
Abstract
The development of the pancreas is a complicated process that is regulated on several levels. Pancreas transcription factor 1, alpha subunit (Ptf1a), also known as p48, is a pancreas-specific basic helix-loop-helix transcription factor that is critical for both exocrine pancreas development and maintenance of acinar cell differentiation. Based on a differential screening assay, we identified Rbms3, a gene encoding a glycine-rich RNA-binding protein, to be specifically expressed in the neural tube and the pancreatic rudiment of e10.5 embryos. The presence of Rbms3 in the early developing pancreas suggests that specific post-transcriptional regulation mechanisms play an important role in controlling pancreas development. In this study, we show that Rbms3 binds to the 3′UTR of Ptf1a mRNA, but not the 3′UTR of Pdx1, which is another pancreatic transcription factor. The ectopic expression of Rbms3 stimulates the translation of a reporter gene carrying the Ptf1a 3′UTR. In addition, when Rbms3 expression is suppressed in the AR42J-B13 pancreatic exocrine cell line, the expression of Ptf1a is also down-regulated. These results suggest that binding of Rbms3 to the 3′UTR of Ptf1a regulates the production of the Ptf1a protein and, thereby, indirectly regulates the expression of the Ptf1a downstream target genes.
Introduction
In addition to the transcriptional regulation of gene expression, the regulation of gene expression at the post-transcriptional level is emerging as a critical factor for many cellular and developmental processes (de Moor et al., 2005; MacNicol and MacNicol, 2010). In many cases, specific RNA-binding proteins interact dynamically with either the coding or untranslated regions of mRNAs to regulate the maturation, stability, and translation of RNAs (Lukong et al., 2008). Rbms3 was first identified because of its ability to bind to a DNA fragment derived from the promoter region of the mouse collagen a2(I) gene (Penkov et al., 2000). However, it did not exhibit any transcriptional activity. Because Rbms3 contains two RNA-binding motifs and localizes to the cytoplasm, it was speculated that Rbms3 is an RNA-binding protein. Later, it was discovered that Rbms3 expression is induced in activated hepatic stellate cells (HSCs) and fibrotic liver tissue. Rbms3 up-regulates the expression of Prx1, a transcription factor that promotes the fibrogenic transformation of HSCs, by binding to the 3′UTR of Prx1 mRNA (Fritz and Stefanovic, 2007).
In this study, we report a mechanism by which Rbms3 contributes to the post-transcriptional regulation of Ptf1a expression. We demonstrate that Rbms3 is expressed in the developing pancreas in a restricted pattern. Its expression corresponds to an increase in the Ptf1 expression level. In addition, the Rbms3 protein binds directly to the 3′UTR of Ptf1a mRNA, and this binding stabilizes Ptf1a mRNA and increases the production of Ptf1a protein. These results reveal a novel mechanism that regulates the expression of Ptf1a in a quantitative and dynamic fashion.
Materials and Methods
In situ hybridization and immunohistochemistry
Tissue preparation, in situ hybridization, and immunohistochemistry were performed as previously described (Wilkinson and Nieto, 1993), except for the antigen retrieval procedure that was added to the immunohistochemistry experiments. For staining Rbms3, sections were heated at 100°C in 10 mM Citrate buffer (pH 6.0) for 30 min before adding blocking solution. For staining Ptf1a and Pdx1, sections were heated in 10 mM Tris (pH 9.0) for 30 min. Rabbit anti-Rbms3 antibody was raised against aa 288–413 of the Rbms3 protein; rabbit anti-Ptf1a antibody was raised against aa 2–153 of the Ptf1a protein; and mouse anti-Pdx1 antibody was purchased from Abcam (Cat. No. ab47383).
Plasmid construct and lentivirus preparation
Full-length mouse Rbms3 cDNA was isolated by reverse transcription (RT)-polymerase chain reaction (PCR) using total RNA from e10.5 pancreatic rudiments and amplified with a 5′primer containing a BamHI site and a 3′ primer containing a KpnI site. The 1.6-kb Rbms3 cDNA was cloned into the BamHI and KpnI sites of the pPGK vector (Academia Sinica). To construct a GST-Rbms3 fusion protein, the full-length Rbms3 cDNA was cloned into the BamHI-XhoI sites of the pGEX4T-1 vector, and the resulting plasmid was named pGEX-Rbms3. The RNA-recognition motifs (RRMs) and the C-terminal lacking the RRMs of Rbms3 were also separately amplified by RT-PCR using proper primers and cloned into the BamHI-XhoI sites of the pGEX4T-1 vector. To generate the reporter constructs, the 3′UTRs of Ptf1a and Pdx1 were amplified from e11.5 mouse pancreas cDNA and cloned into the XhoI-BamHI sites of the pEGFP-C3 vector (Clontech), which placed them immediately behind the EGFP coding sequence. The plasmids used for producing the shRbms3 lentivirus (pLKO.1-shRbms3) and the lentivirus carrying scrambled shRNA (pLKO.1-shNullT) were obtained from the National RNAi Core Facility located at the Institute of Molecular Biology/Genomic Research Center, Academia Sinica in Taiwan. pLKO.1-shRbms3 contained a 21-nucleotide sequence corresponding to the coding sequence 582–602 (5′-GCAAGACCCTACGAATCTATA-3′) of the rat Rbms3 gene inserted into the pLKO.1 vector. All of the lentiviruses were prepared in HEK293T cells according to established protocols (Chen et al., 2010).
