons (Sun et al., 2005; Gao et al., 2011). Failure of downregulation due to ectopic expression of REST resulted in axon path finding errors and blocked the developmental acquisition of sodium channel excitability in neuronal cell lines (Paquette et al., 2000). Conversely, induced downregulation of REST has been shown to promote neuronal commitment in ESCs and mesenchymal stem cells (MSCs). Specifically, the knockdown of REST in ESCs induced neural progenitors formation (Kumar Gupta et al., 2009) and when applied to MSCs, cross-lineage differentiation to neurons was observed (Yang et al., 2008). Although these results suggest that downregulation of REST plays an important role in embryonic neural development, detailed analysis on the effect of REST knockdown at other stages of neural commitment remains limited. In particular, the effects of REST silencing on NPC neuronal differentiation have not been evaluated.
We hypothesize that REST plays an important role in neuronal development even at the neural progenitor stage and that by deliberately silencing the expression of REST during directed NPC differentiation, it may be possible to induce greater and more rapid differentiation of NPCs into neurons. Therefore, in this study, primary mouse NPCs were transfected with REST small-interfering RNAs (siRNAs), and the efficacy of REST knockdown in directing NPCs neuronal differentiation was evaluated under nonspecific (serum-containing) and neuronal instructive (serum-free) culture conditions. It has been well established that the presence of serum tends to facilitate astrocytic differentiation at the expense of neuronal (Bartlett et al., 1998) and oligodendrocytic (Raff et al., 1983) differentiation. Accordingly, the feasibility of REST knockdown in promoting neuronal differentiation was examined for such largely dissimilar induction media to ensure its applicability across varied differentiation conditions. We aim to provide a detailed kinetic analysis on induced differentiation in NPCs and to establish a platform for quantifying the extent of accelerated neuronal differentiation with REST knockdown under these two conditions. Such evaluation of REST knockdown efficacy will provide useful information for translating RNAi in stem cells via REST silencing as a potential biomedical approach for directing stem cell differentiation for therapeutic purposes.
Materials and Methods
Materials
Fibroblast growth factor-2 (FGF-2) was purchased from R&D systems. Epidermal growth factor (EGF) and heparin was purchased from Merck. About 0.1% gelatin was purchased from Stemcell technologies. Penicillin-streptomycin and neurobasal medium were purchased from Gibco. Fetal bovine serum (FBS) was purchased from Hyclone. REST siRNA was purchased from Ambion (ID #: s72890) while scrambled, nontargeting siRNA was purchased from 1st Base. M-MLV Reverse transcriptase (M3681) was purchased from Promega. Laemmli sample buffer and iQ SYBR Green Supermix were purchased from Bio-Rad. BCA assay kit was purchased from Pierce. Retinoic acid, radio immunoprecipitation assay (RIPA) buffer, 10% formalin, tris-buffered saline (TBS), Triton X-100, fluoromount, sucrose, bovine serum albumin (BSA), mouse anti-microtubule associated protein-2 (MAP2), and fluorescein isothiocyanate (FITC) conjugated goat anti-rabbit antibody were purchased from Sigma-Aldrich. Rabbit anti-REST, goat anti-nestin, and mouse anti-β Actin were purchased from Santa Cruz. Mouse anti-glial fibrillary acidic protein (GFAP) and mouse anti-O4 were purchased from Millipore. Rabbit anti-βIII tubulin (Tuj1) was purchased from Covance. HRP-conjugated anti-rabbit IgG and HRP-conjugated anti-mouse IgG were purchased from GE Healthcare. All other reagents were purchased from Invitrogen.
Primary mouse NPCs isolation
All experiments were approved by the Institutional Animal Care and Use Committee (IACUC) and animals were treated in accordance with the IACUC guidelines in an Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) approved animal facility. NPC isolation was performed according to an established protocol with some modifications (Bull and Bartlett, 2005; Peltier et al., 2010). Briefly, six adult 5 weeks old C57/BL6 mice were killed by cervical dislocation and their brains immediately removed. Then the tissues were dissected using two pairs of forceps. First, the midbrain was removed, and then the hippocampus, as a whole, was gently separated from the corpus collosum. The tissue samples were diced with a scalpel blade in Hanks' Balanced Salt Solution (HBSS) and centrifuged at 2000 rpm for 5 min. After removing the supernatant, tissue samples were enzymatically digested with 2 mL of Papain (Worthington) containing DNase I 250 U/mL and Dispase II 1 U/mL at 37° for 30 min. Tissues were then dissociated into single cells by titration and centrifuged at 2000 rpm for 5 min. The pellet was resuspended into 20 mL of 0.9 M sucrose in 0.5× HBSS and centrifuged at 2000 rpm for 10 min. Then, cells were resuspended into 2 mL of culture media composed by Dulbecco's Modified Eagle Medium (DMEM)/nutrient mixture F-12 (F12) supplemented by 1% N2 supplement, 1% penicillin/streptomycin and 1 mM
Cell culture
NPCs isolated from the hippocampi of 5 weeks old mice were used in this study. The murine NPCs are homogeneously dividing progenitor cells which are immunoreactive to nestin, vimentin, and brain lipid binding protein (BLBP). Cells were cultured and expanded in 0.1% gelatin coated tissue culture flask with complete proliferative medium consisting of DMEM/F12, 1% N2 supplement, 1%
REST siRNA transfection and NPC differentiation
One day before transfection, NPCs were trypsinized and plated onto laminin-coated 24-well plate or coverslips at a density of 4.0×104 cells/well in 500 μL of complete proliferative medium. After 24 h incubation, cells were approximately 40%–50% confluent and Lipofectamine RNAimax was used for the transfection of NPCs according to the manufacturer's protocol. Briefly, 1.0 μL of Lipofectamine RNAimax was diluted in 50 μL of complete proliferative medium and siRNA was separately diluted to a final concentration of 40 nM in another 50 μL of complete proliferative medium. Thereafter, the two solutions were mixed with gentle pipetting and incubated for 15 min at room temperature to allow for complex formation. Finally, 100 μL of siRNA-Lipofectamine RNAimax complexes was added drop by drop to the pre-existing media so that the final transfection volume was 600 μL/well and the cells were transfected for 24 h. In our preliminary experiments, three different strands of REST siRNAs were evaluated for gene knockdown and the siRNA with highest silencing efficiency was used for all subsequent experiments. The day of transfection was denoted as day 1 and the cells were induced to differentiate at day 0 when the medium was replaced with either mixed or neuronal induction medium. For mixed lineage differentiation, a second dose of siRNAs was administered 7 days after initial transfection to maintain the silencing effect. For neuronal differentiation, a booster of siRNA was added at day 0 to ensure effective silencing of the target mRNA at the very early stage of differentiation.