RNA UV-crosslinking assays
Uniformly labeled RNA probes were prepared by standard in vitro transcription protocols using [α-32P] UTP (3000 Ci/mmol; PerkinElmer) after linearizing the plasmids with the appropriate restriction enzymes. RNA–protein interaction assays were performed with glutathione S-transferase (GST) and GST fusion proteins purified from E. coli. Briefly, 200 ng of purified protein was mixed with [α-32P] RNA transcripts (10,000 cpm) in 20 μL of UV-crosslinking buffer (25 mM Tris-HCl, pH 7.8, 40 mM NaCl, 0.3 mM MgCl2, 10 μg/μL yeast tRNA, and 5% glycerol) and incubated at room temperature for 5 min. Reaction mixtures were then exposed to 254 nm UV irradiation at a distance of 3 cm for 1 min in a Select™ Series UV Crosslinkers (Spectroline). After UV crosslinking, reactions were treated with RNaseT1 at 37°C for 20 min. Samples were separated on a 10% polyacrylamide SDS gel, which was then stained with Coomassie Brilliant Blue R-250, dried, and exposed to film with an intensifying screen at −80°C overnight. For competition experiments, the protein and radioactive RNA were incubated for 5 min, and then various amounts of cold Ptf1a 3′UTR were added to the reactions and allowed to equilibrate for 20 min.
Quantitative RT-PCR analysis
Total cellular RNA extractions were prepared by using the Trizol reagent (Genemark). cDNA was prepared by using the first-strand cDNA synthesis kit (Epicentre) according to the manufacturer's instructions. Real-time PCR was performed using the StepOne™ Real-Time PCR system (Applied Biosystem). The primer sets used in this study include the following: Rbms3-F, 5′-TGACAAGAATACGAATCAGTGCAAA-3′; Rbms3-R, 5′-TCGTAGGGTCTTGCTCTTGTTG-3′; Ptf1a-F, 5′-TTTTGACAACATAGAGAACGAACCA-3′; Ptf1a-R, 5′-CACGTACAATATGCACAAAGACACA-3′; Cpa1-F, 5′-GTGTGGACCCCAACAGGAACT-3′; Cpa1-R, 5′-TGGGAAATTTGCCTCGGTAA-3′; Amylase-F, 5′-GAATGTAGTCAACGGTCAGCCTTT-3′; Amylase-R, 5′-GAATCCTCTGTTTCCTCTGCTAAAA-3′; Ctrb1-F, 5′-GGCTCCGACGAAGAGAACATC-3′; Ctrb1-R, 5′-GATGTCATTGCGCACAGTGAA-3′; Actin-F, 5′-ACCGTGAAAAGATGACCCAGAT-3′; Actin-R, 5′-AGAGGCATACAGGGACAACACA-3′; Gapdh-F, 5′-GGTGGACCTCATGGCCTACA-3′; Gapdh-R, 5′-AGCAACTGAGGGCCTCTCTCT-3′. The expression levels of the genes analyzed were normalized to the housekeeping genes Actin and Gapdh.
Reporter gene analysis
For the analysis, 0.1 μg of pEGFP-3′Ptf1a (or pEGFP-3′Pdx1) plasmid was co-transfected with various amounts (0.1, 0.4, or 1.6 μg) of pPGK-Rbms3 (or control pPGK) plasmid into HEK293T cells. As an internal control, 0.05 μg of a plasmid containing the β-galactosidase gene driven by an SV40 promoter was included in all transfection experiments. Transfections were performed using the TransIT-LT1 transfection reagent (Mirus). Cells were harvested 2 days after transfection, and total RNA or proteins were extracted. The transfection efficiency was determined by a β-galactosidase assay.
mRNA stability determination
To determine the mRNA half-life, AR42J-B13 cells were first infected by shRbms3 lentivirus or shNullT lentivirus. Two days after the infections, Actinomycin-D (5 μg/mL) was added to the cells, and total RNA was prepared at the times indicated. The mRNA half-lives were calculated after quantifying by quantitative RT-PCR.
Results
Rbms3 is expressed in a restricted pattern in the early developing pancreas
To identify genes that are specifically expressed in the early developing pancreas, single-cell PCR products made from various individual e10.5 pancreatic cells were used to differentially screen microarrays that contained 9734 cDNA clones (Chiang and Melton, 2003). One of the clones, designated Pds1 (pancreatic differential screen 1), was found to match the sequence of Rbms3. The specific expression of Rbms3 in the early developing pancreas was confirmed by in situ hybridization. As shown in Figure 1A, Rbms3 was first detected in the e10.5 pancreatic rudiment, neural tube, and dorsal root ganglion. In addition, within the developing pancreas at this stage, Rbms3 appears to be expressed in the undifferentiated pancreatic progenitor cells because the expression pattern of Rbms3 does not overlap with the expression of protein tyrosine phosphatase-NP (PTP-NP), a marker expressed in cells of the endocrine lineage (Chiang and Flanagan, 1996). Rbms3 protein was also detected in the developing pancreas by immunohistochemical staining (Fig. 1D, G, and H). The staining of the Rbms3-expressing cells by the anti-Rbms3 antibody is consistent with the in situ hybridization result, and remains similar on e12.5 and e13.5 embryo sections (Fig. 1G, H). Within the developing pancreas, the broad staining pattern of Rbms3 is similar to that of a transcription factor, Ptf1a (Fig. 1E), and different from the punctate staining pattern of another transcription factor, Ngn3 (Fig. 1F), demonstrating the specificity of the anit-Rbms3 antibody. Although weak expression of Rbms3 was detected in other organs after e12.5, the developing pancreas and neural tube still exhibited the strongest expression of Rbms3. As shown by quantitative PCR, the expression level of Rbms3 peaked in the pancreatic rudiment at e13.5, and this expression level was at least 10-fold higher than the level detected in any tested adult organs (Fig. 2).
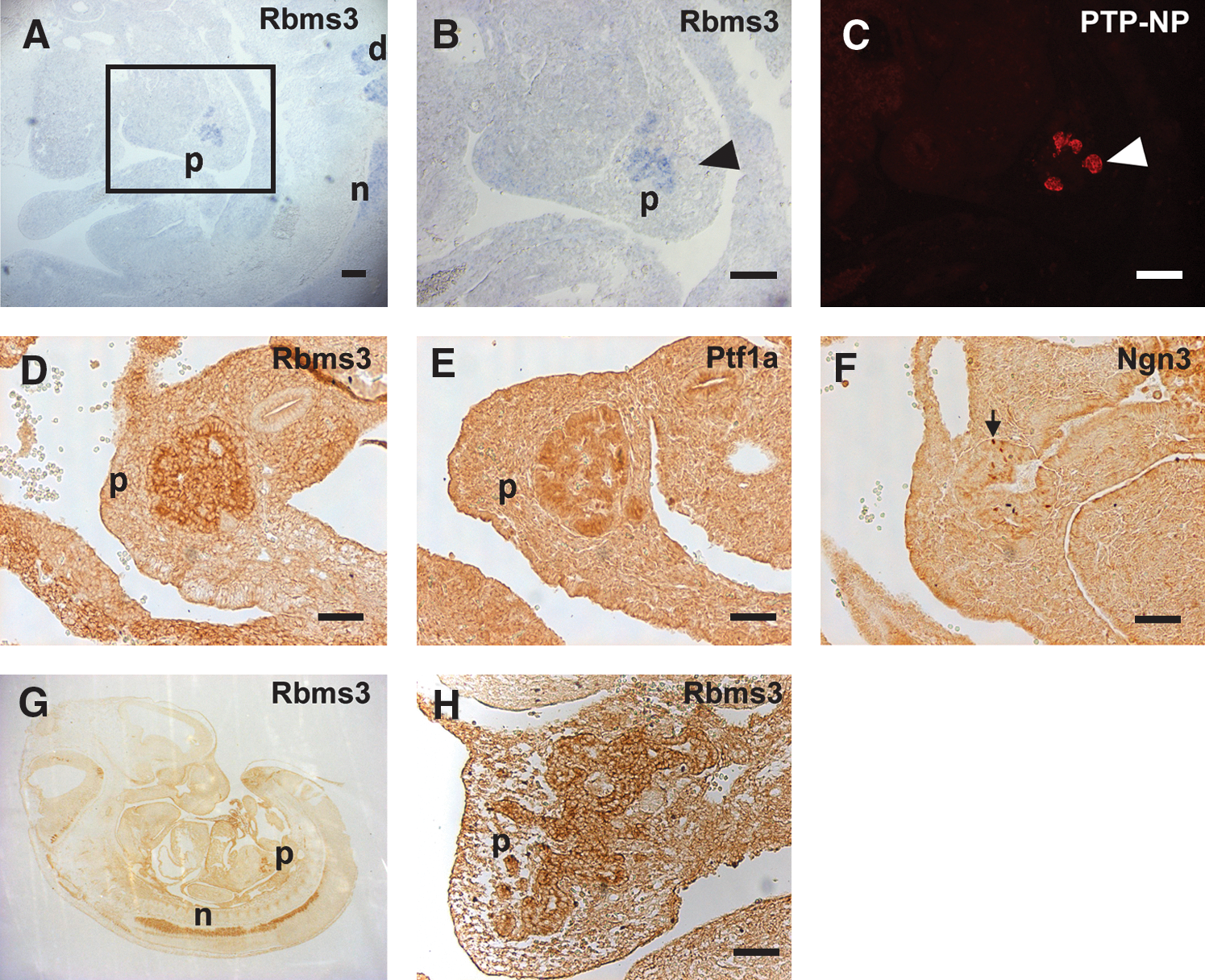
Rbms3 is expressed in the developing pancreas and neural tube in a restricted pattern.