Three sets of differentiation conditions were examined: NPCs differentiating in the presence of REST siRNA (denoted as siREST); nontargeting, negative siRNA (denoted as siNEG); and in the absence of siRNA treatment (denoted as induction). For induction samples, 1.0 μL of Lipofectamine RNAimax diluted in 100 μL of complete proliferative medium was added into each well to normalize any gene expression differences that may result due to Lipofectamine RNAimax treatment. Cells were then harvested at different time points for real-time polymerase chain reaction (PCR) or immunostaining to evaluate the REST expression profile or neural marker expression. The entire experiment was repeated at least three times.
Real-time PCR
At specific time points, cells were lysed using TRIzol® reagent. For each sample, three wells of cells from a 24-well plate were pooled together in order to obtain sufficient RNA for analyses. Reverse transcription was carried out using M-MLV Reverse transcriptase according to the manufacturer's protocol. Following which, 1 μL of reaction mixture was taken as cDNA template for amplification and real-time PCR analysis was performed using iQ SYBR Green Supermix in a StepOnePlus™ Real-time PCR system (Applied Biosystems), with β-actin as the housekeeping gene. The sequences of the primers used are listed in Table 1. The real-time PCR cycling condition used was: 10 min at 95°C, 40 cycles at 95°C for 15 s, followed by 60°C for 1 min. Our preliminary studies showed that these primers had similar amplification efficiency under the parameters used. Therefore, the ΔΔCT method was used for fold change analysis. Unless otherwise stated, all results were normalized with respect to the mRNA expression of undifferentiated NPCs.
Western blot
Total soluble protein was extracted by RIPA buffer and protein concentrations were measured with BCA assay kit according to manufacturer's protocol. After mixing with Laemmli sample buffer supplemented with beta-mercaptoethanol, the samples were denatured by boiling for 10 min. Thereafter, 10 μg of protein was loaded into each well of an sodium dodecyl sulfate (SDS) polyacrylamide slab gel (separating gel: 10% acrylamide/bis-acrylamide [30%/0.8% w/v], 390 mM Tris-HCl, pH 8.8, and 0.1% SDS; stacking gel: 4% acrylamide/bis-acrylamide, 82.5 mM Tris-HCl, pH 6.8, and 0.2% SDS). The protein samples were then resolved in a Mini-PROTEAN 3 electrophoresis unit (Bio-Rad) for 1 h at 120 V. Following that, the resolved proteins were transferred onto a nitrocellulose membrane using a Trans-Blot Semi-Dry Transfer Cell (Bio-Rad) at 15 V for 1 h. After overnight blocking with 5% BSA in TBS, the membrane was washed three times in TBS with Tween-20 (TBST). Primary antibodies (rabbit anti-REST or mouse anti-beta-actin [ACTB]) diluted 1:200 in 5% BSA was then added for 2 h incubation with orbital shaking. After three washes in TBST, the membrane was incubated with secondary antibodies (HRP-conjugated anti-rabbit IgG, HRP-conjugated anti-mouse IgG, diluted 1:6000) in TBST for another 1 h. Finally, chemiluminescence signal produced by ECL Plus™ Western Blotting Detection Reagents (GE Healthcare) was detected and developed on an imaging film (Kodak). For densitometric analysis of scanned blots, ImageJ software (NIH) was used and all results were normalized with respect to the REST/β-Actin intensity ratio of undifferentiated NPCs.
Immunofluorescence staining
For immunofluorescence staining of nestin, Tuj1, MAP2, and GFAP, samples were fixed in 10% formalin, permeablized with 0.05% Triton X-100 in phosphate-buffered saline, and then blocked overnight with blocking buffer containing 3% BSA and 5% FBS at 4°C. Samples were then sequentially incubated with primary antibodies for 2 h and secondary antibodies for 1 h at room temperature. For oligodendrocyte marker, O4, samples were fixed and blocked as indicated above without permeabilization in Triton X-100 to prevent the antigen from dissolving out of the cellular membranes. Thereafter, samples were sequentially incubated with primary antibody overnight at 4°C and secondary antibody for 1 h at room temperature. The primary antibodies used were as follows: mouse anti-GFAP (1:500), rabbit anti-Tuj1 (1:2000), mouse anti-MAP2 (1:500), goat anti-nestin (1:500), and mouse anti-O4 (1:500). The secondary antibodies used were as follows: Alexa-Fluor 633 goat anti-mouse antibody (1:500), Alexa-Fluor 555 donkey anti-mouse antibody (1:500), FITC conjugated goat anti-rabbit antibody (1:400), Alexa-Fluor 647 chicken anti-rabbit antibody (1:500), and Alexa-Fluor 488 donkey anti-goat antibody (1:500). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and samples were then mounted on glass slides using fluoromount and imaged by confocal microscopy (Zeiss; LSM 710 Meta Laser Scanning Confocal Microscope).
Quantitative analysis of immunofluorescence staining results
To determine the proportion of differentiated cells in each sample, images were captured at 200× magnification from at least five randomly selected areas. Thereafter, ImageJ software (NIH) was used to compute the total number of cells (DAPI stained nuclei) and the number of cells expressing Tuj1, GFAP, O4, MAP2, or nestin. At least 1000 cells were counted for each sample.
Neurite outgrowth was characterized by counting the number of neurite extensions and measuring the length of the longest neurite for each Tuj1 positive cell. Thereafter, the number of cells exhibiting less than two or greater than three neurites, and the number of cells with neurite length greater than five times or eight times the cell body length were expressed as a percentage over the total number of Tuj1 positive cells in each sample. At least 50 Tuj1 positive cells per sample were included in the measurement.
Statistical analyses
All data were expressed as the mean±standard error of the mean from three individual experiments. Statistical analyses were carried out using One-way analysis of variance and Tukey post-hoc tests after verifying equal variances. The student's t-test was used for statistical comparisons involving two samples.
Results
Effect of REST knockdown on neural marker expression during mixed lineage differentiation
NPCs with or without REST knockdown were cultured in mixed lineage induction medium, which nonspecifically supports the differentiation of both neurons and glial cells. The efficacy of REST silencing under mixed lineage differentiation conditions was assessed by quantitative PCR analysis and western blotting. Significant knockdown of REST mRNA was observed in siREST compared with siNEG controls on days 3 and 5 (Fig. 1A, 53% silencing on day 3; 35% silencing on day 5). Western blot analysis also showed a similar trend where downregulation of REST protein in siREST was observed at days 3, 5, and 7 compared to siNEG and induction samples (controls) (Fig. 1B, C). These results indicated the successful silencing of REST at both protein and transcript levels in siREST samples within the first week of differentiation. Significantly lower REST protein levels was also observed in siREST, siNEG, and induction samples at day 7 compared with that in day 3, indicating endogenous downregulation of REST protein as a result of the induction medium (Fig. 1C). Although the REST mRNA expression across all samples may similarly be reduced as a result of differentiation, the gradual increase of REST transcript level relative to the siNEG controls prompted a second dose of siRNAs at day 7 to maintain the silencing effect for subsequent differentiation studies.