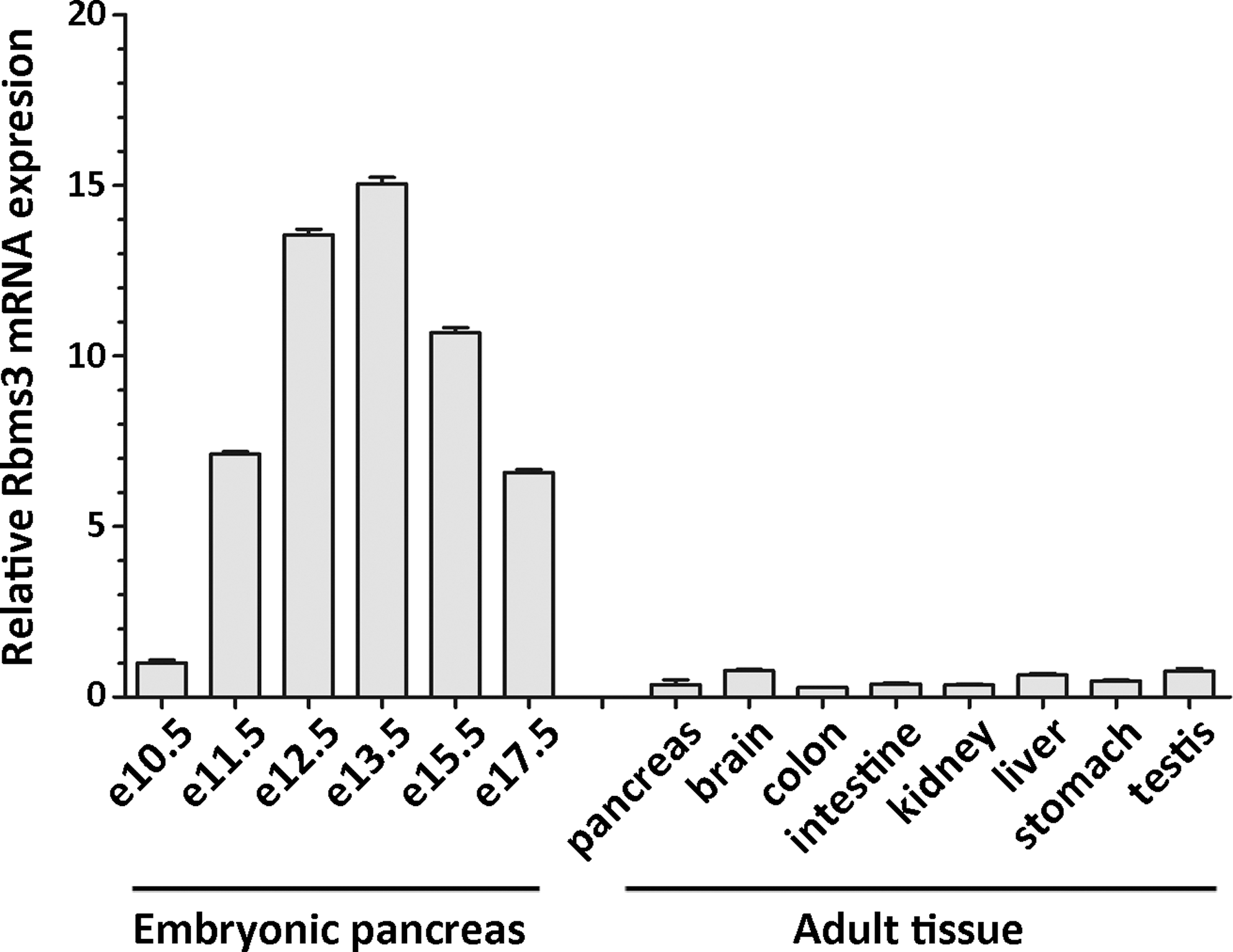
Rbms3 is expressed stronger in developing pancreas than in adult tissues. mRNA of Rbms3 was quantified by quantitative RT-PCR with RNA isolated from embryonic pancreatic rudiments and various adult tissues. The expression levels were determined relative to the level in the e10.5 pancreatic rudiment. PCR, polymerase chain reaction.
Rbms3 specifically binds to the 3′UTR of Ptf1a mRNA
As expected, Rbms3 was expressed in the progenitor cells of the early developing pancreas. Its expression pattern was similar to the expression patterns of two other documented transcription factors, Pdx1 and Ptf1a (Fig. 1E). Rbms3 is an RNA-binding protein that has been shown to regulate protein expression by binding to the 3′UTR of its target genes. Therefore, a UV-crosslinking assay was performed to test the interaction between Rbms3 protein and the 3′UTRs of Pdx1 and Ptf1a mRNA. As shown in Figure 3A, purified GST and various GST-Rbms3 fusion proteins were used in this analysis. Notably, a strong signal corresponding to the migration position of the RNA/GST-Rbms3 complex, as confirmed by Coomassie-blue staining (data not shown), was only observed when GST-Rbms3 was incubated with the labeled Pft1a 3′UTR and not the Pdx1 3′UTR or the Gapdh3′UTR. Even though two RRMs are located in the N-terminal half of Rbms3, neither of the fusion proteins containing only the N-terminal half or the C-terminal half of Rbms3 appeared to bind the 3′UTR of Ptf1a. To further demonstrate the binding specificity between Rbms3 and the 3′UTR of Ptf1a, an excess of cold Ptf1a 3′UTR was added to the reaction after the initial 5-min incubation with GST-Rbms3 and labeled Ptf1a 3′UTR. Figure 3B shows that the labeled Ptf1a 3′UTR that was initially bound to the GST-Rbms3 fusion protein is competed off as the amount of excess Ptf1a 3′UTR increases.
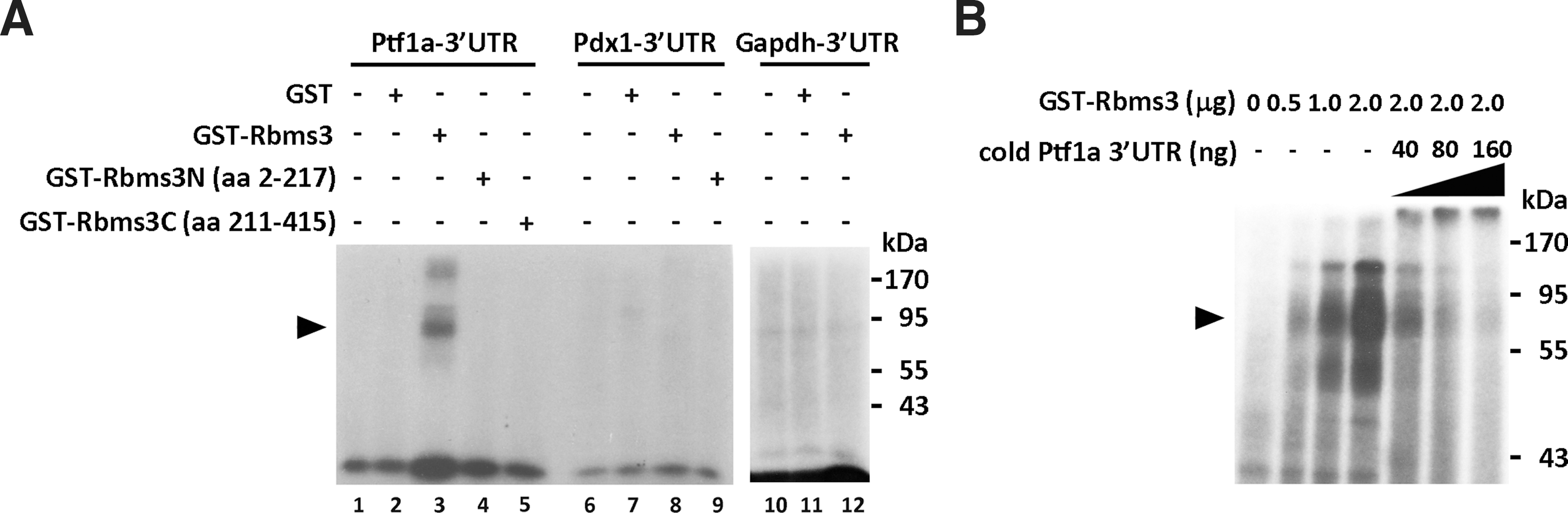
Rbms3 specifically binds to the 3′UTR of Ptf1a.