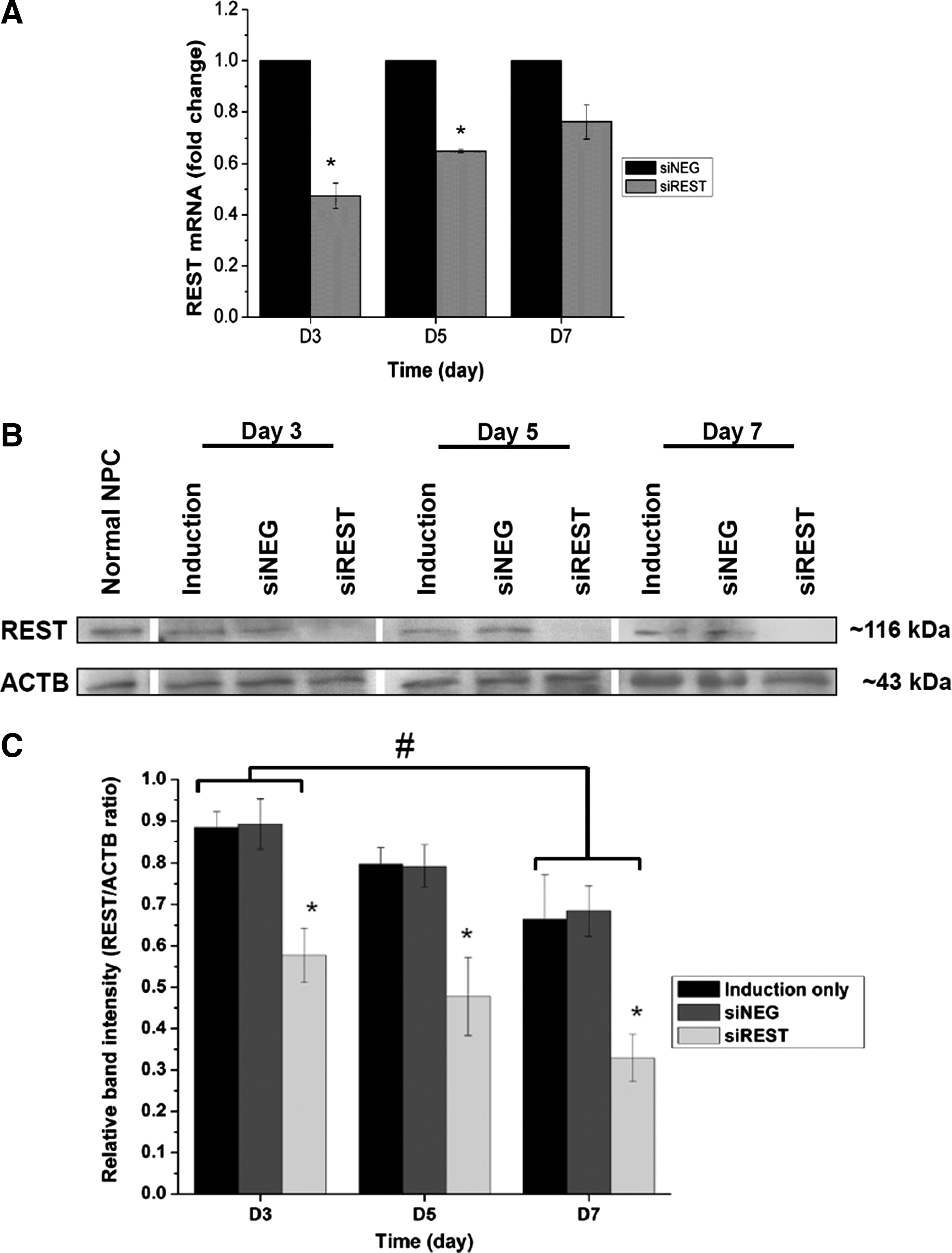
Rest knockdown efficiency in mixed lineage induction medium.
The effects of REST knockdown on NPCs differentiation in mixed lineage induction medium was evaluated with quantitative PCR of several neural markers. Nestin, a marker for NPCs was downregulated with respect to undifferentiated NPCs and the expression levels remained fairly similar across all samples during the 14 days of mixed lineage differentiation (Fig. 2A). Two neuronal markers were tested. Tuj1, a phenotypic marker for early neuronal cell, was significantly upregulated in siREST at days 5 and 14 (Fig. 2B). In particular, REST knockdown induced eightfold higher Tuj1 mRNA expression at day 14 relative to the controls (p<0.05). The expression of mature neuronal marker, MAP2, remained low and no significant difference was observed between the samples for all time points (Fig. 2C). GFAP, a marker for astrocytes, was enhanced overall but was not significantly affected by REST silencing (Fig. 2D). Meanwhile, the oligodendrocyte marker, RIP, was significantly upregulated by REST knockdown compared with the controls at day 14 (p<0.05).