Rbms3 increases the expression of the GFP reporter gene containing the Ptf1a 3 ′UTR
To study the biological relevance of the binding between Rbms3 and the Ptf1a 3′UTR, we constructed two reporter genes. Both of these reporter genes are driven by the CMV promoter and contain the GFP coding sequence fused to either the 3′UTR of mouse Ptf1a or Pdx1 mRNA (Fig. 4A). The reporter genes were co-transfected into HEK293T cells with either the Rbms3 expression plasmid or a control expression plasmid. The pCH110 vector (0.05 μg) carrying the β-galactosidase gene was used as an internal control in each transfection to calibrate the transfection efficiency. Forty-eight hours after the transfection, cell lysates that were adjusted to contain equal amounts of β-galactosidase activity were loaded on SDS-PAGE gel and the expression of the GFP protein was measured by western blot analysis. As shown in Figure 4B, the expression level of the reporter gene containing the Ptf1a 3′UTR increased with the ectopic expression of Rbms3. Conversely, the expression level of the reporter gene containing the Pdx1 3′UTR showed no change with the presence of ectopic Rbms3. Figure 4C also shows that the expression level of the reporter gene containing the Ptf1a 3′UTR increases proportionally to increasing amounts of ectopic Rbms3. The highest level of expression reached an approximately fivefold increase from the basal expression level.
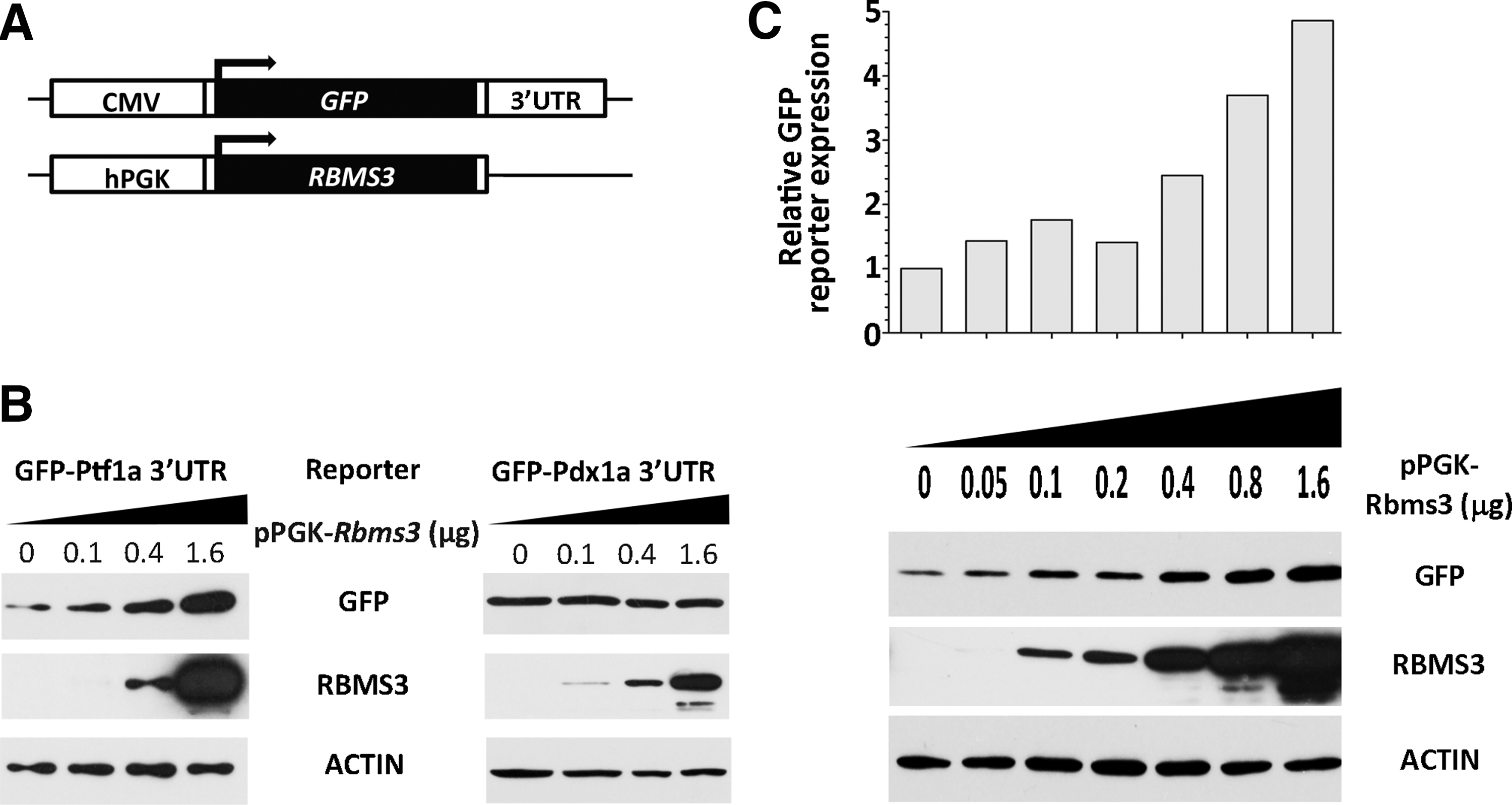
Rbms3 increases the expression of a GFP reporter gene containing the 3′UTR of Ptf1a.
Rbms3 is required in the pancreatic exocrine cell line AR42J-B13 to maintain a stable expression of Ptf1a and its downstream target genes
To verify that Rbms3 regulates the expression of endogenous Ptf1a, we down-regulated the expression of Rbms3 in the pancreatic exocrine cell line AR42J-B13 using an RNA interference technique. Lentiviruses carrying nontargeting shRNA (shNullT) or shRNA targeting Rbms3 (shRbms3) were generated and used to infect AR42J-B13 cells. As shown in Figure 5A, the expression level of Rbms3 was significantly down-regulated in the exocrine cell line infected with shRbms3. This decrease in expression was verified by quantitative PCR. Evidently, the RNA level of Ptf1a also decreased, as well as the RNA level of known Ptf1a downstream targets, such as Carboxypeptidase A1 (CPA1), Amylase, and ChymotrypsinogenB1. Consequently, the protein levels of Ptf1 and its downstream targets were also affected by the down-regulation of Rbms3 (Fig. 5B). Because Rbms3 binds directly to the Ptf1a 3′UTR and affects the expression level of Ptf1a mRNA, we next investigated whether Rbms3 regulates Ptf1a mRNA stability. For this experiment, the transcriptional inhibitor actinomycin D (5 μg/mL) was added to the medium after the AR42J-B13 cells were treated with either shNullT or shRbms3. The cells were then harvested at the indicated times. The level of Ptf1a mRNA was determined by quantitative RT-PCR and compared to the initial level at time zero. Figure 5C shows that the level of Ptf1a mRNA decreased more rapidly when the expression of Rbms3 was down-regulated. This result suggests that Rbms3 is required to stabilize Ptf1a mRNA. When Rbms3 expression is down-regulated, the half-life of Ptf1a mRNA decreases, which results in a decrease in Ptf1a protein production.
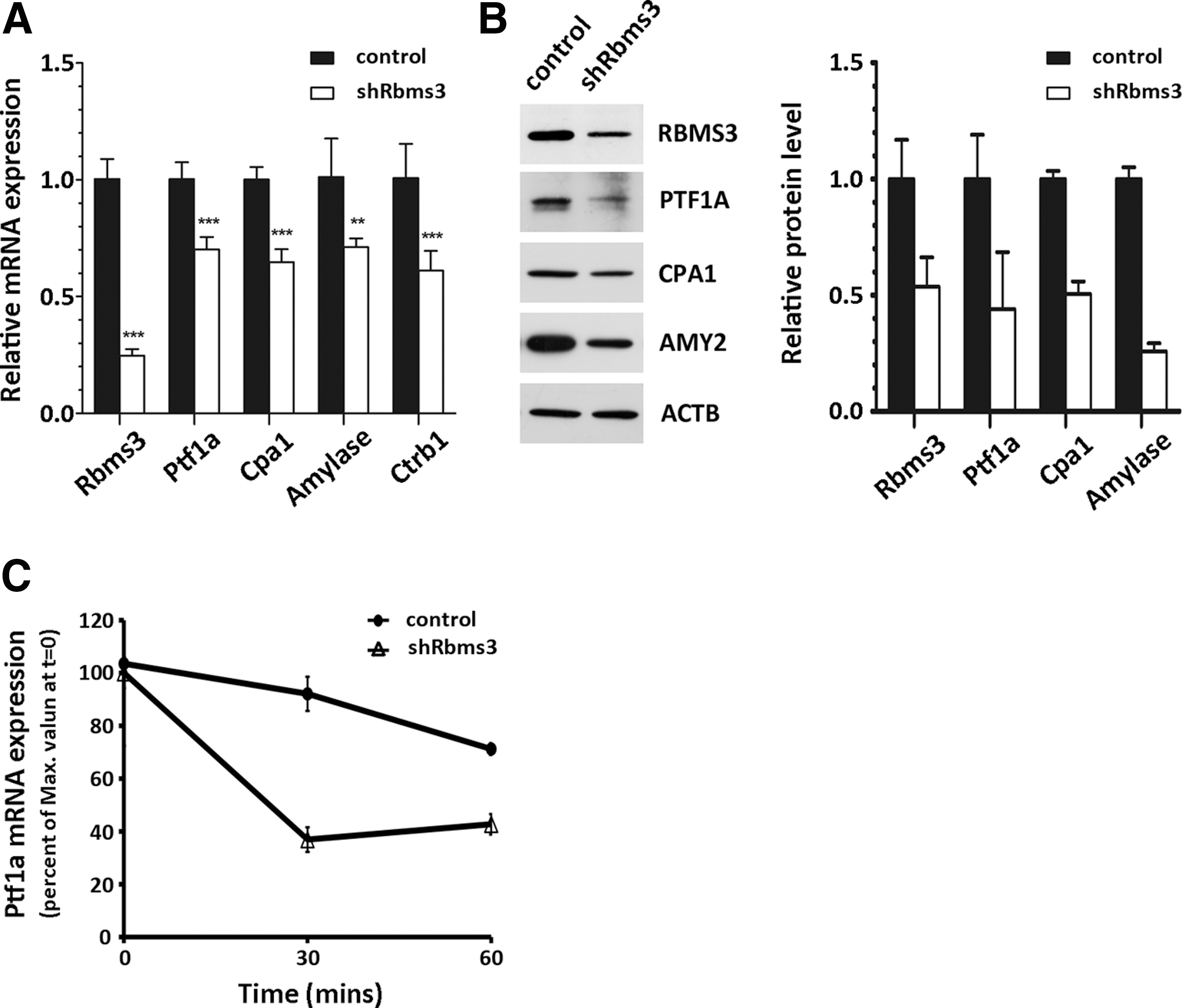
Decreased Rbms3 expression affects the expression of Ptf1a and its downstream target genes in AR42J-B13 exocrine cells.