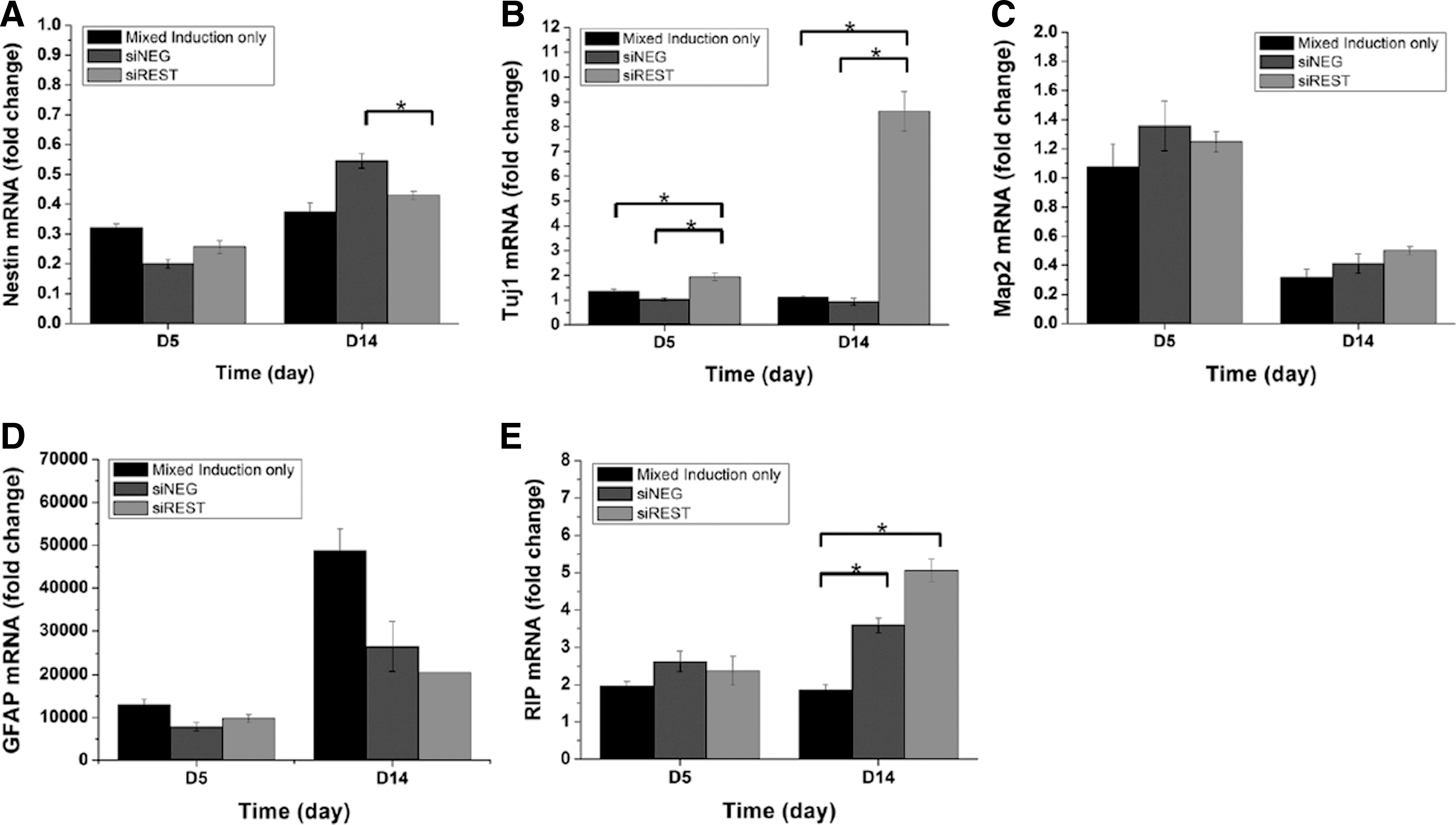
Real-time PCR analysis of neural marker expression by NPCs differentiated in mixed lineage induction medium. Expression of
The relative populations of neuronal and glial cells were evaluated by immunofluorescence staining. Statistical analysis of neural markers expression (Fig. 3) revealed similar trends as real-time PCR results. At day 5, the percentage of nestin positive cells was 39.2%, 34.9%, and 36.4% while the percentage of GFAP positive cells was 24.7%, 22.2%, and 21.1% for induction, siNEG and siREST respectively (Fig. 3B). At day 14, the percentage of nestin positive cells was 24.0%, 20.3%, and 23.1% while that for GFAP positive cells was 25.2%, 23.7%, and 21.8% for induction, siNEG and siREST respectively (Fig. 3C). The percentage of nestin positive and GFAP positive cells showed no significant difference between samples at both days 5 and 14. Similarly, the percentage of O4 positive oligodendrocytes was not significantly different even though a higher percentage was observed in siREST at day 14 (13.1% vs. 5.2% and 9.2% in induction and siNEG respectively). In comparison, the percentage of Tuj1 positive cells was significantly higher in samples with REST knockdown (Fig. 3D, p<0.01). Specifically, the percentage of Tuj1 positive cells was 10-fold higher in siREST at day 5 and 2.2-fold to 3.5-fold higher when compared with siNEG and induction at day 14 respectively. In addition, more Tuj1 positive cells were also generated in siREST at day 5 relative to the controls at day 14 (6.3% vs. 2.1% and 3.3% in the latter, p<0.01).
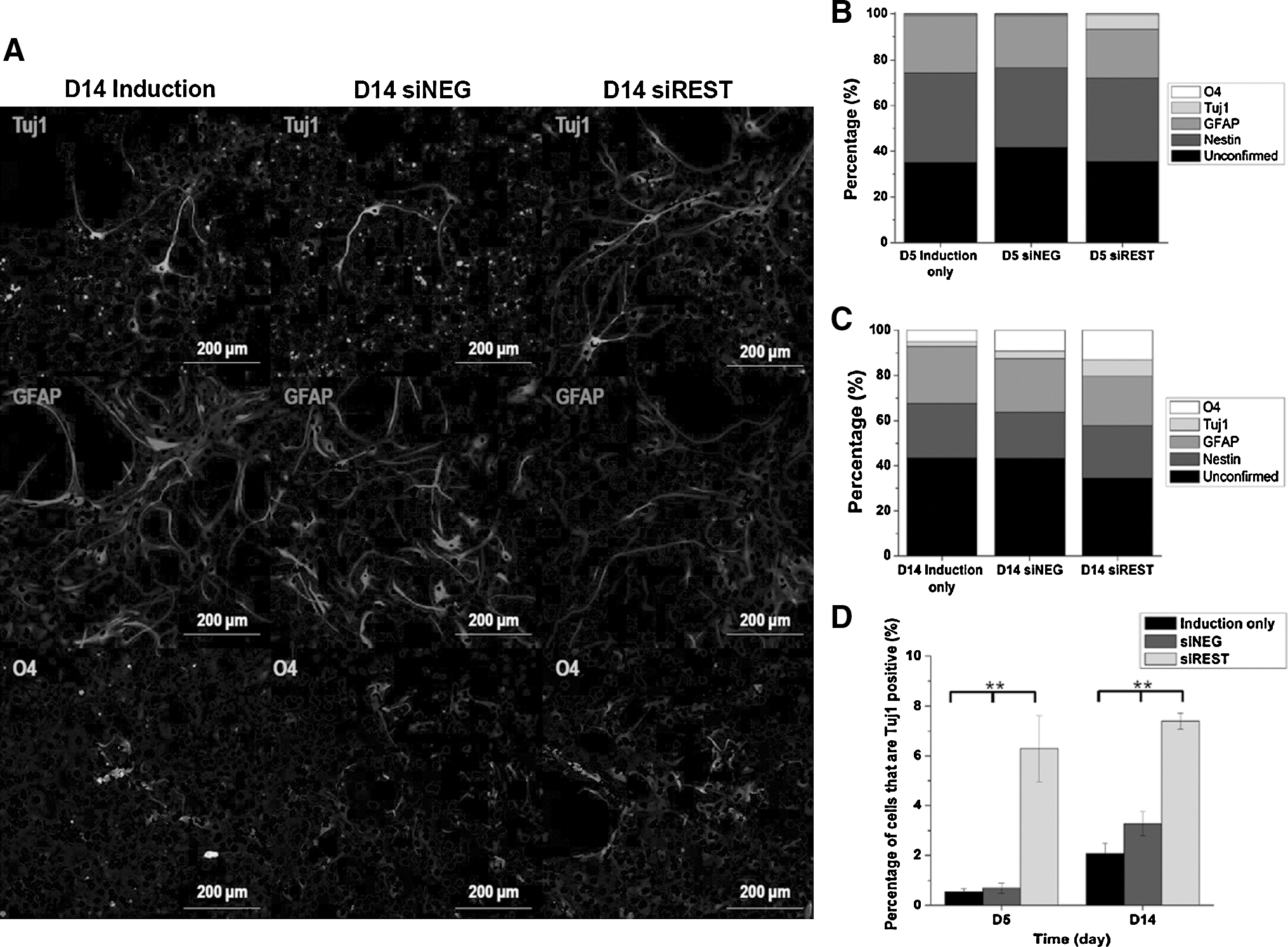
Immunofluorescence analysis of neural marker expression in NPCs differentiated in mixed lineage induction medium.
Morphologically, Tuj1 positive cells in siREST exhibited longer and more complex neurite outgrowth compared with the controls (Fig. 3A). As indicated in Figure 4A, the percentage of Tuj1 positive cells having three or more neurites was 10.0% in siREST while that in induction and siNEG were approximately two times lower (p<0.01). In addition, the percentage of Tuj1 positive cells that developed neurites greater than eight times body length was also larger in siREST (p<0.06, Fig. 4b). These results show that REST knockdown effectively induced early neuronal marker expression in NPCs and led to a shift toward the neuronal lineage during nonspecific mixed-lineage induction.
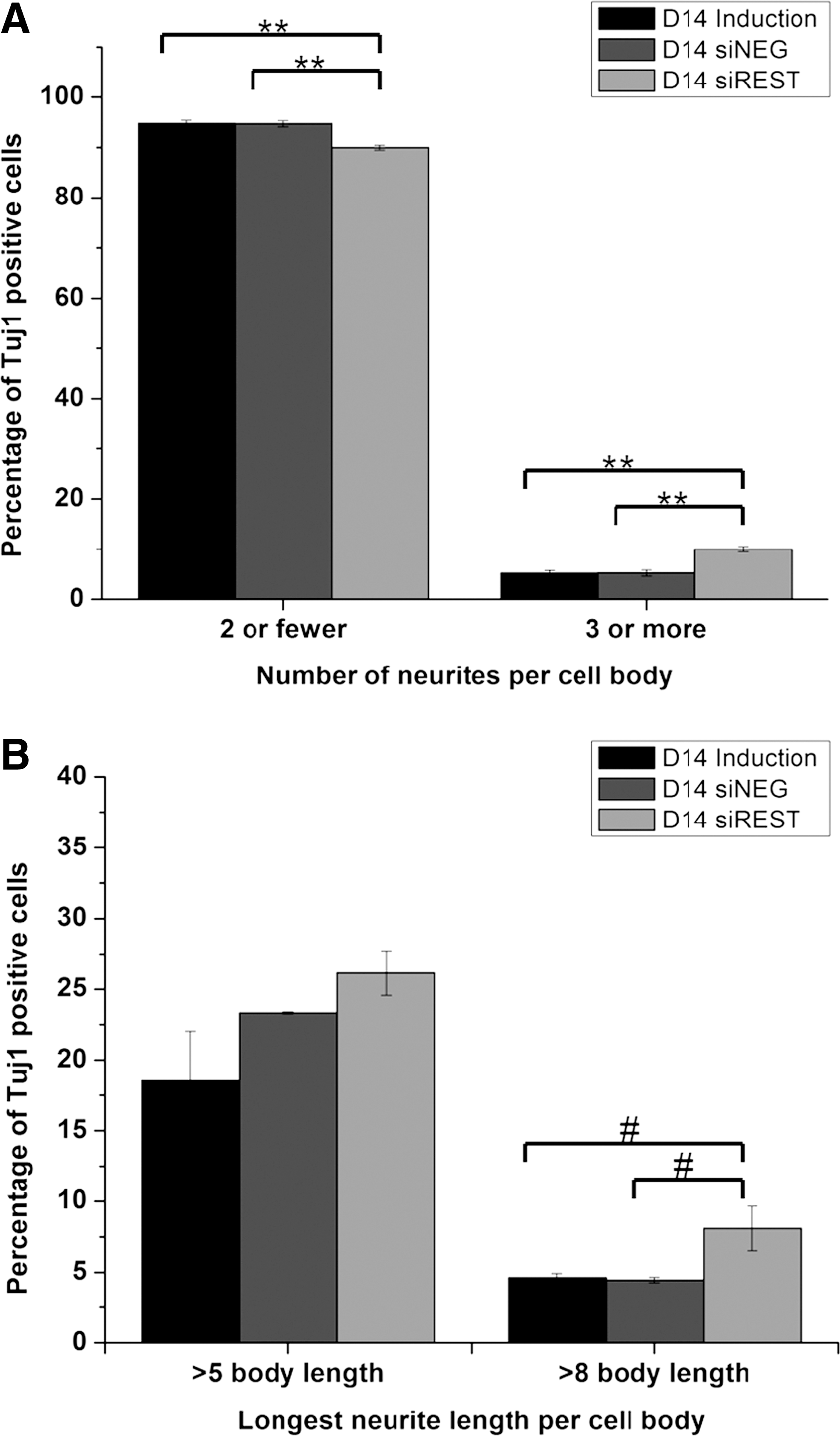
Quantification of neurite length and neurite number after 14 days of mixed lineage induction.
Effect of REST knockdown on neural marker expression with neuronal induction medium
The effect of REST knockdown in combination with stringent neuronal differentiation was then investigated. The neuronal induction medium specifically supports the differentiation of NPCs to neurons. REST silencing efficiency in the presence of neuronal induction medium was assessed by real-time PCR and western blot analysis as shown in Figure 5. The silencing profile of REST mRNA by real-time PCR analysis showed that when compared to scrambled siRNA treated samples, REST transcript level dropped by 35% on day 0 (1 day after transfection) but returned to 20% on day 1 (Fig. 5A). In an attempt to increase the silencing efficiency, as advocated by Ma et al. (2010), a repeated siRNA transfection was performed again at day 0. Using this protocol, the efficiency of REST silencing reached ∼50% at day 1 before gradually returning to 20% by day 3 (Fig. 5B). Therefore, a second booster of siRNA was applied at day 0 for all subsequent differentiation experiments. A similar trend was reflected by the western blot analysis of REST protein. In untransfected or scrambled siRNA-transfected control samples, REST protein levels gradually dropped with neuronal induction from day 1 to day 5. After REST siRNA transfection, REST protein expression decreased on day 1 before returning to a similar level as the controls by day 5 (Fig. 5C, D).