Discussion
Ptf1a plays critical roles during two different stages of pancreas development. In mice, the expression of Ptf1a is required at e10.5 to specify the pancreatic cell fate. At this stage, not all the cells expressing Ptf1a will become exocrine cells. Later during development, however, the sudden rise in the Ptf1a expression level in a majority of the pancreatic cells does result in the cells committing to an exocrine cell fate. In this study, we show that, in addition to the transcriptional regulation responsible for the spatial and temporal expression pattern of Ptf1a, the post-transcriptional regulation of Ptf1a also plays an important role in its rapid increase in expression during the second transition of pancreas development.
Rbms3 was first identified in an expression screening for proteins that can bind to the promoter sequence of the collagen a2(I) gene. It belongs to a family of MSSPs, whose members may be involved in DNA replication, transcription, apoptosis, and cell-cycle progression (Iida et al., 1997; Niki et al., 2000). However, Rbms3 does not appear to affect gene transcription. Because of its two RNA-binding motifs and its localization to the cytoplasm, Rbms3 may affect its target genes at the post-transcriptional level. In this study, we show that Rbms3 is expressed restrictedly in the embryonic pancreas, neural tube, and dorsal root ganglion. The expression of Rbms3 in the early pancreatic progenitor cells and in the differentiating exocrine cells is vastly overlapping, if not identical, with the expression pattern of Ptf1a. However, the expression level of Rbms3 decreases after e13.5, whereas Ptf1a remains strongly expressed in the exocrine cells throughout the development (Supplementary Fig. S1; Supplementary Data are available online at
During early mouse development from e10.5 to e12.5, pancreatic progenitor cells express a number of transcription factors, such as Pdx1, Ptf1a, Nkx2.2, and Nkx6.1, that have overlapped and distinct functions (Chiang and Melton, 2003). At this stage, these Ptf1a-expressing cells have not determined their fates. The expression of Ptf1a, however, becomes restricted to acinar progenitor cells in the mouse pancreas by e13.5, and this Ptf1a expression is much stronger than the expression in the pancreatic progenitor cells. Consequently, this strong expression of Ptf1a initiates the exocrine cell differentiation program. Although it has been shown that a 2.3 kb autoregulatory enhancer region initiates the expression of Ptf1a in the acinar precursor epithelium and then super-induces its expression in nascent acinar cells, the dramatic increase in Ptf1a expression in acinar precursor cells may involve an additional regulatory mechanism. It is of interest that Rbms3 is expressed specifically in the embryonic pancreas, and its expression reaches a peak level at e13.5, which coincides with the timing of exocrine differentiation. Its presence and ability to stabilize Ptf1a mRNA may contribute to the sudden rise in the expression level of Ptf1a and thus help to specify the fate of exocrine cells. After the cells determine their exocrine fate, additional mechanisms may manage to maintain or enhance the expression of Ptf1a in the terminally differentiated exocrine cells. In our experiments, however, the down-regulation of Rbms3 did not change the cell fate of AR42J-B13 cells (data not shown) because the markers of endocrine and ductal cells could not be detected in the treated cells. This result may be because the effect of Rbms3 on cell fate determination is only critical at specific developmental stages, such as e13.5. Therefore, the fate of terminally differentiated exocrine cells could not be reversed by simply down-regulating Ptf1a expression. Nevertheless, the down-regulation of Rbms3 expression does affect the expression of various digestive enzymes, which suggests that Rbms3 also plays an important role in maintaining the physiological function of mature pancreatic exocrine cells.
Footnotes
Acknowledgment
This work was supported by the National Science Council, ROC (NSC98-2311-B-194-001-MY3).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.