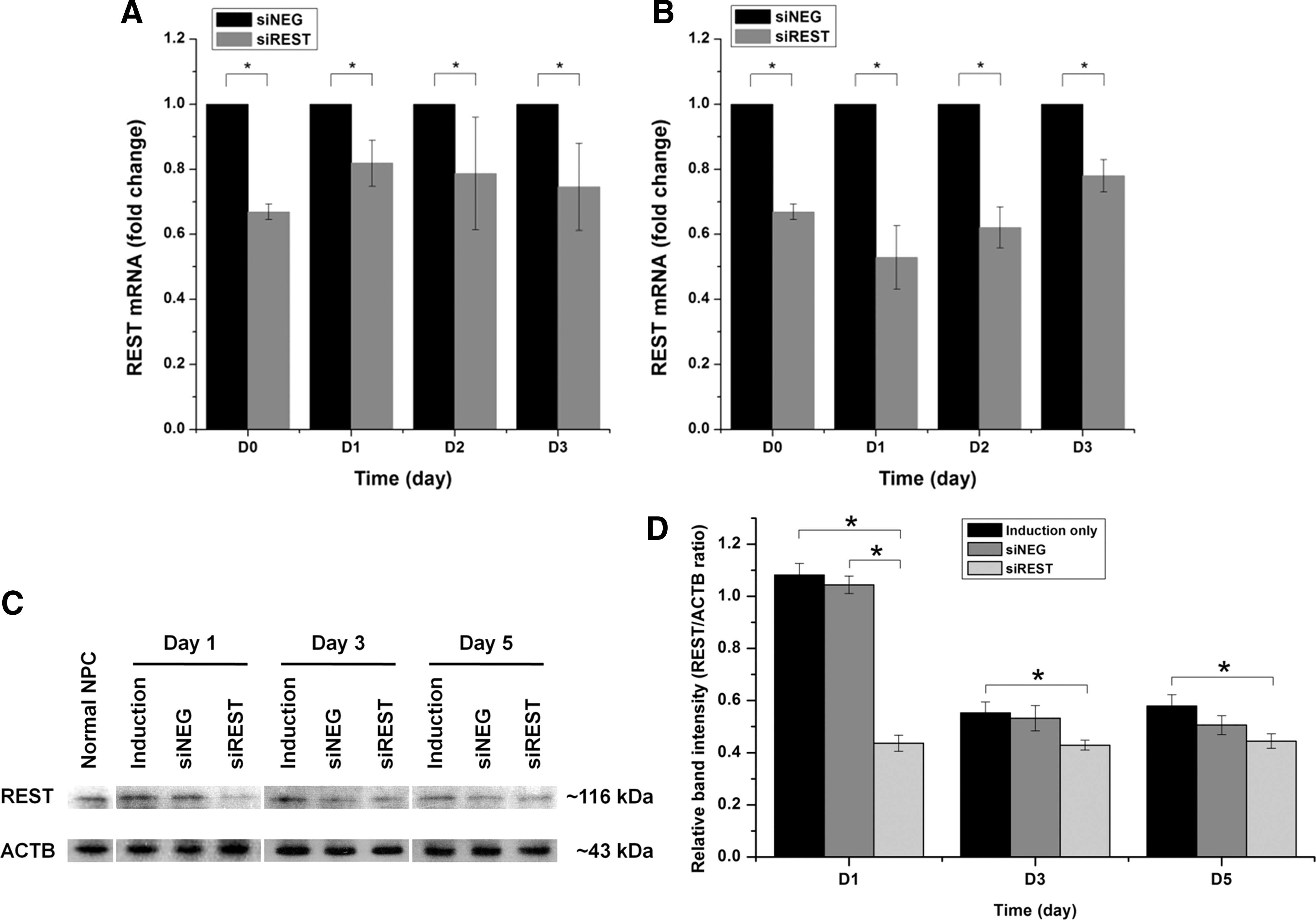
Rest knockdown efficiency in neuronal induction medium.
Figure 6A and B show the mRNA expression of neuronal markers, Tuj1 and MAP2 across 2 weeks of differentiation as evaluated by real-time PCR. Compared with undifferentiated NPCs, Tuj1 and MAP2 exhibited a general increase in their transcript levels with neuronal induction. With the suppression of REST, the rate of Tuj1 and MAP2 induction were further increased. In particular, the induction of Tuj1 began as early as day 0, that is, 1 day after REST siRNA transfection. By day 2, the level of Tuj1 in siREST was comparable to that of NPC under neuronal differentiation for 7 days. At all time points, siREST expressed higher levels of Tuj1 than the control groups. MAP2 was upregulated at day 3 with REST knockdown. At this time point, the level of MAP2 was similar to that of NPC under neuronal differentiation for 10 days. The greatest difference between siREST and the control groups was observed by days 3 to 7, and the difference was gradually reduced toward day 14.

Real-time PCR analysis of neural marker expression by NPCs differentiated in neuronal induction medium. Expression of
The protein expression pattern of Tuj1 and MAP2 agreed with the mRNA expression results as shown in Figures 7 and 8. At day 5, the percentage of Tuj1 (Fig. 8C) and MAP2 (Fig. 8D) positive cells were more than twice in siREST compared with siNEG and induction. However, all sample groups had approximately 80% Tuj1 or MAP2 positive cells by day 14. The percentages of glial cells were also estimated by GFAP and O4 staining. As indicated in Figure 7, REST-silencing had no obvious effect on the differentiation of NPCs into glial cells. The percentages of GFAP-positive cells were 3.5%, 4.1%, and 3.3% at day 5 (Fig. 7C), and 1.2%, 3.0%, and 0.9% at day 14 (Fig. 7D) for induction, siNEG and siREST respectively with no significant difference between sample groups. In addition, no O4 signal was observed for all groups after neuronal differentiation (data not shown).
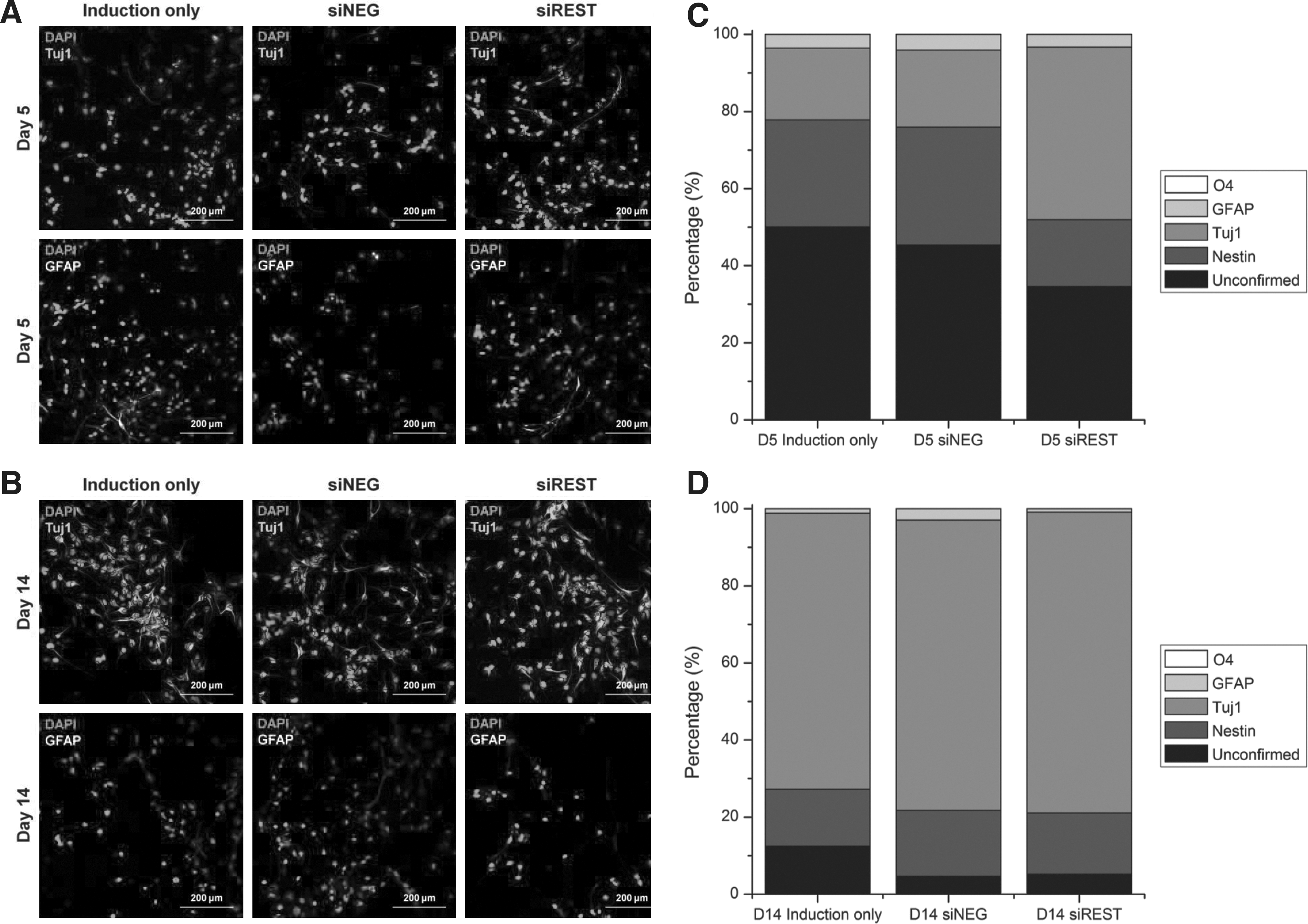
Immunofluorescence analysis of neural marker expression in NPCs differentiated in neuronal induction medium.
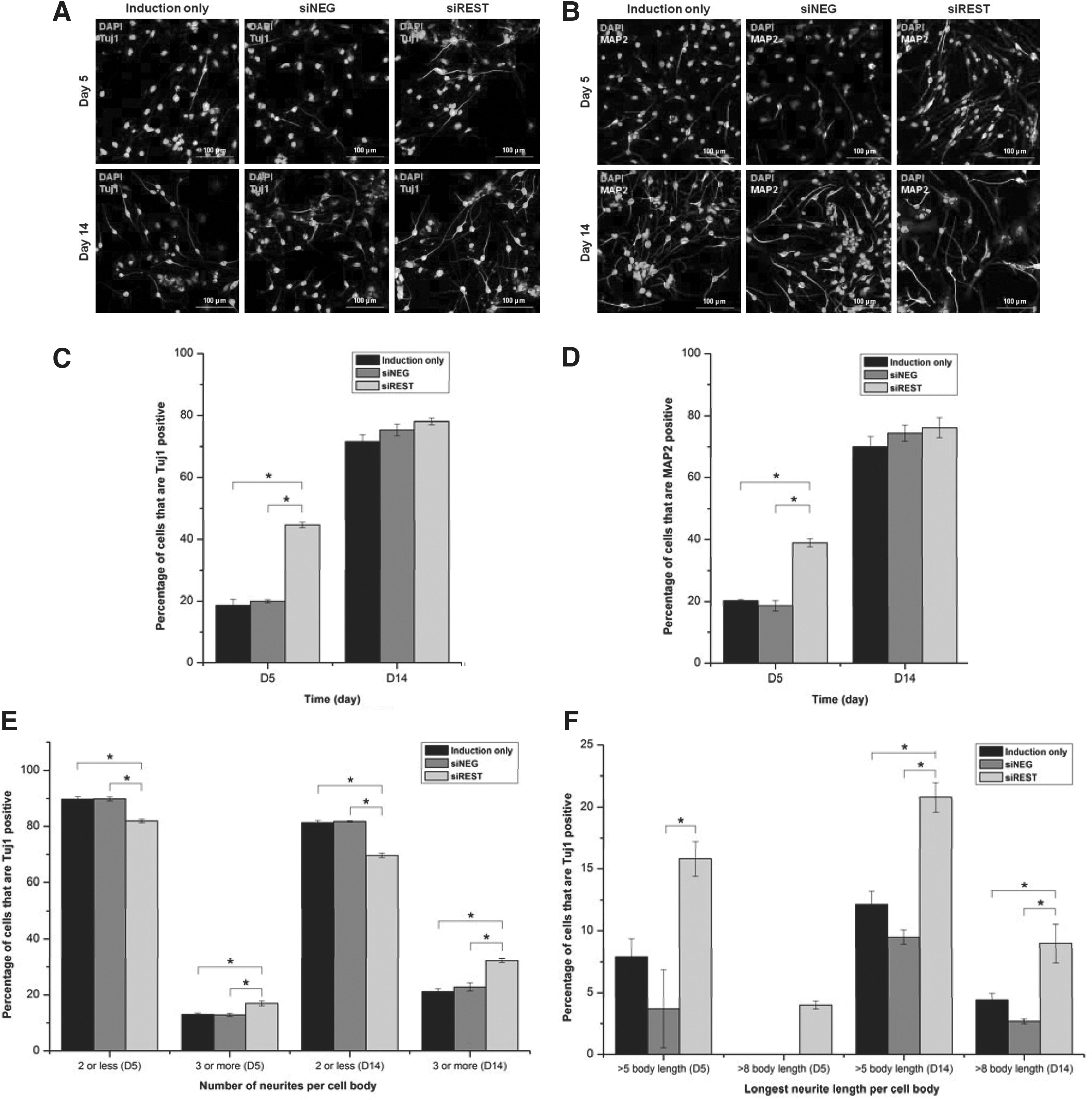
Immunostaining of
In terms of neuron maturation, Figure 8A shows that siREST day 14 cultures exhibited longer and a more complicated network of neurites. Quantitative analysis confirmed that REST-silenced neurons expressed a higher number of neurites than the control groups. At day 14, 30% of these neurons had three or more neurites per cell body while only 20% of differentiated neurons in the control groups fell into this category (Fig. 8E). REST-silencing also created a significant difference in neurite length between the experiemental groups. day 14 neurons could possess neurites of more than eight times of their body length. With REST knockdown, long neurites could be observed as early as day 5, and also at a higher frequency at day 14 (Fig. 8F). Collectively, the results showed that REST knockdown accelerated neuronal marker expression and neuron maturation in combination with neuronal induction.
Discussion
The conventional approach of directing stem cell differentiation often involves the overexpression of lineage-specific inductive factors. However, lineage commitment in stem cells entails complex parallel activation and repression of both positive and negative regulators. Therefore, the incorporation of gene-silencing to existing biochemical based differentiation protocol may provide an alternative synergistic approach to enhance the efficiency of directing stem cell fate. This method can potentially serve as a biomedical strategy for guided stem cell differentiation as demonstrated in this study. To date, only a few research groups have attempted to apply gene-silencing as a tool to promote neuronal differentiation from progenitors (Kim et al., 2010; Wurdak et al., 2010). The potency of this approach in facilitating differentiation has also been poorly evaluated. This study aims to provide a detailed kinetic analysis on induced differentiation and to establish a platform for quantifying the extent of accelerated differentiation.
REST is a negative regulator of many neuronal genes. Modulation of its expression during stem cell differentiation has been shown to facilitate commitment toward the neuronal lineage (Ballas et al., 2005; Yang et al., 2008; Kumar Gupta et al., 2009). However, in NPCs, the efficacy of REST knockdown versus conventional differentiation conditions in aiding neuronal commitment has not been examined to date. Accordingly, a transient knockdown of REST through non-viral delivery of REST siRNAs was carried out to evaluate its silencing effect on the extent of NPC differentiation into neuronal cells under nonspecific mixed-lineage and neuronal induction conditions. While the silencing was sustained for up to 7 days under mixed-lineage differentiation conditions (Fig. 1), efficient knockdown of REST expression was maintained for less than 3 days in neuronal induction medium (Fig. 5). Comparisons between the silencing profiles of REST under different induction conditions have not been previously evaluated in NPCs. Therefore, although the differences may be attributed to the activation of distinct molecular pathways governing the regulation and functionality of REST, the exact mechanisms behind this observation remain to be elucidated. In line with previous studies, our result suggested that gradual suppression of REST, being a physiological outcome of neurogenesis, occurred in both control and experimental samples during neuronal differentiation (Sun et al., 2005; Gao et al., 2011). At this stage, given a low endogenous REST expression level, a further inhibition of REST by RNAi was not as noticeable as that in nonspecific induction conditions. Therefore, our experimental set up for neuronal induction studies aimed to create an early REST knockdown, before the initiation of REST suppression by neuronal inductive signaling. To reinforce its initial suppression in neuronal induction medium, a second dose of siRNAs was added 1 day after the initial transfection to ensure effective knockdown of REST in the subsequent differentiation studies. For mixed-lineage/nonspecific induction conditions, the second dose of siRNAs was added 7 days after initial transfection to prolong the silencing of REST.
Under nonspecific induction, REST silencing alone directed neuronal lineage commitment. Tuj1 mRNA level was induced immediately 1 day after REST siRNA transfection (Fig. 6A) and remained at a higher level than controls after 5 and 14 days of nonspecific differentiation (Figs. 2B and 3D). Although Tuj1 expression was stimulated at a faster rate and in greater numbers for NPCs under REST silencing, the effect was not extended to mature neuronal marker, MAP2, within the 14-day evaluation period. The presence of serum in mixed-lineage induction conditions may have limited the extent of commitment toward the neuronal lineage while driving astrocytic differentiation. These findings deviated from our expectation because a previous study with MSCs has reported that silencing REST could induce MAP2 protein expression after 14 days (Yang et al., 2008).
Under the serum-free neuronal induction condition, REST knockdown was able to augment and accelerate differentiation of NPCs into mature neurons. Quantitative temporal analysis on the transcript level of neuronal markers revealed an earlier and stronger neuronal induction in REST-silenced NPCs compared with controls. This was particularly evident during the first week of differentiation (Fig. 6). This signifies the importance of a detailed kinetic analysis for the holistic evaluation of accelerated neuronal differentiation, which is usually unsatisfactorily presented in similar research works. For example, knocking down STAT3 was found to speed up osteogenesis by 10–14 days (Levy et al., 2010), and repressing Sox 6 shortened the period of cardiomyogenesis by 15 days (Sluijter et al., 2010). However, current research works concerning manipulated neuronal differentiation have lacked related quantitative comments. In contrast, and to achieve the benchmark for crosstalk with other lineages, our kinetic analysis showed that REST knockdown accelerated neuronal differentiation by 5–7 days.
Further quantification of neurite lengths and neurite numbers per Tuj1 positive cells also revealed more extensive neurite outgrowths with REST knockdown. The observed robust development of neuronal processes coincide with a study which reported clear increase in neurite length of neuroblastoma cells as a result of deregulation of genes encoding for neuronal polarity and axonal elongation with REST silencing (Lepagnol-Bestel et al., 2007). Accordingly, the results suggest that the modulation of REST levels not only aids in controlled neuronal differentiation of stem cells but also activates complex molecular mechanisms involved in nerve cell functionality such as neurite outgrowth. This observation further demonstrates the feasibility of utilizing RNAi as a biomedical approach to enhance stem cell differentiation and lineage-specific phenotypic maturation.
Collectively, these results showed that when combined with specific inductive or lineage-supportive factors, REST silencing increased the kinetics of neuronal marker expression, and in particular, greatly enhanced the rate of maturation of committed neuronal cells. Although recent findings demonstrated the efficacy of REST knockdown alone in driving neuronal commitment of ESCs or stem cells of non-neural origin, such kinetics analysis on neuronal marker expression has not been evaluated and our results strongly advocated the added benefit of faster neuron generation with REST silencing as opposed to regular differentiation approaches. Therefore, in neural trauma or pathological conditions where time delay between injury and therapy must be short for optimal therapeutic treatment, such efficient derivation of mature neuronal cells will provide support toward facilitating neural tissue regeneration.
This study has established RNAi of REST as a potential tool in neuronal tissue engineering and sheds light on the rapid generation of mature neuron for transplantation. Future works on neuronal differentiation of NPCs can potentially involve the synthesis of REST siRNA incorporated electrospun scaffolds due to its relative successes in supporting stem cells proliferation and differentiation (Carlberg et al., 2009; Christopherson et al., 2009; Xie et al., 2009; He et al., 2010), while recently demonstrating added versatility as a nanofiber-based siRNA delivery system (Cao et al., 2010; Rujitanaroj et al., 2011). Such biofunctional nanofibrous scaffolds can potentially enhance the differentiation of stem cells through synergistic integration of biochemical and topographical cues with scaffold-mediated RNAi.
Footnotes
Acknowledgments
This work is supported by A*Star BMRC SSCC Grant (09/016). The authors express gratitude to Dr. S. H. Lim for her valuable scientific discussions and for vetting through the manuscript.
Author Disclosure Statement
No conflicts of interest exist.